Mitoza celulară. Diviziunea mitotică
Citeste si
Manual pentru clasele 10-11
Secțiunea II. Reproducerea și dezvoltarea organismelor
Capitolul V. Reproducerea organismelor
În fiecare secundă pe Pământ, un număr astronomic de ființe vii mor din cauza bătrâneții, a bolilor și a prădătorilor și numai datorită reproducerii, această proprietate universală a organismelor, viața pe Pământ nu se oprește.
Poate părea că procesele de reproducere la ființe vii sunt foarte diverse, dar toate pot fi reduse la două forme: asexuată și sexuală. Unele organisme au forme diferite reproducere. De exemplu, multe plante pot fi înmulțite prin butași, stratificare, tuberculi (reproducere asexuată) și semințe (sexual).
În timpul reproducerii sexuale, fiecare organism se dezvoltă dintr-o celulă, formată din fuziunea a două celule germinale - masculin și feminin.
Baza reproducerii și dezvoltării individuale a corpului este procesul de diviziune celulară.
§ 20. Diviziunea celulară. Mitoză
Abilitatea de a se diviza este cea mai importantă proprietate a celulelor. Fără diviziune, este imposibil de imaginat o creștere a numărului de ființe unicelulare, dezvoltarea unui organism multicelular complex dintr-un singur ou fecundat, reînnoirea celulelor, țesuturilor și chiar a organelor pierdute în timpul vieții organismului.
Diviziunea celulară se realizează în etape. La fiecare etapă de divizare au loc anumite procese. Acestea duc la dublarea materialului genetic (sinteza ADN) și la distribuția acestuia între celulele fiice. Perioada de viață a unei celule de la o diviziune la alta se numește ciclu celular.
Pregătirea pentru împărțire. Organismele eucariote, formate din celule cu nuclei, încep să se pregătească pentru diviziune într-un anumit stadiu al ciclului celular, în interfază.
În perioada de interfază în celulă are loc procesul de biosinteză a proteinelor, cromozomii se dublează. De-a lungul cromozomului original din compușii chimici prezenți în celulă, copia sa exactă este sintetizată, molecula de ADN este dublată. Un cromozom dublat este format din două jumătăți - cromatide. Fiecare cromatidă conține o moleculă de ADN.
Interfaza in celulele vegetale si animale dureaza in medie 10-20 de ore.Apoi vine procesul de diviziune celulara - mitoza.
În timpul mitozei, celula trece printr-o serie de faze succesive, în urma cărora fiecare celulă fiică primește același set de cromozomi ca și în celula mamă.
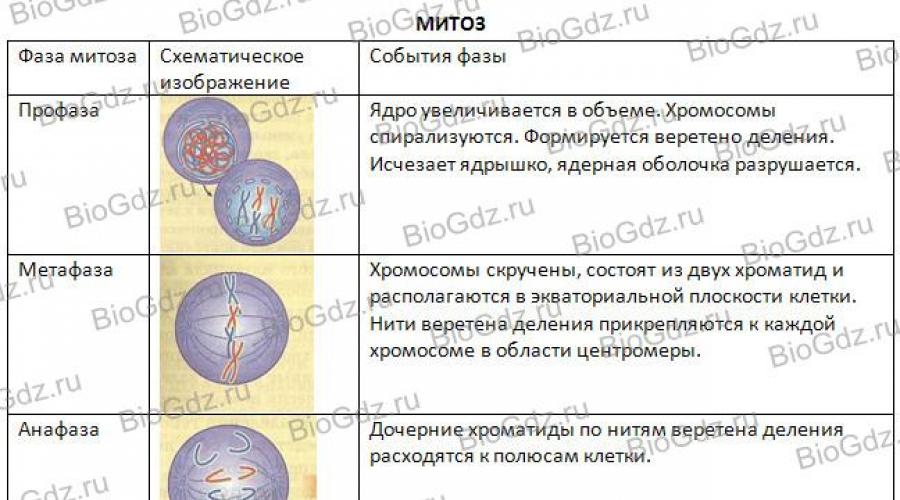
fazele mitozei. Există patru faze ale mitozei: profază, metafază, anafază și telofază. Figura 29 prezintă schematic cursul mitozei. În profază, centriolii sunt clar vizibili - formațiuni situate în centrul celulei și joacă un rol în divergența cromozomilor fiice ai animalelor. (Reamintim că doar unele plante au centrioli în centrul celulei, care organizează divergența cromozomilor.) Vom lua în considerare mitoza folosind exemplul celulă animală, deoarece prezența unui centriol face procesul de segregare a cromozomilor mai vizibil. Centriolii se dublează și diverg către diferiți poli ai celulei. Microtubulii se extind de la centrioli, formând fibre fusiforme, care reglează divergența cromozomilor către polii celulei în diviziune.
Orez. 29. Schema mitozei
La sfârșitul profazei, membrana nucleară se dezintegrează, nucleolul dispare treptat, cromozomii se spiralează și ca urmare se scurtează și se îngroașă și pot fi deja observați la microscop cu lumină. Ele sunt și mai bine văzute în următoarea etapă a mitozei - metafaza.
În metafază, cromozomii sunt localizați în planul ecuatorial al celulei. Se vede clar că fiecare cromozom, format din două cromatide, are o constricție - un centromer. Cromozomii sunt atașați de fibrele fusului prin centromerii lor. După divizarea centromerului, fiecare cromatidă devine un cromozom fiică independent.
Apoi urmează următoarea etapă a mitozei - anafaza, în timpul căreia cromozomii fiice (cromatidele unui cromozom) diverg către diferiți poli ai celulei.
Următoarea etapă a diviziunii celulare este telofaza. Începe după ce cromozomii fiice, formați dintr-o cromatidă, au ajuns la polii celulei. În acest stadiu, cromozomii se despiralizează din nou și capătă aceeași formă pe care o aveau înainte de a începe diviziunea celulară în interfază (filamente lungi și subțiri). În jurul lor ia naștere o înveliș nuclear, iar în nucleu se formează un nucleol, în care sunt sintetizați ribozomii. În procesul de diviziune a citoplasmei, toate organelele (mitocondrii, complexul Golgi, ribozomi etc.) sunt distribuite mai mult sau mai puțin uniform între celulele fiice.
Astfel, ca urmare a mitozei, dintr-o celulă se obțin două celule, fiecare având un număr și o formă caracteristică de cromozomi pentru un anumit tip de organism și, în consecință, o cantitate constantă de ADN.
Întregul proces de mitoză durează în medie 1-2 ore.Durata sa este oarecum diferită pt tipuri diferite celule. Depinde si de conditii Mediul extern(temperatură, regim de lumină și alți indicatori).
Semnificația biologică a mitozei constă în faptul că asigură constanta numărului de cromozomi din toate celulele corpului. În procesul de mitoză, ADN-ul cromozomilor celulei mamă este distribuit strict egal între cele două celule fiice care provin din aceasta. Ca rezultat al mitozei, toate celulele fiice primesc aceeași informație genetică.
- Ce modificări ale celulei preced diviziunea?
- Când se formează axul? Care este rolul lui?
- Descrieți fazele mitozei și descrieți pe scurt cum are loc acest proces.
- Ce este o cromatidă? Când devine un cromozom?
- Ce este un centromer? Ce rol joacă în mitoză?
- Ce este semnificație biologică mitoză?
Amintiți-vă din cursul de botanică, zoologie, anatomie, fiziologie și igiena umană cum are loc reproducerea în lumea organică.
1. Dați definiții conceptelor.
Interfaza- faza de pregătire pentru diviziunea mitotică, când are loc duplicarea ADN-ului.
Mitoză- aceasta este o diviziune, în urma căreia există o distribuție strict identică a cromozomilor exact copiați între celulele fiice, ceea ce asigură formarea de celule identice genetic.
Ciclu de viață
- perioada de viață celulară din momentul apariției acesteia în procesul de diviziune până la moarte sau sfârșitul diviziunii ulterioare.
2. Cum diferă creșterea organismelor unicelulare de creșterea celor pluricelulare?
Creșterea unui organism unicelular este o creștere a dimensiunii și complicarea structurii unei singure celule, iar creșterea organismelor multicelulare este, de asemenea, o diviziune activă a celulelor - o creștere a numărului lor.
3. De ce există neapărat o interfază în ciclul de viață al unei celule?
În interfaza are loc pregătirea pentru diviziunea și duplicarea ADN-ului. Dacă nu s-ar întâmpla, atunci cu fiecare diviziune celulară, numărul de cromozomi s-ar înjumătăți și destul de curând nu ar mai rămâne deloc cromozomi în celulă.
4. Completați grupul „Fazele mitozei”.
5. Folosind figura 52 din § 3.4, completați tabelul.

6. Compuneți un syncwin pentru termenul „mitoză”.
Mitoză
Cu patru faze, uniformă
Împarte, distribuie, împarte
Oferă material genetic celulelor fiice
diviziune celulara.
7. Stabiliți o corespondență între fazele ciclului mitotic și evenimentele care au loc în ele.
faze
1. Anafaza
2. Metafaza
3. Interfaza
4. Telofază
5. Profaza
Evenimente
A. Celula crește, se formează organele, ADN-ul se dublează.
B. Cromatidele se separă și devin cromozomi independenți.
B. Începe spiralizarea cromozomilor, învelișul nuclear este distrus.
D. Cromozomii sunt localizați în planul ecuatorial al celulei. Fibrele fusului se atașează de centromeri.
D. Fusul diviziunii dispare, se formează membrane nucleare, se desfășoară cromozomii.
8. De ce terminarea mitozei - diviziunea citoplasmei are loc diferit în celulele animale și vegetale?
Nu în celulele animale perete celular, ei au membrana celularaîmpinge spre interior, iar celula se divide prin constricție.
În celulele vegetale, membrana se formează în plan ecuatorial în interiorul celulei și, extinzându-se la periferie, împarte celula în jumătate.
9. De ce interfaza durează mult mai mult timp în ciclul mitotic decât diviziunea în sine?
În timpul interfazei, celula se pregătește intens pentru mitoză, în ea au loc procese de sinteză, ADN-ul se dublează, celula crește, trece prin ea. ciclu de viață, fără a include diviziunea în sine.
10. Alegeți răspunsul corect.
Testul 1
Ca rezultat al mitozei, o celulă diploidă produce:
4) 2 celule diploide.
Testul 2
Diviziunea centromerului și divergența cromatidelor la polii celulei are loc în:
3) anafaza;
Testul 3
Ciclul de viață este:
2) viața celulară de la diviziune până la sfârșitul următoarei diviziuni sau moarte;
Testul 4
Care termen este scris greșit?
4) telofaza.
11. Explicați originea și sens general cuvânt (termen), bazat pe sensul rădăcinilor care îl alcătuiesc.

12. Alegeți un termen și explicați cum sensul său modern corespunde sensului inițial al rădăcinilor sale.
Termenul ales este interfaza.
Corespondenţă. Termenul corespunde și înseamnă perioada dintre fazele mitozei, când are loc pregătirea pentru divizare.
13. Formulează și notează ideile principale de la § 3.4.
Ciclul de viață este viața unei celule de la diviziune până la sfârșitul următoarei diviziuni sau moarte. Între diviziuni, celula se pregătește pentru aceasta în timpul interfazei. În acest moment, există o sinteză de substanțe, duplicarea ADN-ului.
Celula se divide prin mitoză. Este format din 4 etape:
Profaza.
Metafaza.
Anafaza.
Telofază.
Scopul mitozei: ca urmare a acesteia, din 1 celulă mamă se formează 2 celule fiice cu un set identic de gene. Cantitatea de material genetic și cromozomi rămâne aceeași, asigurând stabilitatea genetică a celulelor.
Cea mai importantă componentă a ciclului celular este ciclul mitotic (proliferativ). Este un complex de fenomene interconectate și coordonate în timpul diviziunii celulare, precum și înainte și după aceasta. Ciclul mitotic este un set de procese care au loc într-o celulă de la o diviziune la alta și se termină cu formarea a două celule din generația următoare. În plus, conceptul de ciclu de viață include și perioada de performanță de către celulă a funcțiilor sale și perioadele de odihnă. În acest moment, soarta ulterioară a celulei este incertă: celula poate începe să se dividă (intră în mitoză) sau să înceapă să se pregătească pentru a îndeplini funcții specifice.
Stadiile principale ale mitozei
1.
Reduplicarea (autodublarea) informației genetice a celulei mamă și distribuția uniformă a acesteia între celulele fiice. Aceasta este însoțită de modificări ale structurii și morfologiei cromozomilor, în care se concentrează mai mult de 90% din informațiile unei celule eucariote.
2.
Ciclul mitotic constă din patru perioade succesive: G1 presintetic (sau postmitotic), S sintetic, G2 postsintetic (sau premitotic) și mitoza propriu-zisă. Ele constituie interfaza autocatalitică (perioada pregătitoare).
Fazele ciclului celular:
1) presintetice (G1). Apare imediat după diviziunea celulară. Sinteza ADN-ului nu a avut loc încă. Celula crește în mod activ în dimensiune, stochează substanțele necesare divizării: proteine (histone, proteine structurale, enzime), ARN, molecule de ATP. Există o diviziune a mitocondriilor și a cloroplastelor (adică, structuri capabile de autoreproducție). Caracteristicile organizării celulei de interfază sunt restaurate după diviziunea anterioară;
2)
sintetice (S). Materialul genetic este duplicat prin replicarea ADN-ului. Apare într-un mod semi-conservator, când dublu helix al moleculei de ADN diverge în două catene și pe fiecare dintre ele este sintetizată o catenă complementară.
Rezultatul sunt două identice elice duble ADN, fiecare dintre ele constând dintr-un nou și lanț vechi ADN. Cantitatea de material ereditar este dublată. În plus, sinteza de ARN și proteine continuă. De asemenea, o mică parte a ADN-ului mitocondrial suferă replicare (partea sa principală este replicată în perioada G2);
3) postsintetic (G2). ADN-ul nu se mai sintetizează, dar există o corectare a neajunsurilor făcute în timpul sintezei sale în perioada S (reparare). De asemenea, acumulează energie și nutrienți, sinteza de ARN și proteine (în principal nucleare) continuă.
S și G2 sunt direct legate de mitoză, așa că uneori sunt izolate într-o perioadă separată - preprofază.
Aceasta este urmată de mitoză în sine, care constă din patru faze. Procesul de divizare include mai multe faze succesive și este un ciclu. Durata sa este diferită și variază de la 10 la 50 de ore în majoritatea celulelor. În același timp, în celulele corpului uman, durata mitozei în sine este de 1-1,5 ore, perioada de interfază G2 este de 2-3 ore, Perioada S a interfazei este de 6-10 ore.
Durata etapelor individuale este diferită și variază în funcție de tipul de țesut, stare fiziologică organism, factori externi. Cele mai lungi etape sunt asociate cu procesele de sinteză intracelulară: profază și telofază. Cele mai trecătoare faze ale mitozei, în timpul cărora are loc mișcarea cromozomilor: metafaza și anafaza. Procesul real de divergență a cromozomilor către poli nu depășește de obicei 10 minute.
Profaza
Principalele evenimente ale profazei includ condensarea cromozomilor în nucleu și formarea unui fus de fisiune în citoplasma celulei. Dezintegrarea nucleolului în profază este o trăsătură caracteristică, dar nu obligatorie pentru toate celulele.
În mod convențional, momentul apariției cromozomilor vizibili microscopic datorită condensării cromatinei intranucleare este luat drept început de profază. Compactarea cromozomilor are loc datorită helixării pe mai multe niveluri a ADN-ului. Aceste modificări sunt însoțite de o creștere a activității fosforilazelor care modifică histonele care sunt direct implicate în asamblarea ADN-ului. Ca rezultat, activitatea transcripțională a cromatinei scade brusc, genele nucleolare sunt inactivate și majoritatea proteinelor nucleolare se disociază. Cromatidele surori condensate în profaza timpurie rămân împerecheate pe toată lungimea lor cu ajutorul proteinelor cohesine, cu toate acestea, până la începutul prometafazei, legătura dintre cromatide este păstrată doar în regiunea centromerului. Până la profaza târzie, se formează cinetocori maturi pe fiecare centromer al cromatidelor surori, care sunt necesare pentru ca cromozomii să se atașeze de microtubulii fusului în prometafază.
Odată cu procesele de condensare intranucleară a cromozomilor, fusul mitotic începe să se formeze în citoplasmă - una dintre principalele structuri ale aparatului de diviziune celulară responsabilă de distribuția cromozomilor între celulele fiice. La formarea fusului de diviziune în toate celulele eucariote, iau parte corpurile polare, microtubulii și cinetocorii cromozomilor.
Odată cu începutul formării fusului mitotic în profază, sunt asociate modificări dramatice ale proprietăților dinamice ale microtubulilor. Timpul de înjumătățire al unui microtubul mediu scade de aproximativ 20 de ori de la 5 minute la 15 secunde. Cu toate acestea, rata de creștere a acestora crește de aproximativ 2 ori în comparație cu microtubulii din aceeași interfază. Capetele de polimerizare plus sunt „instabile dinamic” și trec brusc de la creșterea uniformă la scurtarea rapidă, care adesea depolimerizează întregul microtubul. Este de remarcat faptul că, pentru buna funcționare a fusului mitotic, este necesar un anumit echilibru între procesele de asamblare și depolimerizare a microtubulilor, deoarece nici microtubulii fusului stabilizați, nici depolimerizați nu sunt capabili să miște cromozomii.
Odată cu modificările observate în proprietățile dinamice ale microtubulilor care alcătuiesc filamentele fusului, în profază se formează poli de fisiune. Centrozomii replicați în faza S diverg în direcții opuse datorită interacțiunii microtubulilor polari care cresc unul spre celălalt. Cu capetele lor minus, microtubulii sunt scufundați în substanța amorfă a centrozomilor, iar procesele de polimerizare se desfășoară din partea capetelor plus orientată spre planul ecuatorial al celulei. În același timp, mecanismul probabil al divergenței polilor este explicat astfel: proteinele dineinei sunt orientate în direcție paralelă polimerizarea capetele plus ale microtubulilor polilor și proteinele asemănătoare kinezinei, la rândul lor, le împing către polii de diviziune.
In paralel cu condensarea cromozomilor si formarea fusului mitotic, in timpul profaza are loc fragmentarea reticulului endoplasmatic, care se desface in mici vacuole, care apoi divergente spre periferia celulei. În același timp, ribozomii pierd contactul cu membranele ER. Cisternele aparatului Golgi își schimbă și ele localizarea perinucleară, dezintegrându-se în dictiozomi separați, distribuiți în citoplasmă fără o ordine specială.
prometafaza
Sfârșitul profazei și debutul prometafazei sunt de obicei marcate de dezintegrarea membranei nucleare. Întreaga linie proteinele laminei sunt fosforilate, în urma căreia învelișul nuclear este fragmentat în vacuole mici, iar complexele porilor dispar. După distrugerea membranei nucleare, cromozomii sunt aranjați aleatoriu în regiunea nucleului. Cu toate acestea, în curând toți încep să se miște.
În prometafază, se observă mișcare intensivă, dar aleatorie a cromozomilor. Inițial, cromozomii individuali se deplasează rapid spre cel mai apropiat pol al fusului mitotic cu o viteză de până la 25 µm/min. În apropierea polilor de diviziune, crește probabilitatea interacțiunii capetelor plus nou sintetizate ale microtubulilor fusului cu cinetocorii cromozomi. Ca urmare a acestei interacțiuni, microtubulii cinetocori sunt stabilizați din depolimerizarea spontană, iar creșterea lor asigură parțial distanța cromozomului conectat la ei în direcția de la pol la planul ecuatorial al fusului. Pe de altă parte, cromozomul este depășit de fire de microtubuli care vin de la polul opus al fusului mitotic. Interacționând cu kinetocorul, ei participă și la mișcarea cromozomului. Ca rezultat, cromatidele surori sunt asociate cu poli opuși ai fusului. Forța dezvoltată de microtubuli de la diferiți poli nu numai că stabilizează interacțiunea acestor microtubuli cu cinetocorii, dar și, în cele din urmă, aduce fiecare cromozom în planul plăcii metafazate.
În celulele de mamifere, prometafaza are loc, de regulă, în 10-20 de minute. La neuroblastele de lăcuste, această etapă durează doar 4 minute, în timp ce la endospermul Haemanthus și fibroblastele de triton durează aproximativ 30 de minute.
metafaza
La sfarsitul prometafazei, cromozomii sunt situati in planul ecuatorial al fusului aproximativ la o distanta egala de ambii poli de diviziune, formand o placa de metafaza. Morfologia plăcii metafazei în celulele animale, de regulă, se distinge printr-o aranjare ordonată a cromozomilor: regiunile centromerice sunt orientate spre centrul fusului, iar umerii sunt îndreptați spre periferia celulei. În celulele vegetale, cromozomii se află adesea în planul ecuatorial al fusului fără o ordine strictă.
Metafaza ocupă o parte semnificativă a perioadei de mitoză și se caracterizează printr-o stare relativ stabilă. În tot acest timp, cromozomii sunt menținuți în planul ecuatorial al fusului datorită forțelor de tensiune echilibrate ale microtubulilor kinetocor, făcând mișcări oscilatorii cu o amplitudine mică în planul plăcii metafazate.
În metafază, precum și în timpul altor faze de mitoză, reînnoirea activă a microtubulilor fusului continuă prin asamblarea intensivă și depolimerizarea moleculelor de tubulină. În ciuda unei anumite stabilizări a fasciculelor de microtubuli kinetocori, există o sortare constantă a microtubulilor interpolari, al căror număr în metafază atinge un maxim.
Până la sfârșitul metafazei, se observă o separare clară a cromatidelor surori, legătura dintre care se păstrează doar în regiunile centromerice. Brațele cromatidelor sunt dispuse paralel între ele, iar golul care le separă devine clar vizibil.
Anafaza
Anafaza este cea mai scurtă etapă a mitozei, care începe cu separarea bruscă și separarea ulterioară a cromatidelor surori către polii opuși ai celulei. Cromatidele se separă cu o viteză uniformă de până la 0,5-2 µm/min și adesea capătă o formă de V. Mișcarea lor se datorează acțiunii unor forțe semnificative, estimate la 10 dine pe cromozom, care este de 10.000 de ori mai mare decât forța necesară pentru a deplasa pur și simplu cromozomul prin citoplasmă cu viteza observată.
De regulă, segregarea cromozomilor în anafază constă din două procese relativ independente numite anafaza A și anafaza B.
Anafaza A este caracterizată prin separarea cromatidelor surori de polii opuși ai diviziunii celulare. În acest caz, aceleași forțe care țineau anterior cromozomii în planul plăcii metafazate sunt responsabile de mișcarea lor. Procesul de separare a cromatidelor este însoțit de o scurtare a lungimii microtubulilor kinetocori depolimerizați. Mai mult, degradarea lor se observă mai ales în regiunea kinetocorilor, din partea capetelor plus. Probabil, depolimerizarea microtubulilor la cinetocori sau în regiunea polilor de diviziune este conditie necesara pentru a muta cromatidele surori, deoarece mișcarea lor se oprește atunci când se adaugă taxol sau apă grea, care au un efect stabilizator asupra microtubulilor. Mecanismul care stă la baza segregării cromozomilor în anafaza A este încă necunoscut.
În timpul anafazei B, polii diviziunii celulare înșiși diverg și, spre deosebire de anafaza A, acest proces are loc datorită asamblarii microtubulilor poli de la capete plus. Firele antiparalele polimerizante ale arborelui, atunci când interacționează, creează parțial forța care împinge polii în afară. Mărimea mișcării relative a polilor în acest caz, precum și gradul de suprapunere a microtubulilor polilor din zona ecuatorială a celulei, variază foarte mult la indivizii diferitelor specii. Pe lângă forțele de respingere, polii de diviziune sunt afectați de forțele de tragere din microtubulii astrali, care sunt create ca urmare a interacțiunii cu proteinele asemănătoare dineinei de pe membrana plasmatică a celulei.
Secvența, durata și contribuția relativă a fiecăruia dintre cele două procese care alcătuiesc anafaza pot fi extrem de diferite. Astfel, în celulele de mamifere, anafaza B începe imediat după începutul divergenței cromatidelor către poli opuși și continuă până la alungirea fusului mitotic de 1,5–2 ori față de cel metafazic. În unele alte celule, anafaza B începe numai după ce cromatidele au ajuns la polii de diviziune. La unele protozoare, în timpul anafazei B, fusul se prelungește de 15 ori în comparație cu metafaza. Anafaza B este absentă în celulele plantelor.
Telofază
Telofaza este considerată stadiul final al mitozei; începutul său este luat ca momentul în care cromatidele surori separate se opresc la polii opuși ai diviziunii celulare. În telofaza timpurie se observă decondensarea cromozomilor și, în consecință, creșterea lor în volum. În apropierea cromozomilor individuali grupați, începe fuziunea veziculelor membranare, care dă naștere reconstrucției membranei nucleare. Materialul pentru construcția membranelor nucleelor fiice nou formate sunt fragmente din membrana nucleară degradată inițial a celulei mamă, precum și elemente ale reticulului endoplasmatic. În acest caz, veziculele individuale se leagă de suprafața cromozomilor și se îmbină împreună. Membranele nucleare exterioare și interioare sunt restaurate treptat, lamina nucleară și porii nucleari sunt restaurați. În procesul de reparare a anvelopei nucleare, veziculele membranare discrete se conectează probabil la suprafața cromozomilor fără a recunoaște secvențe de nucleotide specifice, deoarece experimentele au arătat că repararea membranei nucleare are loc în jurul moleculelor de ADN împrumutate de la orice organism, chiar și de la un virus bacterian. În interiorul nucleilor celulari nou formați, cromatina trece într-o stare dispersată, sinteza ARN-ului se reia și nucleolii devin vizibili.
În paralel cu procesele de formare a nucleelor celulelor fiice în telofază, începe și se termină dezasamblarea microtubulilor fusului de fisiune. Depolimerizarea are loc în direcția de la polii de diviziune la planul ecuatorial al celulei, de la capetele minus la capetele plus. În același timp, microtubulii sunt stocați cel mai lung în partea de mijloc a fusului, care formează corpul Fleming rezidual.
Sfârșitul telofazei coincide în principal cu diviziunea corpului celulei mamă - citokineza. În acest caz, se formează două sau mai multe celule fiice. Procesele care conduc la divizarea citoplasmei încep încă de la mijlocul anafazei și pot continua după sfârșitul telofazei. Mitoza nu este întotdeauna însoțită de diviziunea citoplasmei, astfel încât citokineza nu este clasificată ca o fază separată a diviziunii mitotice și este de obicei considerată ca parte a telofazei.
Există două tipuri principale de citokineză: diviziunea prin constricția transversală a celulei și diviziunea prin formarea unei plăci celulare. Planul de diviziune celulară este determinat de poziția fusului mitotic și merge în unghi drept față de axa lungă a fusului.
La împărțirea printr-o constricție transversală a celulei, locul de diviziune a citoplasmei este stabilit în prealabil în timpul perioadei de anafază, când apare un inel contractil de filamente de actină și miozină în planul plăcii metafazei de sub membrana celulară. Ulterior, datorită activității inelului contractil, se formează o brazdă de fisiune, care se adâncește treptat până când celula este complet divizată. La sfârșitul citokinezei, inelul contractil se dezintegrează complet, iar membrana plasmatică se contractă în jurul corpului Fleming rezidual, care constă dintr-o acumulare de rămășițe din două grupuri de microtubuli poli strâns împachetate împreună cu material dens de matrice.
Diviziunea prin formarea unei plăci celulare începe cu deplasarea micilor vezicule limitate de membrană către planul ecuatorial al celulei. Aici ele fuzionează pentru a forma o structură în formă de disc, închisă în membrană, placa celulară timpurie. Veziculele mici provin în principal din aparatul Golgi și călătoresc spre planul ecuatorial de-a lungul microtubulilor polului rezidual ai fusului, formând o structură cilindrică numită fragmoplast. Pe măsură ce placa celulară se extinde, microtubulii fragmoplastului timpuriu se deplasează simultan la periferia celulei, unde, datorită noilor vezicule membranare, creșterea plăcii celulare continuă până la fuziunea sa finală cu membrana celulei mamă. După separarea finală a celulelor fiice, în placa celulară se depun microfibrile de celuloză, completând formarea unui perete celular rigid.
Pentru a determina finalizarea fiecărei faze a ciclului celular, este necesar să existe puncte de control în ea. Dacă celula „trece” punctul de control, atunci aceasta continuă să „se miște” prin ciclul celular. Dacă unele circumstanțe, cum ar fi deteriorarea ADN-ului, împiedică celula să treacă printr-un punct de control, care poate fi comparat cu un fel de punct de control, atunci celula se oprește și nu are loc o altă fază a ciclului celular, cel puțin până când obstacolele sunt îndepărtate. , împiedicând cușca să treacă prin punctul de control.
Diviziunea celulară mitotică
Mitoză(din greaca. Mitos - fir), numita si cariokineza, sau diviziunea celulara indirecta, este un mecanism universal de diviziune celulara. Mitoza urmează perioada G2 și completează ciclul celular.
Durează 1-3 ore și asigură distribuția uniformă a materialului genetic în celulele fiice. Mitoza are 4 faze principale: profaza, metafaza, anafaza si telofaza.
Mitoza este unul dintre procesele fundamentale ale ontogenezei. Diviziunea mitotică asigură creșterea eucariotelor multicelulare prin creșterea populațiilor de celule tisulare.
Ca urmare a diviziunii mitotice a celulelor meristeme, numărul celulelor din țesutul vegetal crește. Fragmentarea unui ovul fertilizat și creșterea majorității țesuturilor la animale au loc și prin diviziuni mitotice.
Pe baza caracteristicilor morfologice, mitoza este subdivizată condiționat în etape: profază, prometafază, metafază, anafază, telofază. Primele descrieri ale fazelor mitozei și stabilirea secvenței lor au fost întreprinse în anii 70-80 ai secolului XIX. La sfârșitul anilor 1870, histologul german Walter Flemming a inventat termenul de „mitoză” pentru a se referi la procesul de diviziune celulară indirectă.
Durata medie a mitozei este de 1-2 ore. Mitoza celulelor animale, de regulă, durează 30-60 de minute, iar plantele - 2-3 ore. Timp de 70 de ani, în corpul uman sunt efectuate în total aproximativ 10 14 diviziuni celulare.
Primele descrieri incomplete cu privire la comportamentul și modificările nucleelor în celulele în diviziune se găsesc în lucrările oamenilor de știință la începutul anilor 1870.
În lucrarea botanistului rus Russov, datată 1872, plăcile metafază și anafază, constând din cromozomi individuali, sunt descrise și descrise clar.
Un an mai târziu, zoologul german G.A. Schneider chiar mai clar și mai consecvent, dar, desigur, nu a descris pe deplin diviziunea mitotică folosind exemplul zdrobirii ouălor din turbellaria rectală Mesostomum. În opera sa, în esență, descrisă și ilustrată în succesiunea corectă principalele faze ale mitozei: profaza, metafaza, anafaza (devreme și târzie). În 1874, botanistul din Moscova I.D. Chistyakov a observat, de asemenea, faze separate ale diviziunii celulare în sporii de mușchi și coada-calului. În ciuda primelor succese, nici Russov, nici Schneider, nici Chistyakov nu au fost capabili să ofere o descriere clară și consecventă a diviziunii mitotice.
În 1875, au fost publicate lucrări care conțineau descrieri mai detaliate ale mitozelor. O. Byuchli a oferit o descriere a modelelor citologice în zdrobirea ouălor de viermi rotunzi și moluște și în celulele spermatogene ale insectelor.
E. Strasburger a studiat diviziunea mitotică în celulele algei verzi spirogyra, în celulele mamă ale polenului de ceapă și în celulele sporilor mamă ale mușchiului club. Referindu-se la lucrările lui O. Buechli și pe baza propriilor cercetări, E. Strasburger a atras atenția asupra unității proceselor de diviziune celulară în celulele vegetale și animale.
Până la sfârșitul anului 1878 - începutul anului 1879 au apărut lucrări detaliate Schleicher și W. Flemming. În lucrarea sa din 1879, Schleicher a propus termenul de „cariocineză” pentru a se referi la procesele complexe de diviziune celulară, implicând mișcarea părțile constitutive miezuri. Walter Flemming a fost primul care a introdus termenul „mitoză” pentru a se referi la diviziunea celulară indirectă, care mai târziu a devenit general acceptată. Flemming deține și formula finală a definiției mitozei ca proces ciclic, culminând cu diviziunea cromozomilor între celulele fiice.
În 1880 O.V. Baranetsky a stabilit structura elicoidală a cromozomilor. În cursul cercetărilor ulterioare, au fost dezvoltate idei despre spiralizarea și despiralizarea cromozomilor în timpul ciclului mitotic.
La începutul anilor 1900, cromozomii au fost identificați ca purtători informații ereditare, care a explicat în continuare rol biologic mitoza, care constă în formarea de celule fiice identice genetic.
În anii 1970, a început decodificarea și studiul detaliat al regulatorilor diviziunii mitotice, datorită unei serii de experimente privind fuziunea celulelor în diferite etape ale ciclului celular. În acele experimente, când o celulă în faza M a fost combinată cu o celulă în oricare dintre etapele de interfază (G1, S sau G2), celulele de interfază au trecut în starea mitotică (a început condensarea cromozomilor și învelișul nuclear s-a dezintegrat) .
Ca urmare, s-a ajuns la concluzia că există un factor (sau factori) în celula citoplasmatică care stimulează mitoza, sau, cu alte cuvinte, factorul M-stimulator (MSF, din engleza M-phase-promoting factor, MPF) .
Pentru prima dată, „factorul de stimulare a mitozei” a fost descoperit în ouăle mature nefertilizate ale broaștei cu gheare, care se află în faza M a ciclului celular. Citoplasma unui astfel de ou, injectată în ovocit, a condus la o tranziție prematură la faza M și la debutul maturizării ovocitului (inițial, abrevierea MPF înseamnă Maturation Promoting Factor, care se traduce prin „maturare”. factor de promovare"). În cursul experimentelor ulterioare, a fost stabilită o valoare universală și, în același timp grad înalt conservatorismul „factorului de stimulare a mitozei”: extractele preparate din celule mitotice ale unei mari varietăți de organisme, atunci când au fost introduse în ovocitele de broaște cu gheare, le-au transferat în faza M.
Studiile ulterioare au arătat că factorul care stimulează mitoza este un complex heterodimeric format dintr-o proteină ciclină și o protein kinază dependentă de ciclină. Ciclina este proteina reglatoareși se găsește la toate eucariotele. Concentrația sa crește periodic în timpul ciclului celular, atingând un maxim în metafaza mitozei. Odată cu apariția anafazei, se observă o scădere bruscă a concentrației de ciclină, datorită clivajului acesteia cu ajutorul complexelor proteolitice proteice complexe - proteozomi. Protein kinaza dependentă de ciclină este o enzimă (fosforilază) care modifică proteinele prin transferul unei grupări fosfat din ATP la aminoacizii serină și treonină. Astfel, odată cu stabilirea rolului și structurii principalului regulator al diviziunii mitotice, au început studiile asupra mecanismelor subtile de reglare a mitozei, care continuă până în prezent.
Dezvoltarea unei tipologii și clasificări unificate a mitozelor este complicată de o întreagă gamă de caracteristici care, în diverse combinații, creează o varietate și eterogenitate de modele de diviziune mitotică. În același timp, opțiunile de clasificare separate dezvoltate pentru unii taxoni sunt inacceptabile pentru alții, deoarece nu țin cont de specificul mitozelor lor. De exemplu, unele variante ale clasificării mitozelor, caracteristice organismelor animale sau vegetale, se dovedesc a fi inacceptabile pentru alge.
Una dintre caracteristicile cheie care stau la baza diferitelor tipologii și clasificări ale diviziunii mitotice este comportamentul învelișului nuclear. Dacă formarea fusului și diviziunea mitotică în sine se desfășoară în interiorul nucleului fără a distruge membrana nucleară, atunci acest tip de mitoză se numește închis. Mitoza cu prăbușirea învelișului nuclear, respectiv, se numește deschisă, iar mitoza cu prăbușirea membranei numai la polii fusului, cu formarea de „ferestre polare” - semiînchise.
O altă trăsătură caracteristică este tipul de simetrie a fusului mitotic. În pleuromitoză, fusul de diviziune este bilateral simetric sau asimetric și constă, de obicei, din două semifusuri situate în metafază-anafaza la un unghi unul față de celălalt. Categoria ortomitozelor se caracterizează prin simetria bipolară a fusului de fisiune, iar în metafază există adesea o placă ecuatorială distinsă.
În cadrul semnelor indicate, cea mai numeroasă este o ortomitoză tipică deschisă, pe exemplul căreia principiile și etapele diviziunii mitotice sunt discutate mai jos. Acest tip de mitoză este caracteristic animalelor, plantelor superioare și unor protozoare.
Profaza începe cu condensarea cromozomilor, care devin vizibili la microscop cu lumină sub formă de structuri filamentoase. Fiecare cromozom este format din două cromatide surori paralele conectate la centromer. Nucleolul și învelișul nuclear dispar până la sfârșitul fazei (aceasta din urmă se descompune în vezicule membranare similare elementelor EPS, iar complexul de pori și lamina se disociază în subunități). Carioplasma se amestecă cu citoplasma.
Centriolii migrează către polii opuși ai celulei și dau naștere filamentelor fusului mitotic (acromatin). În regiunea centromerului se formează complexe proteice speciale - kinetocori, de care sunt atașați niște microtubuli fusi (microtubuli kinetocori); s-a demonstrat că cinetocorii înșiși sunt capabili să inducă asamblarea microtubulilor și, prin urmare, pot servi ca centre de organizare a microtubulilor. Restul microtubulilor fusului se numesc microtubuli polari, deoarece se extind de la un pol al celulei la celălalt; microtubulii aflați în afara fusului, divergenți radial de la centrii celulari către plasmalema, au primit denumirea de astral sau microtubuli (fire) de strălucire.
Metafaza corespunde nivelului maxim de condensare al cromozomilor, care se aliniază în regiunea ecuatorială a fusului mitotic, formând o imagine a plăcii ecuatoriale (metafază) (vedere laterală) sau a stelei părinte (vedere de la poli). Cromozomii se deplasează în planul ecuatorial și sunt ținuți în acesta datorită tensiunii echilibrate a microtubulilor kinetocor. Până la sfârșitul acestei faze, cromatidele surori sunt separate printr-un gol, dar sunt reținute în regiunea centromerului.
Anafaza începe cu divizarea sincronă a tuturor cromozomilor în cromatide surori (în regiunea centromerului) și mișcarea cromozomilor fiice către polii opuși ai celulei, care are loc de-a lungul microtubulilor fusului cu o viteză de 0,2-0,5 µm/min. Semnalul pentru debutul anafazei include o creștere bruscă (de un ordin de mărime) a concentrației de cationi de calciu în hialoplasmă, secretate de veziculele membranare care formează grupuri la polii fusului. Mecanismul mișcării cromozomilor în anafază nu a fost pe deplin elucidat, cu toate acestea, s-a stabilit că, pe lângă actină, proteine precum miozina și dineina, precum și o serie de proteine reglatoare, sunt prezente în regiunea fusului. Conform unor observații, se datorează scurtării (dezasamblarii) microtubulilor atașați cinetocorilor. Anafaza se caracterizează prin alungirea fusului mitotic datorită unei anumite divergențe a polilor celulari. Se termină cu acumularea a două seturi identice de cromozomi la polii celulei, care formează imagini ale stelelor (stadiul stelelor fiice). La sfarsitul anafazei, datorita contractiei microfilamentelor de actina, concentrandu-se in jurul circumferintei celulei (inel contractil), incepe sa se formeze o constrictie celulara care, adancindu-se, va duce la citotomie in faza urmatoare.
Telofaza este etapa finală a mitozei, în timpul căreia nucleii celulelor fiice sunt reconstruiți și diviziunea lor este finalizată. În jurul cromozomilor condensați ai celulelor fiice din veziculele membranare (conform altor surse, din EPS), se restabilește caryolema, cu care este asociată lamina emergentă, reapar nucleolii, care se formează din secțiuni ale cromozomilor corespunzători. Nucleii celulari cresc treptat, iar cromozomii se despiralizeaza si dispar progresiv, fiind inlocuiti de modelul cromatinic al nucleului de interfaza. În același timp, constricția celulară se adâncește, iar celulele rămân conectate pentru o perioadă de timp printr-o punte citoplasmatică îngustată care conține un mănunchi de microtubuli (corpul mijlociu). Ligarea ulterioară a citoplasmei se termină cu formarea a două celule fiice. În telofază are loc distribuția organelelor între celulele fiice; Uniformitatea acestui proces este facilitată de faptul că unele organele sunt destul de numeroase (de exemplu, mitocondriile), în timp ce altele (cum ar fi EPS și complexul Golgi) se descompun în mici fragmente și vezicule în timpul mitozei.
Mitozele atipice apar atunci când aparatul mitotic este deteriorat și se caracterizează printr-o distribuție neuniformă a materialului genetic între celule - aneuploidie (din greacă an - nu, eu - corect, ploon - add); în multe cazuri, citotomia este absentă, ducând la formarea de celule gigantice. Mitozele atipice sunt caracteristice tumorilor maligne și țesuturilor iradiate. Cu cât frecvența lor este mai mare și cu cât este mai mare gradul de aneuploidie, cu atât tumora este mai malignă. Încălcarea diviziunii celulare mitotice normale poate fi cauzată de anomalii cromozomiale, care sunt numite aberații cromozomiale (din latină Aberratio - deviație). Variantele aberațiilor cromozomiale sunt aderența cromozomilor, ruperea lor în fragmente, pierderea unui loc, schimbul de fragmente, dublarea secțiunilor individuale de cromozomi etc. Aberațiile cromozomiale pot apărea spontan, dar mai des se dezvoltă datorită acţiunea mutagenilor şi a radiaţiilor ionizante asupra celulelor.
Cariotiparea - un studiu de diagnostic pentru evaluarea cariotipului (setul de cromozomi) se realizează prin examinarea cromozomilor din placa metafază. Pentru cariotipare se obtine o cultura celulara in care se introduce colchicina, o substanta care blocheaza formarea fusului mitotic. Din astfel de celule sunt extrași cromozomii, care sunt apoi colorați și identificați. Cariotipul uman normal este reprezentat de 46 de cromozomi - 22 de perechi de autozomi și doi cromozomi sexuali (XY la bărbați și XX la femei). Cariotiparea poate diagnostica o serie de boli asociate cu anomalii cromozomiale, în special, sindromul Down (trisomia celui de-al 21-lea cromozom), Edwards (trisomia celui de-al 18-lea cromozom), Patau (trisomia celui de-al 13-lea cromozom), precum și o serie de sindroame asociate cu anomalii ale cromozomilor sexuali - sindromul Klinefelter ( genotip - XXY), Turner (genotip - XO) și altele.
Se presupune că procesul mitotic complex al organismelor superioare s-a dezvoltat treptat din mecanismele diviziunii procariote. Această presupunere este susținută de faptul că procariotele au apărut cu aproximativ un miliard de ani mai devreme decât primele eucariote. În plus, proteine similare sunt implicate în mitoza eucariotă și fisiunea binară procariotă.
Posibilele etape intermediare între fisiunea binară și mitoză pot fi urmărite la eucariotele unicelulare, în care membrana nucleară nu este distrusă în timpul diviziunii. La majoritatea celorlalte eucariote, inclusiv plante și animale, fusul de fisiune se formează în afara nucleului, iar învelișul nuclear este distrus în timpul mitozei. Deși mitoza la eucariotele unicelulare nu este încă bine înțeleasă, se poate presupune că a provenit din fisiunea binară și a atins în cele din urmă nivelul de complexitate care există în organismele multicelulare.
La multe eucariote protozoare, mitoza a rămas și un proces asociat membranei, dar acum nu mai este plasmă, ci nucleară.
Principalele mecanisme de reglare ale mitozei sunt procesele de fosforilare și proteoliză.
Reacțiile reversibile de fosforilare și defosforilare permit evenimente mitotice reversibile, cum ar fi asamblarea/dezintegrarea fusului sau dezintegrarea/repararea anvelopei nucleare. Proteoliza stă la baza evenimentelor ireversibile ale mitozei, cum ar fi separarea cromatidelor surori în anafaza sau distrugerea ciclinelor mitotice în etapele ulterioare ale mitozei.
Diviziunea tuturor celulelor eucariote este asociată cu formarea unui aparat special pentru diviziunea celulară.
Un rol activ în diviziunea celulară mitotică este adesea atribuit structurilor citoscheletice. Fusul mitotic bipolar, care constă din microtubuli și proteine asociate, este universal atât pentru celulele animale, cât și pentru cele vegetale. Fusul de diviziune asigură o distribuție strict identică a cromozomilor între polii de diviziune, în regiunea cărora se formează nucleii celulelor fiice în telofază.
Procesul de mitoză asigură o distribuție strict uniformă a cromozomilor între doi nuclei fiice, astfel încât într-un organism multicelular toate celulele au exact aceleași (ca număr și caracter) seturi de cromozomi.
Cromozomii conțin informații genetice codificate în ADN și, prin urmare, un proces mitotic regulat, ordonat asigură și transferul complet al tuturor informațiilor către fiecare dintre nucleele fiice; ca urmare, fiecare celulă are toată informația genetică necesară dezvoltării tuturor caracteristicilor organismului. În această privință, devine clar de ce o celulă luată dintr-o plantă adultă complet diferențiată se poate dezvolta, în condiții adecvate, într-o plantă întreagă. Am descris mitoza într-o celulă diploidă, dar acest proces se desfășoară într-un mod similar în celulele haploide, de exemplu, în celulele din generația gametofiților de plante.
Catelul creste si creste in dimensiune datorita diviziunii celulelor somatice, numite mitoză. Mitoza este o diviziune indirectă a unei celule somatice atunci când apar modificări complexe în nucleul și citoplasma acesteia. După fecundarea (ovogamia) ovulului de către spermatozoid (fuziunea sau copularea gameților), un zigot(oocist) - un organism nou, format dintr-o singură celulă. Procesul de creștere și dezvoltare a unui nou organism începe din momentul primei diviziuni mitotice a acestei celule (materne), când două celule fiice (mai precis, surori), complet asemănătoare cu aceasta, iau naștere din ea și continuă până la moarte. .
Fig.1 Structura celulară
În timpul mitozei, apar următoarele:
1- dublarea substanței cromozomilor;
2- modificarea stării fizice și a organizării chimice a cromozomilor;
3- divergența cromozomilor surori la polii celulei;
4- diviziunea ulterioară a citoplasmei și refacerea completă a doi nuclei în celule noi.
În mitoză, ciclul de viață al genelor nucleare este stabilit: duplicare, distribuție și funcționare. Se numește perioada dintre diviziunile celulare interfaza, timp în care în ea au loc procese de viață activă și pregătirea pentru următoarea diviziune. Întregul ciclu de schimbări care au loc în celulă
de la o diviziune la alta se numeste ciclu mitotic. Acesta din urmă constă din două perioade principale - interfaza și mitoza în sine.
Ca urmare a mitozei, dintr-o celulă se formează două celule cu cromozomi identici. Astfel, mitoza asigură continuitatea și constanța numărului și mulțimii, adică specificitatea calitativă a cromozomilor în generații succesive de celule în diviziune (vezi Fig. 2).
În interfaza, perioada dintre două diviziuni celulare succesive are loc replicarea (autoduplicarea sau autodublarea) ADN-ului în nucleu și, prin urmare, numărul de cromozomi din celulă (formarea cromatidelor surori ținute împreună de centromer, adică , corpul care îndeplinește funcția centrului mecanic al cromozomului), precum și despiralizarea acestuia din urmă.
În metafază, sau faza centrală a diviziunii nucleare, un cromozom format din două cromatide se transformă în doi cromozomi fiice.
Orez. 2. Mitoza
 1 - trei perechi de cromozomi; 2 - replicarea cromozomilor cu formarea de cromatide surori conectate în regiunea centromerului; 3 - la polii membranei nucleare, centrozomii sunt vizibili cu raze asteriale mergând în regiunea centromerică a cromatidelor, aliniate de-a lungul ecuatorului pentru a separa centromerii cromatidelor surori și a-i separa la poli diferiți; 4 - despiralizarea cromozomilor, refacerea membranei nucleare și formarea unui sept celular cu formarea a două celule fiice identice cu mama cu exact același număr de cromozomi ca și în ea
1 - trei perechi de cromozomi; 2 - replicarea cromozomilor cu formarea de cromatide surori conectate în regiunea centromerului; 3 - la polii membranei nucleare, centrozomii sunt vizibili cu raze asteriale mergând în regiunea centromerică a cromatidelor, aliniate de-a lungul ecuatorului pentru a separa centromerii cromatidelor surori și a-i separa la poli diferiți; 4 - despiralizarea cromozomilor, refacerea membranei nucleare și formarea unui sept celular cu formarea a două celule fiice identice cu mama cu exact același număr de cromozomi ca și în ea
În anafază, are loc divizarea și divergența cromozomilor fiice la polii celulei, adică restabilirea numărului lor adecvat. În telofază, etapa finală a diviziunii celulare, cromozomii iau aceeași formă ca înainte de începerea diviziunii, iar cantitatea de ADN din fiecare nucleu fiică este înjumătățită față de etapele anterioare. Astfel, ambele celule fiice conțin aceeași cantitate de citoplasmă și seturi identice de cromozomi și sunt gata să sufere mitoză.
Nu toate celulele somatice ale corpului se divid în mod constant. În procesul de dezvoltare embrionară, are loc diferențierea organelor și țesuturilor, dezvoltându-se pe propria lor cale specifică, predeterminată genetic. Prin urmare, unele celule se transformă în celule cerebrale, altele în celule sanguine etc. Mai mult, unele dintre ele se divid în mod constant, în timp ce altele doar la un anumit stadiu de dezvoltare sau, dacă este necesar, sunt responsabile, de exemplu, de regenerare.
procese (restaurative).