Zellwandspannung bei Pflanzen. Zellenwand
Zu Beginn der Entwicklung des Lebens auf der Erde wurden alle Zellformen durch Bakterien repräsentiert. Sie nahmen organische Substanzen, die im Urmeer gelöst waren, über die Körperoberfläche auf.
Im Laufe der Zeit haben sich einige Bakterien daran angepasst, aus anorganischen Substanzen organische Substanzen herzustellen. Dazu nutzten sie die Energie des Sonnenlichts. Der erste entstand ökologisches System, in dem diese Organismen Produzenten waren. Dadurch gelangte der von diesen Organismen freigesetzte Sauerstoff in die Erdatmosphäre. Mit seiner Hilfe können Sie aus derselben Nahrung viel mehr Energie gewinnen und die zusätzliche Energie nutzen, um die Struktur des Körpers zu komplizieren: den Körper in Teile zu teilen.
Einer von wichtige Erfolge Leben - Trennung von Kern und Zytoplasma. Der Zellkern enthält Erbinformationen. Eine spezielle Membran um den Kern ermöglichte den Schutz vor unbeabsichtigter Beschädigung. Bei Bedarf erhält das Zytoplasma vom Zellkern Befehle, die das Leben und die Entwicklung der Zelle steuern.
Organismen, bei denen der Zellkern vom Zytoplasma getrennt ist, haben das nukleare Superreich gebildet (dazu gehören Pflanzen, Pilze und Tiere).
So entstand und entwickelte sich die Zelle – die Grundlage der Organisation von Pflanzen und Tieren – im Laufe der biologischen Evolution.
Schon mit bloßem Auge, oder noch besser unter einer Lupe, kann man erkennen, dass das Fruchtfleisch einer reifen Wassermelone aus sehr kleinen Körnern bzw. Körnern besteht. Dies sind Zellen – die kleinsten „Bausteine“, aus denen der Körper aller lebenden Organismen, einschließlich Pflanzen, besteht.
Das Leben einer Pflanze erfolgt durch die gemeinsame Aktivität ihrer Zellen, wodurch ein einziges Ganzes entsteht. Bei der Mehrzelligkeit von Pflanzenteilen kommt es zu einer physiologischen Differenzierung ihrer Funktionen, einer Spezialisierung verschiedener Zellen je nach ihrer Lage im Pflanzenkörper.
Eine Pflanzenzelle unterscheidet sich von einer tierischen Zelle dadurch, dass sie eine dichte Membran hat, die den inneren Inhalt von allen Seiten bedeckt. Die Zelle ist nicht flach (wie es normalerweise dargestellt wird), sondern sieht höchstwahrscheinlich wie eine sehr kleine Blase aus, die mit schleimigem Inhalt gefüllt ist.
Struktur und Funktionen einer Pflanzenzelle
Betrachten wir eine Zelle als strukturelle und funktionelle Einheit eines Organismus. Die Außenseite der Zelle ist mit einer dichten Zellwand bedeckt, in der sich dünnere Abschnitte, sogenannte Poren, befinden. Darunter befindet sich ein sehr dünner Film – eine Membran, die den Zellinhalt – das Zytoplasma – bedeckt. Im Zytoplasma gibt es Hohlräume – mit Zellsaft gefüllte Vakuolen. In oder nahe der Mitte der Zelle Zellenwand es gibt einen dichten Körper – einen Kern mit einem Nukleolus. Der Kern ist durch die Kernhülle vom Zytoplasma getrennt. Kleine Körper, sogenannte Plastiden, sind im Zytoplasma verteilt.
Struktur Pflanzenzelle
Struktur und Funktionen pflanzlicher Zellorganellen
| Organoid | Zeichnung | Beschreibung | Funktion | Besonderheiten |
Zellwand oder Plasmamembran | Farblos, transparent und sehr langlebig | Leitet Substanzen in die Zelle hinein und aus ihr heraus. | Die Zellmembran ist semipermeabel |
|
Zytoplasma | Dickflüssige Substanz | Alle anderen Teile der Zelle befinden sich darin | Ist in ständiger Bewegung |
|
Zellkern (wichtiger Teil der Zelle) | Rund oder oval | Sorgt für die Übertragung erblicher Eigenschaften auf Tochterzellen während der Teilung | Zentraler Teil der Zelle |
|
Kugelförmige oder unregelmäßige Form | Beteiligt sich an der Proteinsynthese | |||
 | Ein durch eine Membran vom Zytoplasma getrenntes Reservoir. Enthält Zellsaft | Überschüssige Nährstoffe und Abfallprodukte, die die Zelle nicht benötigt, sammeln sich an. | Während die Zelle wächst, verschmelzen kleine Vakuolen zu einer großen (zentralen) Vakuole |
|
Plastiden | Chloroplasten | Sie nutzen die Lichtenergie der Sonne und erzeugen aus Anorganischem Organisches | Die Form von Scheiben, die durch eine Doppelmembran vom Zytoplasma abgegrenzt sind |
|
Chromoplasten | Entsteht durch die Ansammlung von Carotinoiden | Gelb, Orange oder Braun |
||
 | Leukoplasten | Farblose Plastiden | ||
Atomhülle | Besteht aus zwei Membranen (äußere und innere) mit Poren | Trennt den Zellkern vom Zytoplasma | Ermöglicht den Austausch zwischen Zellkern und Zytoplasma |
Der lebende Teil einer Zelle ist ein membrangebundenes, geordnetes, strukturiertes System aus Biopolymeren und inneren Membranstrukturen, die an einer Reihe von Stoffwechsel- und Energieprozessen beteiligt sind, die das gesamte System als Ganzes aufrechterhalten und reproduzieren.
Ein wichtiges Merkmal ist, dass die Zelle keine offenen Membranen mit freien Enden hat. Zellmembranen begrenzen Hohlräume oder Bereiche immer und verschließen sie nach allen Seiten.

Modernes verallgemeinertes Diagramm einer Pflanzenzelle
Plasmalemma(äußere Zellmembran) ist ein ultramikroskopischer Film mit einer Dicke von 7,5 nm, der aus Proteinen, Phospholipiden und Wasser besteht. Dabei handelt es sich um einen sehr elastischen Film, der von Wasser gut benetzt wird und nach einer Beschädigung schnell seine Integrität wiederherstellt. Es hat eine universelle Struktur, d. h. typisch für alle biologischen Membranen. In Pflanzenzellen gibt es außerhalb der Zellmembran eine starke Zellwand, die für äußere Unterstützung sorgt und die Form der Zelle beibehält. Es besteht aus Ballaststoffen (Cellulose), einem wasserunlöslichen Polysaccharid.
Plasmodesmen Pflanzenzellen sind submikroskopisch kleine Röhrchen, die die Membranen durchdringen und mit einer Plasmamembran ausgekleidet sind, die so ohne Unterbrechung von einer Zelle zur anderen gelangt. Mit ihrer Hilfe kommt es zu einer interzellulären Zirkulation von Lösungen, die organische Nährstoffe enthalten. Sie übertragen auch Biopotentiale und andere Informationen.
Porami sogenannte Öffnungen in der Sekundärmembran, wo die Zellen nur durch die Primärmembran und die mittlere Lamina getrennt sind. Die Bereiche der Primärmembran und der Mittelplatte, die die benachbarten Poren benachbarter Zellen trennen, werden Porenmembran oder Verschlussfilm der Pore genannt. Der Verschlussfilm der Pore wird von plasmodesmalen Tubuli durchstoßen, in den Poren bildet sich jedoch meist kein Durchgangsloch. Poren erleichtern den Transport von Wasser und gelösten Stoffen von Zelle zu Zelle. Poren bilden sich in den Wänden benachbarter Zellen, meist einander gegenüber.
Zellmembran hat eine gut definierte, relativ dicke Schale aus Polysacchariden. Die Hülle einer Pflanzenzelle ist ein Produkt der Aktivität des Zytoplasmas. An seiner Entstehung sind der Golgi-Apparat und das endoplasmatische Retikulum aktiv beteiligt.

Struktur der Zellmembran
Die Basis des Zytoplasmas ist seine Matrix oder Hyaloplasma, ein komplexes farbloses, optisch transparentes kolloidales System, das zu reversiblen Übergängen vom Sol zum Gel fähig ist. Die wichtigste Aufgabe des Hyaloplasmas besteht darin, alle Zellstrukturen zu vereinen einheitliches System und Sicherstellung der Interaktion zwischen ihnen in den Prozessen des Zellstoffwechsels.
Hyaloplasma(oder zytoplasmatische Matrix) ist interne Umgebung Zellen. Besteht aus Wasser und verschiedenen Biopolymeren (Proteine, Nukleinsäuren, Polysaccharide, Lipide), von denen der Großteil aus Proteinen unterschiedlicher chemischer und funktioneller Spezifität besteht. Das Hyaloplasma enthält außerdem Aminosäuren, Monosaccharide, Nukleotide und andere niedermolekulare Substanzen.
Biopolymere bilden mit Wasser ein kolloidales Medium, das je nach Bedingungen sowohl im gesamten Zytoplasma als auch in seinen einzelnen Abschnitten dicht (in Form eines Gels) oder flüssiger (in Form eines Sols) sein kann. Im Hyaloplasma sind verschiedene Organellen und Einschlüsse lokalisiert und interagieren miteinander und mit der Hyaloplasmaumgebung. Darüber hinaus ist ihr Standort meist spezifisch für bestimmte Zelltypen. Über die Bilipidmembran interagiert das Hyaloplasma mit der extrazellulären Umgebung. Daher ist Hyaloplasma ein dynamisches Medium und spielt wichtige Rolle in der Funktion einzelner Organellen und dem Leben der Zellen als Ganzes.
Zytoplasmatische Formationen - Organellen
Organellen (Organellen) sind strukturelle Bestandteile des Zytoplasmas. Sie haben eine bestimmte Form und Größe und sind obligatorische zytoplasmatische Strukturen der Zelle. Fehlen sie oder sind sie beschädigt, verliert die Zelle meist ihre Überlebensfähigkeit. Viele der Organellen sind zur Teilung und Selbstreproduktion fähig. Ihre Größe ist so klein, dass sie nur mit einem Elektronenmikroskop sichtbar sind.
Kern
Der Zellkern ist das prominenteste und meist größte Organell der Zelle. Es wurde erstmals 1831 von Robert Brown im Detail erforscht. Der Zellkern stellt die wichtigsten Stoffwechsel- und genetischen Funktionen der Zelle bereit. Die Form ist sehr unterschiedlich: Sie kann kugelförmig, oval, gelappt oder linsenförmig sein.
Der Zellkern spielt eine wichtige Rolle im Leben der Zelle. Eine Zelle, deren Kern entfernt wurde, scheidet keine Membran mehr aus und hört auf zu wachsen und Substanzen zu synthetisieren. Die Zerfalls- und Zerstörungsprodukte verstärken sich darin, wodurch es schnell stirbt. Die Bildung eines neuen Zellkerns aus dem Zytoplasma findet nicht statt. Neue Kerne entstehen nur durch Teilung oder Zerkleinerung des alten.
Der innere Inhalt des Kerns ist Karyolymphe (Kernsaft), der den Raum zwischen den Strukturen des Kerns ausfüllt. Es enthält einen oder mehrere Nukleolen sowie eine beträchtliche Anzahl von DNA-Molekülen, die mit bestimmten Proteinen – Histone – verbunden sind.

Kernstruktur
Nukleolus
Der Nukleolus enthält wie das Zytoplasma überwiegend RNA und spezifische Proteine. Seine wichtigste Funktion besteht darin, dass es Ribosomen bildet, die in der Zelle die Synthese von Proteinen durchführen.
Golgi-Apparat
Der Golgi-Apparat ist ein Organell, das in allen Arten eukaryontischer Zellen universell verbreitet ist. Es handelt sich um ein mehrschichtiges System flacher Membransäcke, die sich entlang der Peripherie verdicken und Blasenfortsätze bilden. Es befindet sich am häufigsten in der Nähe des Kerns.

Golgi-Apparat
Der Golgi-Apparat umfasst notwendigerweise ein System kleiner Vesikel (Vesikel), die von verdickten Zisternen (Scheiben) abgelöst werden und sich entlang der Peripherie dieser Struktur befinden. Diese Vesikel spielen die Rolle intrazellulärer Transportsystem spezifische Sektorkörnchen können als Quelle für zelluläre Lysosomen dienen.
Zu den Funktionen des Golgi-Apparats gehört auch die Ansammlung, Trennung und Freisetzung außerhalb der Zelle mit Hilfe von Vesikeln intrazellulärer Syntheseprodukte, Zerfallsprodukte und toxischer Substanzen. Produkte der synthetischen Aktivität der Zelle sowie verschiedene Substanzen, die aus der Umgebung über die Kanäle des endoplasmatischen Retikulums in die Zelle gelangen, werden zum Golgi-Apparat transportiert, sammeln sich in diesem Organell an und gelangen dann in Form von Tröpfchen oder Körnern in das Zytoplasma und werden entweder von der Zelle selbst genutzt oder nach außen ausgeschieden. . In Pflanzenzellen enthält der Golgi-Apparat Enzyme für die Synthese von Polysacchariden und das Polysaccharidmaterial selbst, das zum Aufbau der Zellwand verwendet wird. Es wird angenommen, dass es an der Bildung von Vakuolen beteiligt ist. Der Golgi-Apparat wurde nach dem italienischen Wissenschaftler Camillo Golgi benannt, der ihn 1897 erstmals entdeckte.
Lysosomen
Lysosomen sind kleine, von einer Membran begrenzte Vesikel, deren Hauptfunktion darin besteht, die intrazelluläre Verdauung durchzuführen. Die Verwendung des lysosomalen Apparats erfolgt während der Keimung eines Pflanzensamens (Hydrolyse der Reserve). Nährstoffe).
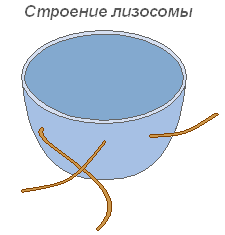
Struktur eines Lysosoms
Mikrotubuli
Mikrotubuli sind membranöse, supramolekulare Strukturen, die aus spiralförmig oder geraden Reihen angeordneten Proteinkügelchen bestehen. Mikrotubuli erfüllen überwiegend eine mechanische (motorische) Funktion und sorgen für die Beweglichkeit und Kontraktilität der Zellorganellen. Sie befinden sich im Zytoplasma, geben der Zelle eine bestimmte Form und sorgen für die Stabilität der räumlichen Anordnung der Organellen. Mikrotubuli fördern die Bewegung von Organellen zu bestimmten Orten physiologische Bedürfnisse Zellen. Eine beträchtliche Anzahl dieser Strukturen befindet sich im Plasmalemma, in der Nähe der Zellmembran, wo sie an der Bildung und Ausrichtung von Cellulose-Mikrofibrillen pflanzlicher Zellwände beteiligt sind.

Mikrotubuli-Struktur
Vakuole
Die Vakuole ist der wichtigste Bestandteil pflanzlicher Zellen. Es handelt sich um eine Art Hohlraum (Reservoir) in der Masse des Zytoplasmas, der mit einer wässrigen Lösung gefüllt ist Mineralsalze, Aminosäuren, organische Säuren, Pigmente, Kohlenhydrate und durch eine Vakuolenmembran – den Tonoplasten – vom Zytoplasma getrennt.
Nur in den jüngsten Pflanzenzellen füllt Zytoplasma den gesamten inneren Hohlraum aus. Während die Zelle wächst, verändert sich die räumliche Anordnung der zunächst zusammenhängenden Zytoplasmamasse deutlich: Es entstehen kleine, mit Zellsaft gefüllte Vakuolen und die gesamte Masse wird schwammig. Bei weiterem Zellwachstum verschmelzen einzelne Vakuolen und drängen die Zytoplasmaschichten an die Peripherie, wodurch die gebildete Zelle meist eine große Vakuole enthält und sich das Zytoplasma mit allen Organellen in der Nähe der Membran befindet.
Wasserlösliche organische und mineralische Verbindungen der Vakuolen bestimmen die entsprechenden osmotischen Eigenschaften lebender Zellen. Diese Lösung einer bestimmten Konzentration ist eine Art osmotische Pumpe für das kontrollierte Eindringen in die Zelle und die Freisetzung von Wasser, Ionen und Metabolitenmolekülen aus dieser.
In Kombination mit der Zytoplasmaschicht und ihren Membranen, die sich durch semipermeable Eigenschaften auszeichnen, bildet die Vakuole ein wirksames osmotisches System. Osmotisch bestimmt werden Indikatoren lebender Pflanzenzellen wie osmotisches Potenzial, Saugkraft und Turgordruck.

Struktur der Vakuole
Plastiden
Plastiden sind die größten (nach dem Zellkern) zytoplasmatischen Organellen, die nur den Zellen pflanzlicher Organismen innewohnen. Sie kommen nicht nur in Pilzen vor. Plastiden spielen eine wichtige Rolle im Stoffwechsel. Sie sind durch eine doppelte Membranhülle vom Zytoplasma getrennt und einige Arten verfügen über ein gut entwickeltes und geordnetes System innerer Membranen. Alle Plastiden sind vom gleichen Ursprung.
Chloroplasten- die häufigsten und funktionell wichtigsten Plastiden photoautotropher Organismen, die photosynthetische Prozesse durchführen, die letztendlich zur Bildung führen organische Substanz und Freisetzung von freiem Sauerstoff. Die Chloroplasten höherer Pflanzen besitzen einen Komplex Interne Struktur.

Chloroplastenstruktur
Die Größe der Chloroplasten in verschiedenen Pflanzen ist nicht gleich, aber im Durchschnitt beträgt ihr Durchmesser 4–6 Mikrometer. Chloroplasten können sich unter dem Einfluss der Bewegung des Zytoplasmas bewegen. Darüber hinaus wird unter dem Einfluss von Licht eine aktive Bewegung von Chloroplasten vom Amöbentyp in Richtung der Lichtquelle beobachtet.
Chlorophyll ist der Hauptstoff der Chloroplasten. Dank Chlorophyll grüne Pflanzen ist in der Lage, Lichtenergie zu nutzen.
Leukoplasten(farblose Plastiden) sind klar definierte zytoplasmatische Körper. Ihre Größe ist etwas kleiner als die Größe von Chloroplasten. Ihre Form ist auch gleichmäßiger und nähert sich der Kugelform.

Leukoplast-Struktur
Kommt in Epidermiszellen, Knollen und Rhizomen vor. Bei Beleuchtung verwandeln sie sich sehr schnell in Chloroplasten mit entsprechender Veränderung der inneren Struktur. Leukoplasten enthalten Enzyme, mit deren Hilfe aus überschüssiger Glukose, die bei der Photosynthese entsteht, Stärke synthetisiert wird, deren Großteil sich in Form von Stärkekörnern in Speichergeweben oder Organen (Knollen, Rhizome, Samen) ablagert. Bei manchen Pflanzen lagern sich Fette in Leukoplasten ab. Die Reservefunktion von Leukoplasten äußert sich gelegentlich in der Bildung von Reserveproteinen in Form von Kristallen oder amorphen Einschlüssen.
Chromoplasten In den meisten Fällen handelt es sich um Derivate von Chloroplasten, gelegentlich auch von Leukoplasten.

Chromoplast-Struktur
Die Reifung von Hagebutten, Paprika und Tomaten geht mit der Umwandlung von Chloro- oder Leukoplasten der Pulpazellen in Caratinoidplasten einher. Letztere enthalten überwiegend gelbe Plastidenpigmente – Carotinoide, die im reifen Zustand darin intensiv synthetisiert werden und farbige Lipidtröpfchen, feste Kügelchen oder Kristalle bilden. In diesem Fall wird Chlorophyll zerstört.
Mitochondrien
Mitochondrien sind für die meisten Pflanzenzellen charakteristische Organellen. Sie haben eine variable Form von Stäbchen, Körnern und Fäden. 1894 von R. Altman mit einem Lichtmikroskop entdeckt und die innere Struktur später mit einem Elektronenmikroskop untersucht.
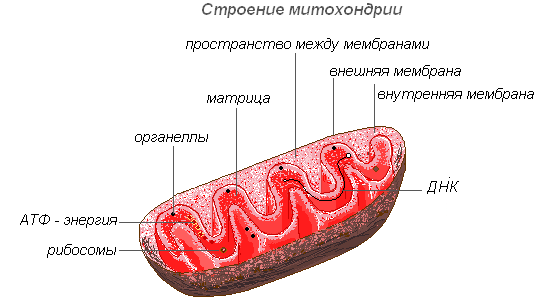
Die Struktur der Mitochondrien
Mitochondrien haben eine Doppelmembranstruktur. Die äußere Membran ist glatt, die innere bildet sich verschiedene Formen Auswüchse sind Röhren in Pflanzenzellen. Der Raum im Mitochondrium ist mit halbflüssigem Inhalt (Matrix) gefüllt, der Enzyme, Proteine, Lipide, Kalzium- und Magnesiumsalze, Vitamine sowie RNA, DNA und Ribosomen umfasst. Der enzymatische Komplex der Mitochondrien beschleunigt den komplexen und miteinander verbundenen Mechanismus biochemischer Reaktionen, die zur Bildung von ATP führen. In diesen Organellen werden Zellen mit Energie versorgt – die Energie chemischer Nährstoffbindungen wird im Prozess der Zellatmung in hochenergetische ATP-Bindungen umgewandelt. In den Mitochondrien erfolgt der enzymatische Abbau von Kohlenhydraten, Fettsäuren und Aminosäuren unter Freisetzung von Energie und deren anschließender Umwandlung in ATP-Energie. Die angesammelte Energie wird für Wachstumsprozesse, für neue Synthesen etc. aufgewendet. Mitochondrien vermehren sich durch Teilung und leben etwa 10 Tage, danach werden sie zerstört.
Endoplasmatisches Retikulum
Das endoplasmatische Retikulum ist ein Netzwerk aus Kanälen, Röhren, Vesikeln und Zisternen, die sich im Zytoplasma befinden. Es wurde 1945 vom englischen Wissenschaftler K. Porter entdeckt und ist ein Membransystem mit ultramikroskopischer Struktur.

Struktur des endoplasmatischen Retikulums
Das gesamte Netzwerk wird mit dem Äußeren zu einem Ganzen zusammengefasst Zellmembran Atomhülle. Es gibt glatte und raue ER, die Ribosomen tragen. Auf den Membranen des glatten ER befinden sich Enzymsysteme, die am Fett- und Kohlenhydratstoffwechsel beteiligt sind. Dieser Membrantyp überwiegt in Samenzellen, die reich an Speichersubstanzen (Proteine, Kohlenhydrate, Öle) sind; an der körnigen EPS-Membran sind Ribosomen befestigt, und bei der Synthese eines Proteinmoleküls wird die Polypeptidkette mit Ribosomen in den EPS-Kanal eingetaucht. Die Funktionen des endoplasmatischen Retikulums sind sehr vielfältig: Stofftransport sowohl innerhalb der Zelle als auch zwischen benachbarten Zellen; Aufteilung einer Zelle in separate Abschnitte, in denen verschiedene physiologische Prozesse und chemische Reaktionen gleichzeitig ablaufen.
Ribosomen
Ribosomen sind zelluläre Organellen ohne Membran. Jedes Ribosom besteht aus zwei Partikeln, deren Größe nicht identisch ist und die in zwei Fragmente geteilt werden können, die nach der Vereinigung zu einem ganzen Ribosom weiterhin die Fähigkeit zur Proteinsynthese behalten.
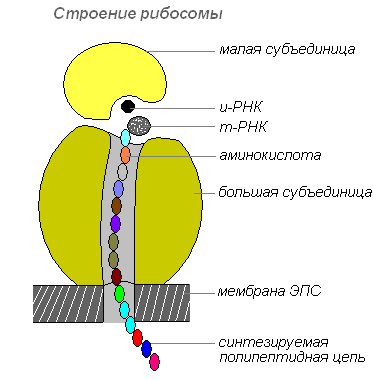
Ribosomenstruktur
Ribosomen werden im Zellkern synthetisiert, verlassen ihn dann und wandern in das Zytoplasma, wo sie an der Außenfläche der Membranen des endoplasmatischen Retikulums befestigt werden oder sich frei befinden. Abhängig von der Art des zu synthetisierenden Proteins können Ribosomen einzeln funktionieren oder zu Komplexen – Polyribosomen – zusammengefasst sein.
Die Zellwand ist ein Derivat des Protoplasten, d.h. entsteht während seiner Lebensaktivität (Abb. 61). Es verleiht der Zelle eine bestimmte Form, schützt den Protoplasten und verhindert durch Widerstand gegen den intrazellulären Druck den Zellbruch. Zellwände erfüllen die Funktionen des inneren Skeletts einer Pflanze und verleihen ihren Organen die notwendige mechanische Festigkeit.
Zellwände lassen Sonnenlicht gut durch und Wasser und darin gelöste Mineralien bewegen sich leicht durch sie hindurch. Zwischen den Wänden benachbarter Zellen gibt es Mittelplatte - Pektinschicht, die eigentlich eine interzelluläre Substanz ist und die Wände benachbarter Zellen zusammenhält. An den Stellen, an denen sich die Zellwände benachbarter Zellen nicht schließen, sind sie mit Wasser gefüllt Interzellularräume. Als Prozess der Zerstörung der Interzellularsubstanz wird bezeichnet, wodurch die Wände benachbarter Zellen getrennt werden Mazeration. Es ist eine natürliche Mazeration zu beobachten
Reis. 61.
A- Diagramm der Struktur der Zellwand; B- Diagramm der Beteiligung des Golgi-Apparats am Aufbau der Zellwand; IN- detaillierter Aufbau der Zellwand: 1 - Mittelplatte; 2 - es ist Zeit; 3 - Sekundärwand;
- 4 - Primärwand; 5 - Diktyosom; 6 - Golgi-Vesikel;
- 7-Plasmalemma; 8- Zellenwand; 9- Makrofibrille;
- 10-Mikrofibrille; 11 - Mizelle; 12 - Zellulosemolekül;
- 13 – Struktur eines Cellulosemolekülfragments
in überreifen Früchten von Äpfeln, Ebereschen, Melonen usw. Eine künstliche Mazeration wird beispielsweise beim Einweichen von Flachsstielen durchgeführt, um Bastfasern von ihnen zu trennen; Es tritt auch bei der Wärmebehandlung von Früchten auf.
Die Zellwand enthält Polysaccharide: Pektine, Hemizellulose und Zellulose. Sehr lange Cellulosemoleküle sind geordnet parallel zueinander angeordnet (jeweils 40–60) und bilden sich Mizellen. Mizellen werden in Bündeln gesammelt - Mikrofibrillen, stellt die Grundstruktureinheit der Cellulose dar. Mikrofibrillen wiederum werden zu kombiniert Makrofibrillen- sehr dünne Fasern ungewisser Länge. Zellulosemakrofibrillen werden in stark gewässertes Wasser eingetaucht Matrix, bestehend aus Pektinen, Hemizellulosen und einigen anderen Stoffen. Die Festigkeit der Zellwand wird durch elastische Zellulose-Mikrofibrillen gegeben, deren Zugfestigkeit der von Stahl nahekommt. Die Stärke und Elastizität der Zellwand liegt ihrer Fähigkeit zugrunde, sich reversibel zu dehnen. Dank Pektinen und Hemizellulose ist die Zellwand sehr wasserdurchlässig – Wasser und darin gelöste Stoffe wandern problemlos von Zelle zu Zelle.
Die Zellwand grenzt außen an das Plasmalemma an, das aktiv an seinem Wachstum teilnimmt. Moleküle aus Pektinen, Hemizellulose, Zellulose und anderen Substanzen werden synthetisiert und in den Dictyosomen-Zisternen des Golgi-Apparats angesammelt. Golgi-Vesikel transportieren sie zur Peripherie des Protoplasten – zum Plasmalemma. An der Kontaktstelle zwischen Vesikel und Plasmalemma löst sich dieses auf und der Inhalt der Vesikel, sobald er sich außerhalb des Plasmalemmas befindet, wird zum Aufbau der Zellwand verwendet. Die Vesikelmembran stellt nicht nur die Integrität des Plasmalemmas wieder her, sondern sorgt auch für dessen Oberflächenwachstum. Das Wachstum der Zellwand erfolgt aufgrund der enzymatischen Aktivität des Plasmalemmas.
Als Wände werden die sich teilenden und wachsenden Zellen bezeichnet primär. Sie enthalten viel Wasser (60-90 %), ihre Trockenmasse wird von Pektinen und Hemizellulose dominiert – der Zelluloseanteil beträgt nicht mehr als 30 %. Wenn sich eine Zelle in der Telophase der Mitose teilt, wird die Mutterzelle durch die Bildung eines Septums in ihrer Äquatorialebene in zwei Tochterzellen geteilt – Mittelplatte. Auf beiden Seiten der Mittelschicht beginnt jede der beiden Tochterzellen, ihre eigene primäre Zellwand zu bilden. Das Wachstum der Mittelplatte und der Primärwände der beiden Tochterzellen erfolgt in zentrifugaler Richtung – vom Zentrum der Mutterzelle bis zu ihrer Peripherie. Die Mittelplatte ist sehr dünn und besteht aus Pektin.
Die durch die Teilung entstehende neue Zelle beginnt zu wachsen und ihr Volumen kann sich um das Hundertfache oder mehr vergrößern. Das Zellwachstum erfolgt hauptsächlich durch Verstauchungen aufgrund der Aufnahme von Wasser und der Vergrößerung des Vakuolenvolumens. Durch den entstehenden Innendruck dehnt sich die Primärwand, in die Cellulosemicellen, Pektine und Hemicellulose leicht eingebettet werden. Methode des Zellwandwachstums nach Implementierung Baumaterial zwischen bestehenden Bauwerken genannt Invagination.
In der primären Zellwand gibt es zunächst dünnere Bereiche, in denen die Zellulosefibrillen lockerer angeordnet sind – primäre Porenfelder. Die primären Porenfelder der Wände zweier benachbarter Zellen fallen normalerweise zusammen. Hier wandern die Tubuli des endoplasmatischen Retikulums von einer Zelle zur anderen – Plasmodesmen. Die Wege, auf denen Plasmodesmen von einer Zelle zur anderen gelangen, werden genannt plasmodesminale Tubuli. Durch diese Tubuli sind die Hyaloplasmen benachbarter Zellen miteinander verbunden. Plasmodesmen übernehmen den interzellulären Transport von Stoffen (Hormone, Aminosäuren, ATP, Zucker usw.). Die mit Hilfe von Plasmodesmen zu einem Ganzen vereinten Protoplasten von Körperzellen werden Symplasten genannt. Als Transport von Stoffen entlang von Plasmodesmen wird bezeichnet symplastisch.(Man nennt die Gesamtheit der Zellwände, der Mittelschicht und der Interzellularräume Apoplast, geht auf ihnen apoplastisch Transport von Stoffen.)
Nachdem das Zellwachstum abgeschlossen ist, kann seine Primärwand dünn bleiben (bei Bildungsgewebezellen) oder beginnen, dicker zu werden (bei permanenten Gewebezellen). Das Wachstum der Zellwanddicke nennt man sekundäre Verdickung. Dadurch lagert es sich an der Innenfläche der Primärwand ab. Sekundärwand was wächst Apposition- Aufbringen von Zellulosemizellen auf eine bestehende Wand. In diesem Fall befinden sich die jüngsten Schichten der sekundären Zellwand neben dem Plasmalemma. Die sekundäre Zellwand übernimmt vor allem tragende und mechanische Funktionen. Es enthält erheblich weniger Wasser als in der Primärmasse, und in der Trockenmasse überwiegt Zellulose (bis zu 50 %). Beispielsweise kann in den Sekundärwänden einzelliger Haare aus Baumwoll- und Flachsbastfasern der Zellulosegehalt bis zu 95 % betragen.
Die sekundäre Verdickung der Zellwand erfolgt ungleichmäßig. Abschnitte der sekundären Zellwand an den Stellen der primären Porenfelder bleiben in der Regel unverdickt. Solche nicht verdickten Bereiche der Zellwand nennt man manchmal. Poren in den Wänden zweier benachbarter Zellen fallen in der Regel zusammen und bilden sich ein paar Mal. Der durch ein Porenpaar gebildete Porenkanal ist verstopft Verschlussfilm der Pore - ein Septum, bestehend aus einer mittleren Lamina und zwei Primärwänden benachbarter Zellen. Der Verschlussfilm der Pore wird von zahlreichen Plasmodesmen durchdrungen, durch die Plasmodesmen hindurchtreten.
Poren werden unterschieden einfach Und begrenzt(Abb. 62). Bei einfachen Poren ist der Durchmesser ihres Porenkanalabschnitts über die gesamte Länge gleich, d. h. es hat eine zylindrische Form. Einfache Poren sind typisch für Parenchymzellen. Umrandete Poren sind charakteristisch für die Wände von Zellen, die Wasser mit gelösten Mineralien leiten – Tracheiden und Gefäßsegmente. Bei solchen Poren hat ihr Teil des Porenkanals die Form eines Trichters, dessen breite Seite an den Verschlussfilm der Pore angrenzt.
In den Zellen leitender Gewebe von Nadelbäumen ist der Porenverschlussfilm nur an den Rändern wasserdurchlässig, da sein zentraler scheibenförmiger Teil verdickt und verholzt ist Torus - lässt kein Wasser durch. Der Torus übernimmt die Rolle einer Klappe. Ist der Wasserdruck in benachbarten Zellen nicht gleich, wird der Schließfilm abgelenkt und der Torus blockiert die Bewegung des Wassers durch den Porenkanal.

Reis. 62.
A- einfach; B- begrenzt; IN- halbkantig:
1 - Abschlussfilm; 2 - Porenkanal; 3 - Torus
Zusätzlich zu den Poren befinden sich in den Wänden wasserleitender Zellen Zähnung- Durchgangslöcher (Gefäßsegmente, wasserspeichernde Zellen von Torfmoos).
Veränderungen der Zellwand. Abhängig von den Funktionen der Zelle kann ihre Wand durch die Ablagerung von Substanzen in ihr verändert werden. Seine üblichen Modifikationen sind: Verholzung, Suberisierung, Kutinisierung, Mineralisierung und Schleimbildung.
Zellwandverholzung, oder Verholzung, entsteht durch die Ablagerung von Lignin in den intermicellaren Räumen - einer Substanz aromatischer Natur mit einem Komplex chemische Struktur. Die Festigkeit und Härte der Wand nehmen zu, ihre Elastizität nimmt jedoch ab. Verholzte Wände sind in der Lage, Wasser und Luft durchzulassen. Bei einer verholzten Zellwand bleibt der Protoplast der Zelle möglicherweise am Leben, stirbt jedoch normalerweise ab. Bei manchen Gehölzen reichert sich bis zu 30 % Lignin im Holz an. Lignin kann sich auch in den Zellwänden alternder Grastriebe ansammeln, was deren Futterwert deutlich verringert und den Zeitpunkt der Heuernte bestimmt. Bei der Gewinnung von Zellulosezellstoff aus Holz, das für die Papierherstellung notwendig ist, wird eine künstliche Entholzung durchgeführt. Eine natürliche Verholzung der Zellwand ist möglich, aber selten.
Probenahme, oder Suberinisierung, - Ablagerung einer persistenten, fettähnlichen, amorphen Substanz Suberin (hydrophobes Polymer) in der Zellwand. Suberisierte Zellwände sind für Gase und Wasser undurchlässig, was zum Absterben des Protoplasten führt. Zellen mit suberisierten Wänden schützen Pflanzen zuverlässig vor Wasserverlust, extremen Temperaturen, pathogenen Bakterien und Pilzen.
Kutinisierung - Ablagerung von Cutin (einer ähnlichen Substanz) in den Zellwänden chemische Zusammensetzung zu Suberin). Cutin wird normalerweise in den oberflächlichen Schichten der Außenwände von Zellen und auf deren Oberflächen abgelagert. In Form eines Films – der Kutikula – bedeckt es beispielsweise die Oberfläche der Zellen des Hautgewebes – der Epidermis.
Mineralisierung Die Zellwand wird durch die Ablagerung von Kalzium- und Kieselsäuresalzen zerstört. Diese Stoffe verleihen der Wand Härte und Sprödigkeit. Der Mineralisierungsprozess kommt besonders gut in den Zellwänden der Epidermis von Getreide-, Seggen- und Schachtelhalmsprossen zum Ausdruck. Aus diesem Grund empfiehlt es sich, die Triebe von Seggen und Gräsern vor der Blüte zu mähen – später werden sie durch die starke Mineralisierung gröber, was die Qualität des Heus beeinträchtigt.
Schleim- Umwandlung von Zellulose und Zellwandpektinen in spezielle Polysaccharide - Schleim und Zahnfleisch, die bei Kontakt mit Wasser stark aufquellen können. Ein Abrutschen der Wand wird bei Samenschalenzellen beobachtet, beispielsweise bei Quitte, Flachs, Gurke und Wegerich. Klebriger Schleim kann die Samenverbreitung fördern (Wegerich); Wenn die Samen keimen, absorbieren und speichern die Schleimstoffe Wasser und schützen sie so vor dem Austrocknen. In der Wurzelkappe fungiert Schleim als Gleitmittel und erleichtert den Durchgang der Wurzel zwischen Erdklumpen. Schleim und Zahnfleisch können in erheblichen Mengen entstehen, wenn sich Zellwände aufgrund ihrer Schädigung auflösen. Kirschen und Pflaumen bilden oft Gummi, wenn Zweige und Stämme verletzt werden. Der sogenannte Kirschkleber ist ein in Form von Perlen aushärtender Kaugummi, der die Oberfläche von Wunden und Frostlöchern bedeckt und so das Eindringen von Infektionen verhindert. Schleim dieser Art nennt man Gumose und gilt als pathologisches Phänomen.
Da sekundäre Zellwände als inneres Skelett der Pflanze fungieren, verleihen sie ihren Organen die nötige Festigkeit (was besonders wichtig ist für Land Pflanzen), oft sind sie in der Lage, sich lokal oder vollständig deutlich zu verdicken, um dem Gewebe und damit dem Pflanzenorgan eine größere Festigkeit zu verleihen. Durch die Ablagerung von Zellulose kommt es zu einer Verdickung der Zellwand.
Die Funktionen von Zellen werden oft ausschließlich von ihren Wänden übernommen, da die Protoplasten der Zellen absterben. Dies betrifft die Korkzellen,

Reis. 63.
Tracheiden, Gefäßsegmente, mechanische Gewebefasern. Das Holz, das die meisten riesigen Baumstämme einnimmt, besteht beispielsweise überwiegend aus verholzten Zellwänden, deren Protoplasten längst abgestorben sind.
Zellwände spielen in unserem Leben eine große Rolle. Aus ihnen werden textile Rohstoffe (Baumwollsamenhaare, Flachsfasern etc.) und Rohstoffe für die Herstellung von Seilen und Schnüren (Hanffasern, Seilfasern, Sisal etc.) gewonnen. Aus Zellwänden gewonnene Zellulose wird zur Herstellung von Papier (Fichten- und Espenholz), Acetatseide, Viskose, Kunststoffen, Zellophan und vielem mehr verwendet. Kork ist ein Gewebe aus abgestorbenen Zellen mit suberisierten Wänden und wird seit langem als wertvolles wasser- und luftdichtes Wärmedämmmaterial verwendet und wird zunehmend im modernen Bauwesen eingesetzt.
Die pflanzliche Zellwand wird unter Beteiligung der Plasmamembran gebildet und ist ein extrazelluläres (extrazelluläres) mehrschichtiges Gebilde, das die Zelloberfläche schützt und als äußeres Skelett der Pflanzenzelle dient (Abb. 1). Die pflanzliche Zellwand besteht aus zwei Komponenten: einer amorphen gelartigen Kunststoffmatrix (Basis) mit hohem Wassergehalt und einem unterstützenden Fibrillensystem. Zusätzliche Polymersubstanzen und Salze, die häufig in der Zusammensetzung der Schalen enthalten sind, verleihen ihnen Steifheit und machen sie nicht benetzbar.
Reis. 1. Schema des Aufbaus der pflanzlichen Zellwand: 0 – Mittelplatte; 1 – Primärschale (zwei Schichten auf beiden Seiten von 0); 2 – Schichten der Sekundärhülle; 3 - Tertiärschale; PM – Plasmamembran; B - Vakuole; Ich bin der Kern
Chemisch gesehen gehören die Hauptbestandteile der Pflanzenhülle zu den strukturellen Polysacchariden. Die Matrix der Pflanzenhüllen umfasst heterogene Gruppen von Polysacchariden, die sich in konzentrierten Alkalien, Hemizellulosen und Pektinsubstanzen lösen. Hemicellulosen sind verzweigte Polymerketten bestehend aus verschiedenen Hexosen (Glucose, Mannose, Galactose etc.), Pentosen (Xylose, Arabinose) und Uronsäuren (Glucuronsäure und Galacturonsäure). Diese Bestandteile der Hemicellulosen verbinden sich in unterschiedlichen Mengenverhältnissen miteinander und bilden verschiedene Kombinationen. Ketten aus Hemicellulosemolekülen kristallisieren nicht und bilden keine Elementarfibrillen. Aufgrund der polaren Gruppen der Uronsäuren sind sie stark hydratisiert.
Pektinstoffe sind eine heterogene Gruppe, zu der verzweigte, stark hydratisierte Polymere gehören, die aufgrund vieler Galacturonsäurereste negative Ladungen tragen. Aufgrund der Eigenschaften ihrer Bestandteile ist die Matrix eine mit Fibrillen verstärkte weiche Kunststoffmasse.
Die faserigen Bestandteile pflanzlicher Zellwände bestehen normalerweise aus Cellulose, einem linearen, nicht verzweigten Polymer aus Glucose. Das Molekulargewicht von Cellulose variiert zwischen 5·104 und 5·105, was 300–3000 Glucoseresten entspricht. Solche linearen Cellulosemoleküle können zu Bündeln oder Fasern zusammengefasst werden. In der Zellwand bildet Cellulose Fibrillen, die aus bis zu 25 nm dicken submikroskopischen Mikrofibrillen bestehen, die wiederum aus vielen parallelen Ketten von Cellulosemolekülen bestehen.
Die Mengenverhältnisse von Zellulose zu Matrixsubstanzen (Hemizellulose) können bei verschiedenen Objekten sehr unterschiedlich sein. Über 60 % der Trockenmasse der Primärschalen besteht aus ihrer Matrix und etwa 30 % aus der Skelettsubstanz – Zellulose. In rohen Zellmembranen ist fast das gesamte Wasser mit Hemicellulosen verbunden, sodass die Masse der Hauptsubstanz im gequollenen Zustand 80 % des Nassgewichts der gesamten Membran erreicht, während der Gehalt an Faserstoffen auf nur 12 % reduziert ist. In Baumwollhaaren beträgt der Zelluloseanteil 90 %; In Holz macht Zellulose 50 % der Zellwandbestandteile aus.
Zur Zusammensetzung der Zellwände gehören neben Zellulose, Hemizellulose und Pektinen zusätzliche Komponenten, was ihnen besondere Eigenschaften verleiht. So führt die Verkrustung (Einbau) der Membranen mit Lignin (Polymer von Coniferylalkohol) zur Verholzung der Zellwände und erhöht so deren Festigkeit (Abb. 2). Lignin vermischt in solchen Schalen die plastischen Stoffe der Matrix und übernimmt die Rolle des Hauptstoffes mit hoher Festigkeit. Oftmals wird die Matrix mit Mineralien (SiO2, CaCO3 etc.) verstärkt.

Reis. 2. Verkrustung der Zellmembran: a - fibrilläres Gerüst und interfibrilläre Matrix; b – mit Lignin verkrustete Schale mit Matrixresten, die ihre Dehnfähigkeit verloren hat; c – nachträgliche Einlagerung mit Phenolen und/oder Mineralien, was zu einer Erhöhung der Schalenhärte führt
Auf der Oberfläche der Zellmembran können sich verschiedene verkrustende Substanzen wie Cutin und Suberin ansammeln, was zur Suberisierung der Zellen führt. In Epidermiszellen lagert sich Wachs auf der Oberfläche der Zellmembranen ab und bildet eine wasserdichte Schicht, die verhindert, dass die Zelle Wasser verliert.
Aufgrund ihrer porösen, lockeren Struktur ist die pflanzliche Zellwand weitgehend durchlässig für niedermolekulare Verbindungen wie Wasser, Zucker und Ionen. Makromoleküle dringen jedoch nur schlecht in Zellulosehüllen ein: Die Größe der Poren in den Hüllen, die die freie Diffusion von Substanzen ermöglichen, beträgt nur 3–5 nm.
Experimente mit markierten Verbindungen haben gezeigt, dass beim Wachstum der Zellmembran die Stoffe, aus denen sie aufgebaut ist, über die gesamte Zelloberfläche freigesetzt werden. Amorphe Matrixsubstanzen, Hemicellulosen und Pektine werden in den Vakuolen des Golgi-Apparats synthetisiert und durch Exozytose durch das Plasmalemma freigesetzt. Cellulosefibrillen werden durch spezielle Enzyme synthetisiert, die im Plasmalemma eingebaut sind.
Die Membranen differenzierter, reifer Zellen sind meist mehrschichtig, in den Schichten sind die Zellulosefibrillen unterschiedlich ausgerichtet und auch ihre Anzahl kann stark schwanken. Typischerweise werden primäre, sekundäre und tertiäre Zellwände beschrieben (siehe Abb. 1). Um den Aufbau und das Aussehen dieser Membranen zu verstehen, ist es notwendig, sich mit ihrer Entstehung nach der Zellteilung vertraut zu machen.
Wenn sich Pflanzenzellen nach einer Chromosomendivergenz teilen, entsteht in der Äquatorialebene der Zellen eine Ansammlung kleiner Membranvesikel, die im zentralen Teil der Zellen miteinander zu verschmelzen beginnen (Abb. 3). Dieser Prozess der Verschmelzung kleiner Vakuolen erfolgt vom Zentrum der Zelle bis zur Peripherie und setzt sich fort, bis die Membranvesikel miteinander und mit der Plasmamembran der Seitenoberfläche der Zelle verschmelzen. Auf diese Weise entsteht eine Zellplatte oder ein Phragmoplast. In seinem zentralen Teil befindet sich eine amorphe Matrixsubstanz, die die verschmelzenden Vesikel füllt. Es ist erwiesen, dass diese primären Vakuolen aus den Membranen des Golgi-Apparats stammen. Die primäre Zellwand enthält auch eine kleine Menge Protein (ca. 10 %), das reich an Hydroxyprolin ist und viele kurze Oligosaccharidketten aufweist, was dieses Protein als Glykoprotein definiert. Entlang der Peripherie der Zellplatte ist bei Betrachtung im polarisierten Licht eine deutliche Doppelbrechung zu erkennen, die dadurch verursacht wird, dass sich an dieser Stelle ausgerichtete Zellulosefibrillen befinden. Somit besteht die wachsende primäre Zellwand bereits aus drei Schichten: der zentralen – der mittleren Platte, die nur aus einer amorphen Matrix besteht, und zwei peripheren – der Primärmembran, die Hemizellulose und Zellulosefibrillen enthält. Wenn die mittlere Schicht ein Produkt der Aktivität der Elternzelle ist, wird die Primärmembran durch die Sekretion von Hemizellulose und Zellulosefibrillen durch zwei neue Zellkörper gebildet. Und jede weitere Zunahme der Dicke der Zellwand (oder besser gesagt der Interzellularwand) erfolgt aufgrund der Aktivität zweier Tochterzellen, die von gegenüberliegenden Seiten Substanzen der Zellmembran absondern, die sich durch die Schichtung immer neuer Schichten verdickt. Die Freisetzung von Matrixsubstanzen erfolgt von Anfang an dadurch, dass sich Vesikel des Golgi-Apparats der Plasmamembran nähern, sie mit der Membran verschmelzen und ihren Inhalt über das Zytoplasma hinaus freisetzen. Hier, außerhalb der Zelle, auf ihrer Plasmamembran, findet die Synthese und Polymerisation von Cellulosefibrillen statt. So entsteht nach und nach eine sekundäre Zellmembran. Es ist schwierig, die Primärschale von der Sekundärschale mit ausreichender Genauigkeit zu bestimmen und unterscheiden zu können, da sie durch mehrere Zwischenschichten miteinander verbunden sind.
Die Hauptmasse der Zellwand, die ihre Bildung abgeschlossen hat, ist die Sekundärmembran. Sie gibt ihr den Käfig Finale Form. Nachdem sich die Zelle in zwei Tochterzellen geteilt hat, wachsen neue Zellen, ihr Volumen nimmt zu und ihre Form verändert sich; Zellen verlängern sich oft. Gleichzeitig kommt es zu einer Dickenzunahme der Zellmembran und einer Umstrukturierung ihrer inneren Struktur.
Wenn die primäre Zellmembran gebildet wird, sind in ihrer Zusammensetzung noch wenige Cellulosefibrillen vorhanden, und diese befinden sich mehr oder weniger senkrecht zur zukünftigen Längsachse der Zelle. Später, während der Elongationsperiode (Verlängerung der Zelle durch das Wachstum von Vakuolen im Zytoplasma), erfährt die Ausrichtung dieser quer gerichteten Fibrillen passive Veränderungen: Die Fibrillen beginnen, sich im rechten Winkel zueinander anzuordnen und enden schließlich länglich mehr oder weniger parallel zur Längsachse der Zelle. Der Prozess läuft ständig weiter: In den alten Schichten (näher an der Mitte der Schale) unterliegen die Fibrillen passiven Verschiebungen, und die Ablagerung neuer Fibrillen in den inneren Schichten (am nächsten an der Zellmembran) wird gemäß dem ursprünglichen Design der Zelle fortgesetzt Hülse. Dieser Prozess schafft die Möglichkeit, dass Fibrillen relativ zueinander gleiten, und die Umstrukturierung der Zellmembranverstärkung ist aufgrund des gelatinösen Zustands der Komponenten ihrer Matrix möglich. Wenn anschließend Hemizellulose in der Matrix durch Lignin ersetzt wird, nimmt die Beweglichkeit der Fibrillen stark ab, die Schale wird dichter und es kommt zur Verholzung.
Reis. 3. Schema des Zellmembranwachstums vom Beginn während der Zellteilung (I) bis zur vollständigen Reifung (V): 1 – Primärmembran; 2 – Schichten der Sekundärhülle; 3 - Tertiärschale; B - Vakuolen; SP – Mittelplatte; PM – Plasmamembranen zweier benachbarter Zellen
Unter der Sekundärhülle befindet sich häufig eine Tertiärhülle, die als getrockneter Rückstand der degenerierten Schicht des Zytoplasmas selbst betrachtet werden kann.
Es ist zu beachten, dass bei der Zellteilung von Pflanzen der Bildung der Primärmembran nicht in allen Fällen die Bildung einer Zellplatte vorausgeht. So entstehen bei der Grünalge Spirogyra durch die Bildung von Vorsprüngen an den Seitenwänden der ursprünglichen Zelle neue Querwände, die, allmählich zur Zellmitte hin wachsend, die Zelle schließen und in zwei Teile teilen.
Wie bereits erwähnt, kommt es, wenn einer Zelle in einer wässrigen, hypotonischen Umgebung ihre Membran entzogen wird, zur Lyse, also zum Bruch der Zelle. Es stellte sich heraus, dass es durch die Auswahl geeigneter Konzentrationen von Salzen und Zuckern möglich ist, den osmotischen Druck außerhalb und innerhalb von Zellen ohne Membran auszugleichen. In diesem Fall erwerben solche Protoplasten Kugelform(Sphäroplasten). Wenn in der Umgebung, in der sich die Protoplasten befinden, eine ausreichende Menge an Nährstoffen und Salzen (darunter Ca2+) vorhanden ist, stellen die Zellen ihre Zellmembran wieder her und regenerieren sie. Darüber hinaus sind sie in Gegenwart von Hormonen (Auxinen) in der Lage, sich zu teilen und Zellkolonien zu bilden, die zum Wachstum der gesamten Pflanze, aus der die Zelle entnommen wurde, führen können.
Der Hauptfaserbestandteil der Zellwand großer Pilzgruppen (Basibiomyceten, Ascomyceten, Zygomyceten) ist Chitin; Es handelt sich um ein Polysaccharid, dessen Hauptsaccharid N-Acetylglucosamin ist. Die Zusammensetzung der Zellwand von Pilzen kann neben Chitin auch Matrixsubstanzen, Glykoproteine usw. umfassen verschiedene Proteine, im Zytoplasma synthetisiert und von der Zelle nach außen abgegeben.
Neue Hülle entsteht bei der Zellteilung im Endstadium der Mitose – Telophase. Nach der Divergenz der Chromosomen erscheint in der Äquatorialebene der Zellen eine Ansammlung kleiner Membranvesikel, die im zentralen Teil der Zellen miteinander zu verschmelzen beginnen. Dieser Prozess der Verschmelzung kleiner Vakuolen erfolgt vom Zentrum der Zelle bis zur Peripherie und setzt sich fort, bis die Membranvesikel miteinander und mit der Plasmamembran der Seitenoberfläche der Zelle verschmelzen. So entsteht es Zellplatte oder Phragmoplast.
IN Hauptteil Es enthält eine amorphe Matrixsubstanz, die die verschmelzenden Vesikel füllt. Es ist nachgewiesen, dass diese primären Vakuolen ihren Ursprung haben Membranen des Golgi-Apparats. Die primäre Zellwand enthält auch eine kleine Menge Protein (ca. 10 %), das reich an Hydroxyprolin ist und viele kurze Oligosaccharidketten aufweist, was dieses Protein als Glykoprotein definiert.
Nach der Bildung der Mittelplatte lagert der Protoplast benachbarter Zellen die Primärmembran darauf ab. Die Zelluloseschicht, die sich während des Zellwachstums ablagert, wird als Zelluloseschicht bezeichnet primäre Zellmembran.
Primärschalen enthalten neben Zellulose, Hemizellulose und Pektin auch Strukturprotein – Glykoprotein. Primärschalen können auch verholzt sein, Lignin ist für sie jedoch in der Regel nicht charakteristisch. Der charakteristischste Teil ist jedoch die Primärschale Pektinkomponente. Es verleiht der Schale Plastizität und ermöglicht es ihr, sich mit der Länge der Organe zu dehnen: Wurzel, Stängel, Blatt. Pektinstoffe sind stark quellfähig, daher enthalten die Primärschalen viel Wasser (60-90 %). Hemizellulosen und Pektinstoffe machen 50-60 % des Trockengewichts der Primärhülle aus, der Zellulosegehalt überschreitet nicht 30 %, Strukturprotein beträgt bis zu 10 %.
Der fortlaufende Prozess der Freisetzung von Matrixsubstanzen erfolgt durch Annäherung an die Plasmamembran der Vesikel des Golgi-Apparats, deren Verschmelzung mit der Membran und die Freisetzung ihres Inhalts über das Zytoplasma hinaus. Hier, außerhalb der Zelle, auf ihrer Plasmamembran, findet die Synthese und Polymerisation von Cellulosefibrillen statt. So entsteht es nach und nach sekundäre Zellwand. Es ist schwierig, die Primärschale von der Sekundärschale mit ausreichender Genauigkeit zu bestimmen und zu unterscheiden, da sie durch mehrere miteinander verbunden sind Zwischenschichten.
Schüttgut Sobald die Zellwand ihre Bildung abgeschlossen hat, bildet sie die Sekundärmembran. Es gibt der Zelle ihre endgültige Form. Nachdem sich die Zelle in zwei Tochterzellen geteilt hat, wachsen neue Zellen, ihr Volumen nimmt zu und ihre Form verändert sich; Zellen verlängern sich oft. Gleichzeitig kommt es zu einer Dickenzunahme der Zellmembran und einer Umstrukturierung ihrer inneren Struktur.
Während der Dehnungsphase beginnen die Fibrillen, sich im rechten Winkel zueinander anzuordnen, und verlängern sich schließlich mehr oder weniger parallel zur Längsachse der Zelle. Der Prozess läuft ständig weiter: In den alten Schichten (näher an der Mitte der Schale) unterliegen die Fibrillen passiven Verschiebungen, und die Ablagerung neuer Fibrillen in den inneren Schichten (am nächsten an den Zellmembranen) wird gemäß dem ursprünglichen Design der Schale fortgesetzt Hülse. Dieser Prozess schafft die Möglichkeit, dass Fibrillen relativ zueinander gleiten, und die Umstrukturierung der Zellmembranverstärkung ist aufgrund des gelatinösen Zustands der Komponenten ihrer Matrix möglich. Anschließend wird Hemizellulose in der Matrix durch Lignin ersetzt Die Beweglichkeit der Fibrillen nimmt stark ab, die Schale wird dichter und es kommt zu einer Verholzung. Inhalt verschiedene Substanzen Ungefähr so: sehr wenig Wasser, Cellulose 40–50 %, Lignin 25–30 %, Hemicellulose 20–30 % und praktisch keine Pektinstoffe.
Die Sekundärschale ist nicht immer gleichmäßig verteilt. In einigen spezialisierten wasserleitenden Zellen sieht es so aus Ringe oder Spiralbänder. Solche Zellen behalten die Fähigkeit, sich auch nach dem Tod in Längsrichtung auszudehnen.
Wird oft unter der Sekundärhülle gefunden Tertiärschale, der als getrockneter Überrest der degenerierten Schicht des Zytoplasmas selbst betrachtet werden kann.
Zellwand (Zellmembran)- ein charakteristisches Merkmal einer Pflanzenzelle, das sie von einer tierischen Zelle unterscheidet. Die Zellwand gibt der Zelle ihre spezifische Form. Auf speziellen Nährmedien kultivierte Pflanzenzellen, bei denen die Wand enzymatisch entfernt wird, nehmen immer eine Kugelform an. Die Zellwand verleiht der Zelle Festigkeit und schützt den Protoplasten; sie gleicht den Turgordruck aus und verhindert so den Bruch des Plasmalemmas. Die Ansammlung von Zellwänden bildet das innere Skelett, das den Pflanzenkörper stützt und ihm mechanische Festigkeit verleiht.
Die Zellwand ist farblos und transparent und lässt Sonnenlicht leicht durch. Normalerweise sind die Wände mit Wasser gesättigt. Das Zellwandsystem transportiert Wasser und darin gelöste niedermolekulare Verbindungen (Transport entlang des Apoplasten).
Die Zellwand besteht hauptsächlich aus Polysacchariden, die in unterteilt werden können Skelettsubstanzen Und Matrixsubstanzen.
Skelettsubstanz pflanzliche Zellwand ist Zellulose (Faser), das ein Beta-1,4-D-Glucan ist. Dies ist die häufigste organische Substanz in der Biosphäre. Cellulosemoleküle sind sehr lange unverzweigte Ketten, sie liegen in Gruppen von mehreren Dutzend parallel zueinander und werden durch zahlreiche Wasserstoffbrückenbindungen zusammengehalten. Infolge, Mikrofibrillen, die den strukturellen Rahmen der Wand bilden und ihre Festigkeit bestimmen. Zellulose-Mikrofibrillen sind nur mit einem Elektronenmikroskop sichtbar, ihr Durchmesser beträgt 10–30 nm und ihre Länge erreicht mehrere Mikrometer.
Cellulose ist unlöslich und quillt nicht in Wasser. Es ist chemisch sehr inert und löst sich nicht in organischen Lösungsmitteln, konzentrierten Laugen und verdünnten Säuren. Zellulose-Mikrofibrillen sind elastisch und sehr zugfest (ähnlich wie Stahl). Diese Eigenschaften bestimmen die weit verbreitete Verwendung von Zellulose und ihren Produkten. Die weltweite Produktion von Baumwollfasern, die fast ausschließlich aus Zellulose bestehen, beträgt 1,5 · 10 7 Tonnen pro Jahr. Aus Zellulose werden rauchfreies Schießpulver, Acetatseide und Viskose, Zellophan und Papier gewonnen. Qualitative Reaktion durchgeführt auf Zellulose mit einem Reagenz Chlor-Zink-Jod, die Zellwand der Zellulose färbt sich blauviolett.
Bei Pilzen handelt es sich um die Skelettsubstanz der Zellwand Chitin– ein Polysaccharid, das aus Glucosaminresten aufgebaut ist. Chitin ist noch haltbarer als Zellulose.
Mikrofibrillen werden amorph eingetaucht Matrix, normalerweise ein wassergesättigtes Kunststoffgel. Die Matrix ist eine komplexe Mischung aus Polysacchariden, deren Moleküle aus Resten mehrerer verschiedener Zucker bestehen und kürzere und verzweigte Ketten als Cellulose sind. Matrixpolysaccharide bestimmen Eigenschaften der Zellwand wie starke Quellung, hohe Durchlässigkeit für Wasser und darin gelöste niedermolekulare Verbindungen sowie Kationenaustauscheigenschaften. Matrixpolysaccharide werden in zwei Gruppen eingeteilt – Pektinstoffe Und Hemizellulosen.
Pektische Substanzen stark quellen oder sich in Wasser auflösen. Sie werden durch Laugen und Säuren leicht zerstört. Die einfachsten Vertreter der Pektinstoffe sind wasserlöslich Pektinsäuren– Polymerisationsprodukte von Alpha-D-Galacturonansäure (bis zu 100 Einheiten), verbunden durch 1,4-Bindungen zu linearen Ketten (Alpha-1,4-D-Galacturonan). Pektinsäuren (Pektine)– Hierbei handelt es sich um Polymerverbindungen der Alpha-D-Galacturonsäure mit höherem Molekulargewicht (100–200 Einheiten), bei denen die Carboxylgruppen teilweise methyliert sind. Pektate Und Pektinate– Calcium- und Magnesiumsalze von Pektin und Pektinsäuren. Pektinsäuren, Pektate und Pektinate sind in Gegenwart von Zuckern und organischen Säuren in Wasser löslich und bilden dichte Gele.
Die Zellwände von Pflanzen enthalten hauptsächlich Protopektine– hochmolekulare Polymere aus methoxylierter Polygalacturonsäure mit Arabinanen und Galactanen; in zweikeimblättrigen Pflanzen enthalten die Galacturonanketten eine geringe Menge Rhamnose. Protopektine sind in Wasser unlöslich.
Hemizellulosen sind verzweigte Ketten, die aus neutralen Zuckerresten aufgebaut sind; häufiger sind Glucose, Galactose, Mannose und Xylose; Polymerisationsgrad 50-300. Hemicellulosen sind chemisch stabiler als Pektinstoffe, sie lassen sich schwerer hydrolysieren und quellen in Wasser weniger leicht auf. Hemizellulosen können als Reservestoffe in den Zellwänden von Samen eingelagert werden (Dattelpalme, Kaki). Pektinstoffe und Hemizellulosen sind durch gegenseitige Übergänge verbunden. Neben Polysacchariden ist in der Zellwandmatrix ein spezielles Strukturprotein vorhanden. Es ist an Arabinose-Zuckerreste gebunden und somit ein Glykoprotein.
Matrixpolysaccharide füllen nicht nur die Räume zwischen Cellulose-Mikrofibrillen. Ihre Ketten sind geordnet angeordnet und bilden sowohl untereinander als auch mit Mikrofibrillen zahlreiche Bindungen, was die Festigkeit der Zellwand deutlich erhöht.
Pflanzliche Zellwände unterliegen häufig chemischen Veränderungen. Verholzung, oder Verholzung tritt auf, wenn es in der Matrix abgelagert wird Lignin– eine wasserunlösliche Polymerverbindung phenolischer Natur. Die verholzte Zellwand verliert ihre Elastizität, ihre Härte und Druckfestigkeit nehmen stark zu und ihre Wasserdurchlässigkeit nimmt ab. Reagenzien für Lignin sind: 1) Phloroglucinol Und konzentrierte Salzsäure oder Schwefelsäure(verholzte Wände bekommen eine kirschrote Farbe) und 2) Sulfat Anilin, unter dessen Einfluss die verholzten Wände zitronengelb werden. Die Verholzung ist charakteristisch für die Zellwände des leitenden Gewebes des Xylems (Holz) und des mechanischen Gewebes des Sklerenchyms.
Probenahme, oder Suberinisierung entsteht durch Ablagerung hydrophober Polymere auf der Innenseite der Zellwand - suberina Und Wachs. Suberin ist eine Mischung aus Estern polymerer Fettsäuren. Wachsmonomere sind Fettalkohole und Wachsester. Das Wachs lässt sich leicht durch organische Lösungsmittel extrahieren und schmilzt schnell unter Bildung von Kristallen. Suberin ist eine amorphe Verbindung, die in organischen Lösungsmitteln weder schmilzt noch sich auflöst. Suberin und Wachs, die abwechselnd parallele Schichten bilden, kleiden den gesamten Zellhohlraum von innen in Form eines Films aus. Der Suberinfilm ist praktisch undurchlässig für Wasser und Gase, sodass die Zelle nach seiner Bildung normalerweise abstirbt. Die Suberisierung ist charakteristisch für die Zellwände des Kork-Integumentargewebes. Das Reagenz für suberisierte Zellwände ist SudanIII, orangerote Farbe.
Coutinisierung Die Außenwände der epidermalen Gewebezellen werden freigelegt. Kutin Und Wachs in abwechselnden Schichten auf der Außenfläche der Zellwand in Form eines Films abgelagert - Nagelhaut. Cutin ist eine fettähnliche Polymerverbindung, die in ihrer chemischen Natur und ihren Eigenschaften Suberin ähnelt. Die Nagelhaut schützt die Pflanze vor übermäßiger Verdunstung von Wasser von der Pflanzenoberfläche. Sie können es mit einem Reagenz färben SudanIII in orange-roter Farbe.
Mineralisierung Zellwand entsteht durch Ablagerung in der Matrix große Menge Mineralstoffe, am häufigsten Kieselsäure (Siliziumoxid), seltener Calciumoxalat und Carbonat. Mineralien verleihen der Wand Härte und Zerbrechlichkeit. Die Ablagerung von Kieselsäure ist charakteristisch für die Epidermiszellen von Schachtelhalmen, Seggen und Gräsern. Die durch die Verkieselung erworbene Steifheit der Stängel und Blätter dient als Schutzmittel gegen Schnecken und verringert zudem die Schmackhaftigkeit und den Nährwert der Pflanzen erheblich.
Einige spezialisierte Zellen haben Schleim Zellenwand. In diesem Fall werden anstelle einer Zellulose-Sekundärwand amorphe, hochhydratisierte saure Polysaccharide in der Form eingelagert Schleim Und Zahnfleisch, in seiner chemischen Natur den Pektinsubstanzen ähnlich. Schleim löst sich gut in Wasser und bildet Schleimlösungen. Zahnfleisch ist klebrig und dehnt sich zu Fäden aus. Im trockenen Zustand haben sie eine geile Konsistenz. Bei der Ablagerung von Schleim wird der Protoplast allmählich in Richtung der Zellmitte gedrückt, sein Volumen und das Volumen der Vakuole nehmen allmählich ab. Schließlich kann es sein, dass sich die Zellhöhle vollständig mit Schleim füllt und die Zelle stirbt. In einigen Fällen kann Schleim durch die primäre Zellwand an die Oberfläche gelangen. Der Golgi-Apparat spielt eine wichtige Rolle bei der Synthese und Sekretion von Schleim.
Von Pflanzenzellen abgesonderter Schleim erfüllt verschiedene Funktionen. Somit dient der Schleim der Wurzelkappe als Gleitmittel und erleichtert das Wachstum der Wurzelspitze im Boden. Die Schleimdrüsen insektenfressender Pflanzen (Sonnentau) scheiden Fangschleim aus, an dem Insekten haften bleiben. Der von den äußeren Zellen der Samenschale (Lein, Quitte, Wegerich) abgesonderte Schleim fixiert den Samen an der Bodenoberfläche und schützt den Sämling vor dem Austrocknen. Schleim wird mit einem Reagenz gefärbt Methylenblau in blauer Farbe.
Die Freisetzung von Zahnfleisch erfolgt normalerweise, wenn Pflanzen verletzt werden. Beispielsweise wird bei Kirschen und Pflaumen häufig Zahnfleischausfluss aus verletzten Bereichen von Stämmen und Ästen beobachtet. Kirschleim ist ein gehärteter Gummi. Das Zahnfleisch erfüllt eine Schutzfunktion, indem es die Wunde von der Oberfläche her abdeckt. Zahnfleisch wird hauptsächlich in Gehölzen aus der Familie der Hülsenfrüchte (Akazie, Traganth-Astragalus) und Rosaceae der Unterfamilie Pflaume (Kirsche, Pflaume, Aprikose) gebildet. Zahnfleisch und Schleimstoffe werden in der Medizin verwendet.
Die Zellwand ist ein Produkt der lebenswichtigen Aktivität des Protoplasten. Im Golgi-Apparat werden Matrixpolysaccharide, Wandglykoprotein, Lignin und Schleim gebildet. Die Cellulosesynthese, Bildung und Ausrichtung der Mikrofibrillen erfolgt durch das Plasmalemma. Eine wichtige Rolle bei der Ausrichtung der Mikrofibrillen spielen Mikrotubuli, die parallel zu den abgelagerten Mikrofibrillen in der Nähe des Plasmalemmas liegen. Werden Mikrotubuli zerstört, entstehen nur isodiametrische Zellen.
Die Zellwandbildung beginnt während der Zellteilung. In der Teilungsebene bildet sich eine Zellplatte, eine einzige Schicht, die den beiden Tochterzellen gemeinsam ist. Es besteht aus Pektinsubstanzen mit halbflüssiger Konsistenz; es gibt keine Zellulose. In einer erwachsenen Zelle bleibt die Zellplatte erhalten, erfährt jedoch Veränderungen, weshalb sie so genannt wird Median, oder Interzellularplatte (interzelluläre Substanz) (Reis. 2.16). Die Mittelplatte ist meist sehr dünn und kaum zu unterscheiden.
Unmittelbar nach der Bildung der Zellplatte beginnen die Protoplasten der Tochterzellen mit dem Aufbau einer eigenen Zellwand. Es lagert sich von innen sowohl auf der Oberfläche der Zellplatte als auch auf der Oberfläche anderer Zellwände ab, die zuvor zur Mutterzelle gehörten. Nach der Teilung tritt die Zelle in die Elongationswachstumsphase ein, die durch eine intensive osmotische Wasseraufnahme durch die Zelle verursacht wird, die mit der Bildung und dem Wachstum der zentralen Vakuole verbunden ist. Der Turgordruck beginnt die Wand zu dehnen, sie reißt jedoch nicht, da sich ständig neue Anteile von Mikrofibrillen und Matrixsubstanzen in ihr ablagern. Die Ablagerung neuer Materialanteile erfolgt gleichmäßig über die gesamte Oberfläche des Protoplasten, sodass die Dicke der Zellwand nicht abnimmt.
Als Wände werden die sich teilenden und wachsenden Zellen bezeichnet primär. Sie enthalten viel (60-90 %) Wasser. In der Trockenmasse dominieren Matrixpolysaccharide (60–70 %), der Zellulosegehalt beträgt nicht mehr als 30 % und es ist kein Lignin enthalten. Die Dicke der Primärwand ist sehr gering (0,1–0,5 Mikrometer).
Bei vielen Zellen hört die Zellwandablagerung gleichzeitig mit dem Aufhören des Zellwachstums auf. Solche Zellen sind bis zum Lebensende von einer dünnen Primärwand umgeben ( Reis. 2.16).
Reis. 2.16. Parenchymzelle mit Primärwand.
In anderen Zellen setzt sich die Wandablagerung fort, selbst nachdem die Zelle ihre endgültige Größe erreicht hat. In diesem Fall nimmt die Wandstärke zu und das vom Zellhohlraum eingenommene Volumen ab. Dieser Vorgang wird aufgerufen sekundäre Verdickung Wände, und die Wand selbst wird genannt sekundär(Reis. 2.17).
Die Sekundärwand kann als zusätzliche Wand betrachtet werden, die hauptsächlich eine mechanische Stützfunktion übernimmt. Es ist die Sekundärwand, die für die Eigenschaften von Holz, Textilfasern und Papier verantwortlich ist. Die Sekundärwand enthält deutlich weniger Wasser als die Primärwand; Es wird von Cellulose-Mikrofibrillen (40-50 % des Trockenmassegewichts) dominiert, die parallel zueinander angeordnet sind. Von den Matrixpolysacchariden sind Hemizellulosen (20–30 %) typisch, Pektinstoffe sind sehr selten. Sekundäre Zellwände verholzen normalerweise. In nicht verholzten Sekundärwänden (Flachsbastfasern, Baumwollhaare) kann der Zellulosegehalt bis zu 95 % betragen. Der hohe Gehalt und die streng geordnete Ausrichtung der Mikrofibrillen bestimmen die Höhe mechanische Eigenschaften Nebenwände. Oftmals sterben Zellen mit einer sekundären verholzten Zellwand ab, nachdem die sekundäre Verdickung abgeschlossen ist.
Die mittlere Lamina klebt benachbarte Zellen zusammen. Wird es aufgelöst, verlieren die Zellwände den Kontakt zueinander und trennen sich. Dieser Vorgang wird aufgerufen Mazeration. Weit verbreitet ist die natürliche Mazeration, bei der die Pektinstoffe der Mittelplatte mithilfe des Enzyms Pektinase in einen löslichen Zustand überführt und anschließend mit Wasser abgewaschen werden (überreife Früchte von Birne, Melone, Pfirsich, Banane). Häufig wird eine teilweise Mazeration beobachtet, bei der sich die Mittelplatte nicht auf der gesamten Oberfläche, sondern nur in den Ecken der Zellen auflöst. Aufgrund des Turgordrucks werden benachbarte Zellen an diesen Stellen abgerundet, was zur Bildung führt Interzellularräume(Reis. 2.16). Die Interzellularräume bilden ein einziges verzweigtes Netzwerk, das mit Wasserdampf und Gasen gefüllt ist. Somit verbessern Interzellularräume den Gasaustausch der Zellen.
Ein charakteristisches Merkmal der Sekundärwand ist ihre ungleichmäßige Ablagerung auf der Primärwand, wodurch unverdickte Bereiche in der Sekundärwand verbleiben - Poren. Wenn die Sekundärwand keine große Dicke erreicht, sehen die Poren wie kleine Vertiefungen aus. In Zellen mit einer starken Sekundärwand sehen die Poren im Querschnitt wie radiale Kanäle aus, die sich vom Zellhohlraum bis zur Primärwand erstrecken. Basierend auf der Form des Porenkanals gibt es zwei Arten von Poren: einfach und über kantig(Abb. 2.17).

Reis. 2.17. Porentypen: A – Zellen mit Sekundärwänden und zahlreichen einfachen Poren; B – ein Paar einfacher Poren; B – Paar begrenzter Poren.
U einfache Poren Der Durchmesser des Porenkanals ist über seine gesamte Länge gleich und hat die Form eines schmalen Zylinders. Einfache Poren sind charakteristisch für Parenchymzellen, Bast- und Holzfasern.
Poren in zwei benachbarten Zellen neigen dazu, einander gegenüber zu erscheinen. Diese gemeinsame Poren Sie sehen aus wie ein Kanal, getrennt durch eine dünne Trennwand der Mittelplatte und der Primärwand. Diese Verbindung zweier Poren benachbarter Wände benachbarter Zellen nennt man Porenpaare und funktioniert als Ganzes. Der sie trennende Wandabschnitt wird genannt Abschlussfilm Poren, oder Porenmembran. In lebenden Zellen ist der Verschlussfilm der Pore zahlreich durchdrungen Plasmodesmen(Reis. 2.18).
Plasmodesmen kommen nur in Pflanzenzellen vor. Es handelt sich um Zytoplasmastränge, die die Wand benachbarter Zellen durchqueren. Die Anzahl der Plasmodesmen in einer Zelle ist sehr groß – von mehreren Hundert bis Zehntausenden; normalerweise werden Plasmodesmen in Gruppen gesammelt. Der Durchmesser des plasmodesmalen Kanals beträgt 30–60 nm. Seine Wände sind mit Plasmalemma ausgekleidet, das mit dem Plasmalemma benachbarter Zellen übergeht. Im Zentrum der Plasmodesmen befindet sich ein Membranzylinder – Zentralstange Plasmodesmen, kontinuierlich mit den Membranen der Elemente des endoplasmatischen Retikulums beider Zellen. Zwischen dem zentralen Stab und der Plasmamembran im Kanal befindet sich Hyaloplasma, das mit dem Hyaloplasma benachbarter Zellen übergeht.

Reis. 2.18. Plasmodesmen unter dem Elektronenmikroskop (Diagramm): 1 – im Längsschnitt; 2 – im Querschnitt; Pl– Plasmalemma; CA– zentraler Stab aus Plasmodesmen; ER– Element des endoplasmatischen Retikulums.
Somit sind Zellprotoplasten nicht vollständig voneinander isoliert, sondern kommunizieren über Plasmodesmata-Kanäle. Sie übernehmen den interzellulären Transport von Ionen und kleinen Molekülen und übertragen auch hormonelle Reize. Durch Plasmodesmen bilden Protoplasten von Zellen in einem pflanzlichen Organismus ein einziges Ganzes namens simplast, und der Stofftransport durch Plasmodesmen wird genannt symplastisch im Gegensatz zu apoplastisch Transport entlang der Zellwände und Interzellularräume.
U begrenzte Poren(Reis. 2.17) Der Kanal verengt sich während der Ablagerung der Zellwand stark, sodass die innere Öffnung der Pore, die in den Zellhohlraum mündet, viel enger ist als die äußere, die an der Primärwand anliegt. Umrandete Poren sind charakteristisch für früh absterbende Zellen wasserleitender Holzelemente. Bei ihnen erweitert sich der Porenkanal trichterförmig zum Verschlussfilm hin und die Sekundärwand hängt walzenförmig über dem erweiterten Teil des Kanals und bildet eine Porenkammer. Der Name „umrandete Pore“ kommt daher, dass das innere Loch von der Oberfläche aus wie ein kleiner Kreis oder schmaler Schlitz aussieht, während das äußere Loch das innere Loch in Form eines Kreises mit größerem Durchmesser oder breiterem Durchmesser zu begrenzen scheint Schlitz.
Poren erleichtern den Transport von Wasser und gelösten Stoffen von Zelle zu Zelle, ohne die Festigkeit der Zellwand zu verringern.