Permeabilitatea și conductivitatea membranei celulare. Porii lipidici: stabilitatea și permeabilitatea membranei
Citeste si
3.3. Potențial de repaus al membranei
3.3.1. Caracteristici generale și cauza imediată de formare
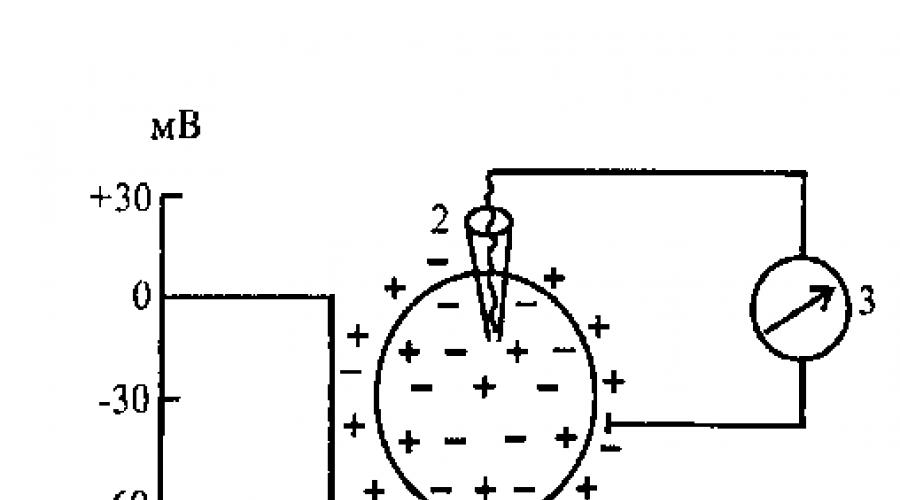
Potențial de repaus (1111) - diferența de potențiale electrice dintre părțile exterioare și interioare ale membranei celulare. A lui
valoarea în celulele nervoase este de 60-80 mV. La înregistrarea PP, fasciculul osciloscopului în timpul puncției membranei celulare cu un microelectrod deviază brusc și prezintă o sarcină negativă în interiorul celulei (Fig. 3.1).
PP joacă un rol extrem de important în viața neuronului în sine și a organismului în ansamblu.În special, formează baza pentru ra informația din celula nervoasă, asigură reglarea activității organelor interne și a sistemului musculo-scheletic prin declanșarea proceselor de excitare și contracție în mușchi. Conform teoriei membrana-ionice (Bernstein, Hodgkin, Huxley, Katz), cauza directă a formării PP este concentrația inegală de anioni și cationi în interiorul și în afara celulei (Tabelul 3.1).

În neuroni, concentrația de ioni K + în interiorul celulei este de 20-40 de ori mai mare decât în afara celulei; concentrația ionilor de Na + în afara celulei este de 10-12 ori mai mare decât în celulă. Există de 10-20 de ori mai mulți ioni SG în afara celulei decât în interiorul celulei. Există o cantitate mică de ioni de Mg 2+ în celulă. Ionul de Ca 2+ din celulă este legat în principal de ATP, citrat și glutamat. Rezervorul pentru ionii de Ca 2+ este reticulul endoplasmatic. În stare liberă, calciul se găsește mai ales în afara celulei; în hialoplasmă
foarte putin. Acest lucru se datorează parțial transportului activ al ionilor de Ca 2+ în exterior prin membrana celulară, parțial datorită absorbției acestuia de către reticulul endoplasmatic și alte organite, cum ar fi mitocondriile. În celulă se găsesc și anioni cu molecule mari, în principal molecule de proteine încărcate negativ, cum ar fi glutamatul, aspartatul și, de asemenea, fosfații organici. Diferiți ioni sunt distribuiți neuniform pe ambele părți ale membranei celulare, în primul rând, datorită permeabilității inegale a membranei celulare pentru diferiți ioni și, în al doilea rând, ca rezultat al funcționării pompelor ionice care transportă ioni în și din celulă împotriva concentrație și gradienți electrici. În ceea ce privește definiția „permeabilității” și „conductivității”, nu există încă un consens.
3.3.2. Rolul permeabilității membranei celulare și sarcinile sale de suprafață
DAR. Terminologie.În prezent, diferiți autori interpretează diferit termenii „permeabilitate” și „conductivitate”. Sub permeabilitatea membranei celulare, ne referim la capacitatea acesteia de a trece apa și particulele - încărcate (ioni) și neîncărcate conform legilor difuziei și filtrării. Permeabilitatea membranei celulare este determinată de următorii factori: 1) prezența diferitelor canale ionice în membrană - controlate (cu un mecanism de poartă) și necontrolate (canale de scurgere); 2) dimensiunile canalelor și dimensiunile particulelor; 3) solubilitatea particulelor în membrană (membrana celulară este permeabilă la lipidele solubile în ea și impermeabilă la peptide).
Termenul „conductivitate” ar trebui folosit numai în legătură cu particulele încărcate. Prin urmare, prin conductivitate înțelegem capacitatea particulelor încărcate (ioni) de a trece prin membrana celulară în funcție de gradientul electrochimic (o combinație de gradienți electrici și de concentrație).
După cum se știe, ionii, ca și particulele neîncărcate, trec printr-o membrană dintr-o regiune de concentrație mare într-o regiune de concentrație scăzută. Cu un gradient de concentrație mare și o bună permeabilitate a membranei care separă soluțiile corespunzătoare, conductivitatea ionică poate fi ridicată și se observă curentul ionic unidirecțional. Când concentrația de ioni de pe ambele părți ale membranei devine egală, conductivitatea ionică va scădea, curentul ionic unidirecțional se va opri, deși permeabilitatea va rămâne aceeași - ridicată. În plus, conductivitatea unui ion la o permeabilitate constantă a membranei depinde și de sarcina ionului: sarcinile asemănătoare se resping reciproc, spre deosebire de sarcinile se atrag, adică. Un rol important în conductivitatea unui ion este jucat de sarcina electrică a acestuia. Este posibilă o situație când, cu o bună permeabilitate a membranei, conductivitatea ionilor prin membrană se dovedește a fi scăzută sau nulă, în absența unei forțe motrice (concentrație și/sau gradienți electrici).
Astfel, conductivitatea unui ion depinde de gradientul său electrochimic și de permeabilitatea membranei: cu cât sunt mai mari, cu atât conductivitatea ionului prin membrană este mai bună. Mișcarea ionilor în și în afara celulei în funcție de concentrație și gradienți electrici celulă în repaus realizată în primul rând prin negestionate(fără mecanism de poartă) canale (canale de scurgere). Canalele necontrolate sunt întotdeauna deschise, practic nu își schimbă capacitatea în timpul acțiunii electrice asupra membranei celulare și a excitării acesteia. Canalele neadministrate sunt împărțite în ion-selectiv canale (de exemplu, canale lente de potasiu necontrolate) și ion-neselectiv canale. Acesta din urmă lasă să treacă diverși ioni: K + , Na + , C1 _ .
B. Rolul permeabilității membranei celulare și al diferiților ioni în formarea 1111(Fig. 3.2.). Vasul este separat de o membrană semi-permeabilă. Ambele jumătăți sunt umplute cu mortar. K2 ASA DE4 diferite concentrații (O și C2), cu Ci< С2. Мембрана проницаема для иона К + и непроницаема для S04 2 ". Ионы К + перемещаются согласно

Ionii de Na + și K + dintr-o celulă vie în repaus se deplasează și ei prin membrană conform legilor difuziei, în timp ce K + părăsește celula într-o cantitate mult mai mare decât Na + intră în celulă, deoarece permeabilitatea membranei celulare pt. K+ este de aproximativ 25 de ori mai permeabil la Na+.
anioni organici datorită dimensiunilor lor mari, nu pot părăsi celula, prin urmare, în interiorul celulei în stare de repaus, există mai mulți ioni negativi decât
pozitiv. Din acest motiv, celula din interior are o sarcină negativă. Interesant, în toate punctele celulei, sarcina negativă este aproape aceeași. Acest lucru este evidențiat de aceeași valoare RI atunci când microelectrodul este introdus la diferite adâncimi în celulă, așa cum a fost cazul în experimentele lui Hodgkin, Huxley și Katz. Sarcina din interiorul celulei este negativă atât în mod absolut (hialoplasma celulară conține mai mulți anioni decât cationi), cât și în raport cu suprafața exterioară a membranei celulare.
Potasiul este ionul principal responsabil de formare PP. Acest lucru este dovedit de rezultatele experimentului cu perfuzia conținutului intern al axonului de calmar gigant cu soluții saline. Odată cu scăderea concentrației ionilor K + în soluția de perfuzie, PP scade, cu creșterea concentrației lor, PP crește. În starea de repaus a celulei, se stabilește un echilibru dinamic între numărul de ioni K + care părăsesc celula și intră în celulă. Gradienții electrici și de concentrație se contracarează reciproc: în funcție de gradientul de concentrație, K + tinde să părăsească celula, sarcina negativă din interiorul celulei și sarcina pozitivă a suprafeței exterioare a membranei celulare împiedică acest lucru. Când concentrația și gradienții electrici sunt echilibrate, numărul de ioni K + care părăsesc celula este comparat cu numărul de ioni K + care intră în celulă. În acest caz, așa-numitul potenţial de echilibru.
Potențialul de echilibru pentru un ion poate fi calculat folosind formula Nernst. Concentrația unui ion încărcat pozitiv în afara celulei este scrisă la numărător în formula Nernst, iar concentrația unui ion în interiorul celulei este scrisă la numitor. Pentru ionii încărcați negativ, aranjamentul este opus.
E_LG. 1p M,
~~ ZF [ ion\ "
Unde E-tp este potențialul creat de ionul dat; R - constanta de gaz (8,31 Dm); T - temperatura absolută (273+37°С); Z este valența ionului; F - constanta Faraday (9,65 x 10 4); [scop],- - concentrația ionilor în interiorul celulei din interior; [ ion] 0 - concentrația ionilor în mediul extern al celulei în exterior. Potențialul de echilibru al ionului Na + celule nervoase Eieu - +55 mV, ion de potasiu Ek --70 mV.
ContribuţieN / A + iar C1 ~ în crearea PP. Permeabilitatea membranei celulare în repaus pentru ionul Na + este foarte scăzută, mult mai mică decât pentru ionul K + este însă prezenți, prin urmare, ionii Na +, în funcție de concentrație și gradienți electrici, tind și în o cantitate mică trece în celulă. Acest lucru duce la o scădere a PP, deoarece numărul total de ioni încărcați pozitiv de pe suprafața exterioară a membranei celulare scade, deși ușor, iar o parte din ionii negativi din interiorul celulei sunt neutralizați de ionii Na + încărcați pozitiv care intră în celulă. Intrarea ionilorN / A + în interiorul celulei se reduce PP. Efectul SG asupra valorii PP este opus și depinde de permeabilitatea membranei celulare pentru ionii SG. Cert este că ionul SG, în funcție de gradientul de concentrație, tinde și trece în celulă. Previne intrarea ionului SG în celulă printr-un gradient electric, de deoarece sarcina din interiorul celulei este negativă, la fel ca și sarcina lui SG. Pe există un echilibru între forțele gradientului de concentrație, care favorizează intrarea ionului SG în celulă, și gradientul electric, care împiedică intrarea ionului SG în celulă. Prin urmare, concentrația intracelulară a ionilor SG este mult mai mică decât cea extracelulară. Când ionul SG intră în celulă, numărul de sarcini negative din exteriorul celulei scade oarecum, în timp ce în interiorul celulei crește: ionul SG este adăugat la anionii mari, pe bază de proteine, aflați în interiorul celulei. Acești anioni, datorită dimensiunilor lor mari, nu pot trece prin canalele membranei celulare în exteriorul celulei - în interstițiu. În acest fel, ionul SG, pătrunzând în celulă, crește PP. Parțial, precum și în afara celulei, ionii Na + și C1~ din interiorul celulei se neutralizează reciproc. Ca rezultat, aportul comun de ioni Na + și C1~ în celulă nu afectează semnificativ valoarea PP.
C. Un anumit rol în formarea PP îl au sarcinile de suprafață ale membranei celulare în sine și ionii de Ca. 2+ . Externe și interne suprafețele membranei celulare poartă propriile sarcini electrice, predominant negativ. Acestea sunt molecule polare ale membranei celulare: glicolipide, fosfolipide, glicoproteine. Sarcinile negative externe fixe, neutralizând sarcinile pozitive ale suprafeței exterioare a membranei, reduc RI. Sarcinile negative interne fixe ale membranei celulare, dimpotrivă, adunându-se cu anioni în interiorul celulei, cresc PP.
Rolul ionilor de Ca 2+ în formarea PP constă în faptul că acestea interacționează cu sarcinile fixe negative externe ale membranei celulare și le neutralizează, ceea ce duce la creșterea și stabilizarea PP.
Astfel, RI nu este doar suma algebrică a tuturor sarcinilor ionice din exteriorul și din interiorul celulei, ci și suma algebrică a sarcinilor negative de suprafață externe și interne ale membranei în sine.
În timpul măsurătorilor, se presupune că potențialul mediului din jurul celulei este zero.În raport cu potențialul zero al mediului extern, potențialul mediului intern al neuronului, după cum s-a menționat, este de aproximativ -60-80 mV. Deteriorarea celulelor duce la creșterea permeabilității membranele celulare, în urma căreia diferența de permeabilitate pentru ionii K + și Na + scade, în timp ce PP scade. Modificări similare apar în timpul ischemiei tisulare. În celulele grav deteriorate, PP poate scădea la nivelul echilibrului Donann, când concentrația în interiorul și în exteriorul celulei va fi determinată doar de permeabilitatea selectivă a membranei celulare în starea de repaus a celulei, ceea ce poate duce la o perturbare a celulei. activitatea electrică a neuronilor. Cu toate acestea, chiar și în mod normal, ionii se mișcă în funcție de gradientul electrochimic, dar PP nu este perturbat.
| " |
PERMEABILITATE- capacitatea celulelor și țesuturilor de a absorbi, elibera și transporta substanțe chimice, trecându-le prin membranele celulare, pereții vasculari și celulele epiteliale. Celulele și țesuturile vii sunt într-o stare de schimb chimic continuu. substanțe cu mediul înconjurător. Principala barieră (vezi Funcțiile de barieră) în mișcarea substanțelor este membrana celulară. Prin urmare, din punct de vedere istoric, mecanismele lui P. au fost studiate în paralel cu studiul structurii și funcției membranelor biologice (vezi Membrane biologice).
Există P. pasiv, transport activ de substanțe și cazuri speciale de P. asociate cu fagocitoză (vezi) și pinocitoză (vezi).
În conformitate cu teoria membranei a lui P., P. pasiv se bazează pe diferite tipuri de difuzie a unei substanțe prin membranele celulare (vezi Difuzia
unde dm este cantitatea de substanță care se difuzează în timpul dt prin zona S; dc/dx - gradient de concentrație a substanței; D este coeficientul de difuzie.
Orez. Fig. 1. Organizarea moleculară a unui antibiotic ionofor (valinomicina): a - formula structurală a unei molecule de valinomicină care conține șase aminoacizi dextrogitori (D) și șase levogitori (L), toate grupele laterale [-CH 3 -CH (CH 3) 2] sunt hidrofobe; b - reprezentarea schematică a configurației spațiale a complexului de valinomicină cu un ion de potasiu (în centru). Unele dintre grupările carbonil ale complexului formează legături de hidrogen cu atomii de azot, în timp ce altele formează legături de coordonare cu cationul (ionul de potasiu). Grupările hidrofobe formează sfera hidrofobă exterioară a complexului și asigură solubilitatea acestuia în faza hidrocarburică a membranei; 1 - atomi de carbon, 2 - atomi de oxigen, 3 - cation (ion de potasiu), 4 - atomi de azot, 5 - legături de hidrogen, 6 - legături de coordonare. Ionul de potasiu „captat” de molecula de valinomicină este transportat de această moleculă prin membrana celulară și eliberat. În acest fel, se asigură permeabilitatea selectivă a membranei celulare pentru ionii de potasiu.
În studiul P., celulele pentru un dizolvat în loc de un gradient de concentrație folosesc conceptul de diferență de concentrație a unei substanțe care difuzează pe ambele părți ale membranei și, în loc de coeficientul de difuzie, coeficientul de permeabilitate (P), care depinde si de grosimea membranei. Una dintre modalitățile posibile de pătrundere a substanțelor în celulă este dizolvarea lor în lipidele membranelor celulare, ceea ce este confirmat de existența unei relații direct proporționale între coeficientul de permeabilitate al unei clase mari de substanțe chimice. compuși și coeficientul de distribuție al substanței în sistemul ulei-apă. În același timp, apa nu se supune acestei dependențe, rata de penetrare a acesteia este mult mai mare și nu este proporțională cu coeficientul de distribuție în sistemul ulei-apă. Pentru apa si substantele cu greutate moleculara mica dizolvate in ea, cea mai probabila cale a P. este trecerea prin porii membranei. Astfel, difuzia substantelor prin membrana poate avea loc prin dizolvarea acestor substante in lipidele membranei; prin trecerea moleculelor prin pori polari formați din grupe polare încărcate de lipide și proteine, precum și prin trecerea prin pori neîncărcați. Sunt facilitate tipuri speciale și difuzie de schimb, asigurate de proteine și substanțe purtătoare solubile în grăsimi, care sunt capabile să lege substanța transportată pe o parte a membranei, să difuzeze cu ea prin membrană și să o elibereze pe cealaltă parte a membranei. Viteza de transfer a unei substanțe prin membrană în cazul difuziei facilitate este mult mai mare decât în difuziunea simplă. Rolul purtătorilor de ioni specifici poate fi îndeplinit de unele antibiotice (valinomicină, nigericină, monensin și o serie de altele), care se numesc ionofori (vezi Ionofori). A fost descifrată organizarea moleculară a complexelor antibioticelor ionoforice cu cationi. În cazul valinomicinei (Fig. 1), s-a arătat că după legarea de cationul de potasiu, molecula peptidică își schimbă conformația, căpătând forma unei brățări cu diametrul interior de cca. 0,8 nm, în Krom ionul de potasiu este reținut ca urmare a interacțiunilor ion-dipol.
Un tip comun de P. pasiv al membranelor celulare pentru substanțele polare este P. prin pori. Deși observarea directă a porilor din stratul lipidic al membranei este o sarcină dificilă, datele experimentale indică existența lor reală. Datele despre proprietățile osmotice ale celulelor mărturisesc și ele în favoarea existenței reale a porilor. Valoarea presiunii osmotice în soluțiile din jurul celulei poate fi calculată prin formula:
π=σCRT,
unde π - presiunea osmotică; C este concentrația substanței dizolvate; R este constanta gazului; T este temperatura absolută; σ este coeficientul de reflexie. Dacă viteza de trecere a unei molecule de solut prin membrană este proporțională cu viteza de trecere a moleculelor de apă, atunci mărimea forțelor va fi aproape de zero (nu există nicio modificare osmotică a volumului celulei); dacă membrana celulară este impermeabilă la o substanță dată, atunci valoarea lui σ tinde spre 1 (modificarea osmotică a volumului celulei este maximă). Viteza de pătrundere a moleculelor prin membrana celulară depinde de mărimea moleculei și astfel, prin selectarea moleculelor de o anumită dimensiune și observând modificarea volumului celulei într-o soluție dintr-o anumită substanță, se poate determina dimensiunea celulei. porii. De exemplu, membrana axonului calmarului este ușor permeabilă la moleculele de glicerol, care au o rază de cca. 0,3 nm, dar permeabil la substanțe cu dimensiuni moleculare mai mici (tabel). Experimente similare cu alte celule au arătat că dimensiunile porilor în membranele celulare, în special, în membranele eritrocitelor, Escherichia coli, celulele epiteliale intestinale etc., se potrivesc destul de precis în intervalul 0,6-0,8 nm.
Celulele și țesuturile vii se caracterizează printr-un alt mod de pătrundere a substanțelor în celulă și din ea - transportul activ al substanțelor. Transportul activ este transferul unei substanțe printr-o membrană celulară (sau intracelulară) (transport activ transmembranar) sau printr-un strat de celule (transport activ transcelular) care curge împotriva unui gradient electrochimic (vezi Gradient). adică cu cheltuirea energiei libere a corpului (vezi Metabolism și energie). Sistemele moleculare responsabile de transportul activ al substanțelor sunt localizate în membrana celulară (sau intracelulară). În membranele citoplasmatice ale celulelor implicate în transportul ionic activ - celule musculare, neuroni, eritrocite, celule renale - există o cantitate semnificativă de enzimă Na +, o ATPază independentă, care este implicată activ în mecanismele de transport ionic (vezi Ion). Transport). Mecanismul de funcționare al acestei enzime este cel mai bine studiat pe eritrocite și axoni, care au o capacitate pronunțată de a acumula ioni de potasiu și de a elimina (pompa) ionii de sodiu. Se presupune că eritrocitele conțin un dispozitiv molecular - o pompă de potasiu-sodiu (pompa de potasiu-sodiu), care asigură absorbția selectivă a ionilor de potasiu și îndepărtarea selectivă a ionilor de sodiu din celulă, iar elementul principal al acestei pompe este Na +, K+-ATPaza. Studiul proprietăților enzimei a arătat că enzima este activă numai în prezența ionilor de potasiu și sodiu, ionii de sodiu activând enzima din partea citoplasmei și ionii de potasiu din partea soluției înconjurătoare. Un inhibitor specific al enzimei este glicozidul cardiac ouabaina. Au fost găsite și alte ATPaze de transport, în special, care transportă ioni de Ca +2.
În membranele mitocondriilor, este cunoscut un sistem molecular care asigură pomparea ionilor de hidrogen, enzima H + -ATP-aza, iar în membranele reticulului sarcoplasmatic, enzima Ca++ -ATP-aza. Mitchell (P. Mitchell) - autorul teoriei chimiosmotice a fosforilării oxidative în mitocondrii (vezi Fosforilarea) - a introdus conceptul de „transport secundar al substanțelor”, care se realizează datorită energiei potențialului membranei și (sau) gradientul de pH. Dacă pentru ATPazele ionice, mișcarea antigradient a ionilor și utilizarea ATP sunt asigurate de același sistem enzimatic, atunci în cazul transportului activ secundar, aceste două evenimente sunt furnizate. sisteme diferiteși pot fi separate în timp și spațiu.
Pătrunderea în celule a macromoleculelor mari de proteine, acizi nucleici. enzimele celulare și celulele întregi se efectuează conform mecanismului de fagocitoză (captarea și absorbția particulelor solide mari de către celulă) și pinocitoză (captarea și absorbția de către o parte a suprafeței celulare a fluidului înconjurător cu substanțe dizolvate în ea).
P. membranele celulare este mai importantă pentru funcționarea celulelor și țesuturilor.
Transportul activ al ionilor și absorbția însoțitoare a apei în celulele epiteliului renal are loc în tubii proximali ai rinichiului (vezi Rinichi). Până la 1800 de litri de sânge trec zilnic prin rinichii unui adult. În același timp, proteinele sunt filtrate și rămân în sânge, 80% din săruri și apă, precum și toată glucoza, sunt returnate în sânge. Se crede că cauza principală a acestui proces este transportul activ transcelular al ionilor de sodiu, furnizat de ATP-aza dependentă de Na+ K+, localizată în membranele celulare ale epiteliului bazal. Dacă în canalul tubului proximal renal concentrația ionilor de sodiu este de cca. 100 mmol / l, apoi în interiorul celulei nu depășește 37 mmol / l; ca urmare, fluxul pasiv al ionilor de sodiu este direcționat în celulă. Pătrunderea pasivă a cationilor în citoplasmă este facilitată și de prezența unui potențial de membrană (suprafața interioară a membranei este încărcată negativ). Acea. ionii de sodiu pătrund în celulă pasiv în conformitate cu concentrația și gradienții electrici (vezi Gradient). Eliberarea ionilor din celulă în plasma sanguină se realizează în funcție de concentrație și gradienți electrici. S-a stabilit că tocmai în membrana bazală este localizată pompa de sodiu-potasiu, care asigură îndepărtarea ionilor de sodiu. Se presupune că anionii de clorură se deplasează după ionii de sodiu prin spațiul intercelular. Ca urmare, presiunea osmotică a plasmei sanguine crește, iar apa din canalul tubului începe să curgă în plasma sanguină, asigurând reabsorbția sării și apei în tubii renali.
Sunt folosite diferite metode pentru a studia P. pasiv și activ. Metoda atomilor marcați a devenit utilizată pe scară largă (vezi Izotopi, Medicamente radioactive, Cercetarea radioizotopilor). Izotopii 42 K, 22 Na și 24 Na, 45 Ca, 86 Rb, 137 Cs, 32 P și alții sunt utilizați pentru a studia P. ionic al celulelor; a studia P. apei - apa cu deuteriu sau tritiu, precum si apa marcata cu oxigen (18O); pentru studiul P. zaharuri și aminoacizi - compuși marcați cu carbon 14 C sau sulf 35 S; pentru studiul proteinelor P. - preparate iodate marcate cu 1 31 I.
Coloranții vitali sunt aplicați pe scară largă la cercetările lui P.. Esența metodei este de a observa la microscop viteza de penetrare a moleculelor de colorant în celulă. Pentru majoritatea coloranților vitali (roșu neutru, albastru de metilen, rodamină etc.), se fac observații în partea vizibilă a spectrului. De asemenea, sunt utilizați compuși fluorescenți, printre care fluoresceina de sodiu, clortetraciclină, murexida și altele.În studiul mușchilor, s-a demonstrat că pigmentarea moleculelor de colorant depinde nu numai de proprietățile membranei celulare, ci și de capacitatea de sorbție. a structurilor intracelulare, cel mai adesea proteine si acizi nucleici.-t, cu care se leaga colorantii.
Metoda osmotică este utilizată pentru studiul P. al apei și al substanțelor dizolvate în ea. În același timp, folosind un microscop sau măsurând împrăștierea luminii a unei suspensii de particule, se observă o modificare a volumului celulelor în funcție de tonicitatea soluției din jur. Dacă celula este într-o soluție hipertonă, atunci apa din aceasta intră în soluție și celula se micșorează. Efectul opus se observă în soluția hipotonică.
Din ce în ce mai mult, metode potențiometrice sunt folosite pentru studiul P. membranelor celulare (vezi Metoda de cercetare a microelectrodului, Conductibilitatea electrică a sistemelor biologice); O gamă largă de electrozi specifici ionilor face posibilă studierea cineticii de transport a multor ioni anorganici (potasiu, sodiu, calciu, hidrogen etc.), precum și a unor ioni organici (acetați, salicilați etc.). Toate tipurile de P. ale membranelor celulare sunt într-o oarecare măsură caracteristice sistemelor membranare tisulare multicelulare - pereții vaselor de sânge, epiteliul rinichilor, membrana mucoasă a intestinelor și stomacului. În același timp, P. vaselor se caracterizează prin unele trăsături care se manifestă prin încălcarea P. vasculară (vezi mai jos).
Fiziologia patologică a permeabilității vasculare
Termenul „permeabilitate vasculară” a fost folosit pentru a se referi la metabolismul histohematic și transcapilar, distribuția substanțelor între sânge și țesuturi, P. țesut, tranziția hemolimfatică a substanțelor și alte procese. Unii cercetători folosesc acest termen pentru a se referi la funcția trofică a structurilor de țesut capilar-conjunctiv. Ambiguitatea utilizării termenului a fost unul dintre motivele inconsecvenței opiniilor cu privire la o serie de probleme, în special cele legate de reglarea P vasculară. În anii '70. Secolului 20 termenul „permeabilitate vasculară” a început să folosească Ch. arr. pentru a indica permeabilitatea selectivă sau funcția de barieră de transport a pereților microvaselor de sânge. Există tendința de a atribui P. vasculară și P. pereții nu numai ai microvaselor (sânge și limfa), ci și a vaselor mari (până la aortă).
Se observă modificări ale P. vasculare hl. arr. sub forma unei creșteri a P. selectivă pentru macromolecule și celule sanguine. Un exemplu tipic în acest sens este exsudația (vezi). Scăderea P. vasculară este legată în general de impregnarea proteinică și inspizia ulterioară a pereților vasculari, care se observă, de exemplu, la o hipertensiune idiopatică (vezi).
Există o opinie cu privire la posibilitatea tulburării de către P. a peretelui vascular în principal în direcția interstițiului sau dinspre interstițiu în sânge. Cu toate acestea, mișcarea predominantă a substanțelor într-o direcție sau alta în raport cu peretele vascular nu dovedește încă legătura acesteia cu starea funcției de barieră-transport a peretelui vascular.
Principii pentru studierea tulburărilor de permeabilitate vasculară
O evaluare a stării P. vasculare trebuie efectuată ținând cont de faptul că peretele vascular asigură o distincție și o legătură funcțională între două medii adiacente (sânge și mediul interstițial), care sunt principalele componente ale mediului intern al corp (vezi). Schimbul între aceste medii adiacente în ansamblu se realizează datorită microcirculației (vezi Microcirculația), iar peretele vascular cu funcția sa de barieră de transport acționează doar ca bază a specializării organelor a metabolismului histohematologic. Prin urmare, metoda de studiere a stării P. vasculare poate fi considerată adecvată doar atunci când permite aprecierea parametrilor calitativi ai metabolismului histohematic, ținând cont de specificul lor de organ și indiferent de starea microcirculației organelor și de natura proceselor metabolice care se formează. în afara peretelui vascular. Din acest punct de vedere, cea mai adecvată dintre metodele existente este metoda microscopică electronică pentru studierea P. vasculară, care face posibilă observarea directă a căilor și mecanismelor de penetrare a substanțelor prin peretele vascular. Deosebit de fructuoasă a fost combinația dintre microscopia electronică cu așa-numita. indicatoare de urmărire sau trasoare, care marchează căile deplasării lor prin peretele vascular. Ca atare indicatori, pot fi utilizate orice substanțe netoxice detectate prin microscopie electronică sau tehnici speciale (histochimice, radioautografice, imunocitochimice etc.). În acest scop, se utilizează feritina proteică cu conținut de fier, diverse enzime cu activitate peroxidază, cărbune coloidal (cerneală neagră purificată) etc.
Dintre metodele indirecte de studiere a stării funcției de barieră-transport a pereților vaselor de sânge, cea mai utilizată este înregistrarea penetrării prin peretele vascular a indicatorilor naturali sau artificiali care penetrează slab sau nu penetrează peretele sub conditii normale. În încălcarea microcirculației, care este adesea observată cu încălcarea P. vasculară, aceste metode pot fi neinformative și apoi ar trebui combinate cu metode de monitorizare a stării de microcirculație, de exemplu. folosind biomicroscopie sau indicatori ușor de difuzat, al căror schimb histohematic nu depinde de starea P. vasculară și de metabolismul tisular. Dezavantajul tuturor metodelor indirecte bazate pe înregistrarea acumulării de substanțe indicator în afara patului vascular este necesitatea de a lua în considerare masa de factori care pot afecta semnificativ nivelul indicatorului în zona studiată. În plus, aceste metode sunt destul de inerțiale și nu permit studierea modificărilor pe termen scurt și reversibile ale P. vascular, în special în combinație cu o modificare a microcirculației. Aceste dificultăți pot fi depășite parțial prin utilizarea metodei vaselor marcate, care se bazează pe determinarea pătrunderii în peretele vascular a unui indicator slab difuzabil care se acumulează în perete și îl pătează. Locurile pictate (etichetate) ies la lumină cu ajutorul unui microscop cu lumină și sunt dovada încălcării P. a unui endoteliu. Ca indicator, poate fi utilizat cărbunele coloidal, care formează acumulări întunecate ușor detectabile în locurile de încălcare gravă a barierei endoteliale. Modificările în activitatea de transport microvezicular nu sunt înregistrate prin această metodă și este necesar să se utilizeze alți indicatori transportați prin endoteliu de către microvezicule.
Posibilitățile de a studia tulburările P. vasculare într-un cadru clinic sunt mai limitate, deoarece majoritatea metodelor bazate pe utilizarea indicatorilor micromoleculari ușor de difuzat (inclusiv radioizotopi) nu permit să se judece fără ambiguitate starea funcției de barieră de transport a pereții vaselor de sânge.
O metodă bazată pe determinarea diferențelor cantitative ale conținutului de proteine din probele de sânge arterial și venos prelevate simultan este utilizată relativ pe scară largă (vezi testul Landis). Atunci când se calculează procentul de pierdere de proteine din sânge în timpul trecerii acestuia de la patul arterial la cel venos, este necesar să se cunoască procentul de pierdere de apă, care este determinat de diferența de hematocrit al sângelui arterial și venos. În cercetările lor asupra oameni sanatosi V. P. Kaznacheev și A. A. Dzizinsky (1975), ca indicatori ai P. normală a vaselor membrului superior, au derivat următoarele valori: pentru apă, o medie de 2,4-2,6%, pentru proteine, 4-4,5%, adică atunci când trecând prin patul vascular 100 ml de sânge în limfă. albia râului intră în cca. 2,5 ml apă și 0,15-0,16 g proteine. În consecință, în corpul uman ar trebui să se formeze cel puțin 200 de litri de limfă pe zi, ceea ce este de zece ori mai mare decât valoarea reală a producției zilnice de limfă în corpul unui adult. În mod evident, dezavantajul metodei este presupunerea că, potrivit Krom, diferențele de hematocrit al sângelui arterial și venos se explică doar printr-o modificare a conținutului de apă din sânge datorită ieșirii acesteia din patul vascular.
Într-o pană practică, starea P. vasculară regională este adesea judecată după prezența acumulărilor interstițiale sau cavitare de lichid liber bogat în proteine. Cu toate acestea, atunci când se evaluează starea P. vasculară, de exemplu. în cavitatea abdominală se poate face o concluzie eronată, întrucât microvasele metabolice ale acestor organe și țesuturi sunt caracterizate în mod normal de P. mare pentru macromolecule din cauza discontinuității sau porozității endoteliului lor. O creștere a presiunii de filtrare în astfel de cazuri duce la formarea unei efuzii bogate în proteine. Sinusurile venoase și sinusoidele sunt deosebit de permeabile la moleculele de proteine.
Trebuie remarcat faptul că producția crescută de proteine plasmatice în țesut și dezvoltarea edemului tisular (vezi) nu însoțesc întotdeauna o creștere a P. vasculară. Microvasele (capilare și venule), al căror endoteliu este în mod normal slab permeabil la macromolecule. , dobândesc defecte endoteliale; prin aceste defecte intră cu ușurință în spațiul subendotelial introdus în fluxul sanguin indicatori - macromolecule și microparticule. Cu toate acestea, nu există semne de edem tisular - așa-numitul. formă edematoasă de afectare a permeabilității vasculare. Un fenomen similar se observă, de exemplu, în mușchii animalelor în timpul dezvoltării unui proces neurodistrofic în ei asociat cu secțiunea nervului motor. Modificări similare în țesuturile umane sunt descrise, de exemplu, în timpul îmbătrânirii și Diabet când așa-numitul capilare acelulare, adică microvase metabolice cu celule endoteliale descuamate parțial sau complet (nu există nici semne de edem tisular). Toate aceste fapte vorbesc, pe de o parte, despre relativitatea relației edemului tisular cu creșterea P. vasculară, iar pe de altă parte, despre existența unor mecanisme extravasculare responsabile de distribuția apei și a substanțelor între sânge. și țesuturi.
Factori de afectare a permeabilității vasculare
Factorii de încălcare a permeabilității vasculare sunt împărțiți în mod convențional în două grupuri: exogeni și endogeni. Factorii exogeni de perturbare a P. vasculare de natură variată (fizică, chimică etc.) sunt la rândul lor împărțiți în factori care afectează direct peretele vascular și funcția sa de barieră de transport, de exemplu, histamina introdusă în patul vascular, diverse toxine. , etc. .), și factori de încălcare P. de acțiune indirectă, al căror efect este mediat prin factori endogeni.
Un număr mare de altele, în special prostaglandine (vezi), au început să fie atribuite factorilor endogeni deja cunoscuți ai tulburării P. vasculare (histamină, serotonină, kinine), iar acestea din urmă nu numai că cresc P. vascular, ci și intensifică acțiunea altor factori; mulți dintre factorii endogeni sunt produși de diverse sisteme enzimatice ale sângelui (sistemul factorului Hageman, sistemul complementului etc.).
Creșterea P. vasculară și complexele imune. Din factorul responsabil pentru creșterea „întârziată” a P. vasculară în timpul dezvoltării fenomenului Arthus, Yosinaga (1966) a evidențiat pseudoglobulina; Kuroyanagi (1974) a descoperit un nou factor P., desemnat de el drept Ig-PF. Prin proprietățile sale, diferă semnificativ de histamina, kinine, anafilatoxină și kalikreină, acționează mai mult decât histamina și bradikinina și este inhibată de vitaminele K1 și K2.
Mulți factori de perturbare a P. vasculare sunt produși de leucocite. Astfel, o protează este asociată cu suprafața neutrofilelor, care formează un mediator peptidic neutru din proteinele plasmatice care crește P vascular. Substratul proteic al proteazei are un mol. greutate (masă) 90.000 și diferită de kininogen.
Lizozomii și granulele specifice de celule sanguine conțin proteine cationice care pot perturba P vascular. Acțiunea lor este mediată de histamina mastocitară.
Diferiți factori endogeni de perturbare a P. vasculare acționează în țesături simultan sau secvenţial, determinând în. vascular P. schimbări de fază. În acest sens, se disting modificările precoce, întârziate și tardive ale P. vasculară Faza precoce este faza de acțiune a histaminei (vezi) și a serotoninei (vezi). A doua fază se dezvoltă după o perioadă de bunăstare imaginară, la 1-3 ore după leziunea primară - o fază întârziată, sau întârziată; dezvoltarea sa este cauzată de acțiunea kininelor (vezi) sau a prostaglandinelor. Dezvoltarea acestor două faze depinde de nivelul complementului și este inhibată de serul imun anticomplementar. La o zi după deteriorare, se dezvoltă a treia fază, asociată cu acțiunea enzimelor cito- și proteolitice eliberate din lizozomii leucocitelor și limfocitelor. În funcție de natura agentului dăunător primar, numărul de faze poate fi diferit. Într-o fază precoce vasculară P. este ruptă de hl. arr. la nivelul venulelor, în fazele ulterioare procesul se extinde până la patul capilar și arteriole.
Recepția factorilor de permeabilitate de către peretele vascular. Factorii endogeni ai tulburării P. reprezintă cel mai important grup de cauze ale tulburării vasculare P.. Unii dintre ei sunt în formă gata preparată în țesuturi (histamină, serotonină) și, sub influența diferitelor influențe patogene, sunt eliberate din depozit, care sunt mastocite și celule sanguine (bazofile, trombocite). Alți factori sunt produsul diferitor biochimie. sisteme atât la locul deteriorării primare, cât și la distanță de acesta.
Întrebările privind originea factorilor P. sunt în sine importante pentru rezolvarea problemelor practice de prevenire și tratare a tulburărilor P. vasculare. Cu toate acestea, apariția factorului P. nu este încă suficientă pentru P. vasculare. tulburare „Văzut”, adică prescris, de peretele vascular (cu excepția cazului în care are o capacitate de destructurare precum agenții citolitici). Se știe, de exemplu, că histamina, introdusă în circulația generală, perturbă P. vascular doar în anumite organe și țesuturi, în timp ce în alte țesuturi (creier, țesut pulmonar, endoneur etc.) nu este eficientă. La broaște, introducerea serotoninei și a bradikininei în patul vascular nu provoacă deloc perturbarea P vasculară. Cu toate acestea, motivele ineficienței histaminei în ambele cazuri sunt diferite.
Conform datelor moderne, endoteliul microvaselor metabolice ale animalelor și oamenilor cu sânge cald este sensibil la un număr mare de agenți diferiți, adică se caracterizează printr-o capacitate mare de receptor. În ceea ce privește histamina, unul dintre principalii factori ai P., care provoacă o tulburare acută și semnificativă (deși pe termen scurt) a P. vasculară, datele experimentale indică prezența în endoteliu a două tipuri de receptori de histamină H1 și H2, care joacă roluri diferite în mecanismul de acțiune al histaminei. Este stimularea receptorilor H1 care duce la perturbarea P. vasculară, care este caracteristică acțiunii histaminei.
Sub acțiunea unor factori endogeni P., în special histaminei, se observă tahifilaxia (vezi) și utilizarea repetată (după 30 de minute) a agentului nu mai încalcă P. vasculară. în unele cazuri, acesta poate fi cazul. În cazul histaminei, mecanismul tahifilaxiei, conform unor rapoarte, are o localizare extra-receptor. Acest lucru este dovedit, în special, de dezvoltarea tahifilaxiei încrucișate, când utilizarea histaminei duce la dezvoltarea rezistenței endoteliale nu numai la histamină în sine, ci și la sărurile de lantan care ocolesc receptorii. Apariţia tahifilaxiei încrucişate poate fi unul dintre motivele ineficienţei factorilor P. individuali care acţionează simultan sau secvenţial.
Baze ultrastructurale și mecanisme efectoare ale tulburărilor de permeabilitate vasculară
Orez. Fig. 2. Modalităţi şi mecanisme ale metabolismului transcapilar în condiţii normale (a) şi patologie (b): 1 - difuzie transcelulară; 2 - difuzie și ultrafiltrare în zona joncțiunilor intercelulare dense; 3 - difuzie și ultrafiltrare în zona conexiunilor intercelulare simple; 4 - transport microvezicular ocolind joncțiunile intercelulare strânse; 3a și 4a - canale intercelulare patologice de tip „goluri de histamină”; 5 - transport microvezicular; 6 - formarea unui canal transcelular prin fuziunea microveziculelor; 7 - vacuole fagocitare în pericite; 8 - microparticule ale indicatorului de permeabilitate vasculară (BM - membrana bazală, EN1, EN2, EN3 - endoteliocite, PC - pericite).
Studiile microscopice electronice au arătat că morfolul. baza creșterii P. vasculare este formarea de canale largi în zona conexiunilor intercelulare în endoteliu (Fig. 2). Astfel de canale, sau „scurgeri”, sunt adesea numite crăpături de histamină, deoarece formarea lor este tipică acțiunii asupra peretelui vascular al histaminei și a fost studiată mai întâi în detaliu în timpul acțiunii sale. Fisurile de histamina sunt formate de hl. arr. în pereții venulelor acelor organe și țesuturi în care nu există bariere histohematice cu permeabilitate scăzută, cum ar fi bariera hemato-encefalică etc. Discrepanțele locale în contactele intercelulare au fost găsite în tulburări de neuroreglare, mecanice, termice, chimice și alte tipuri de afectarea țesuturilor, sub acțiunea diverșilor bioregulatori (serotonina, bradikinina, prostaglandinele E1 și E2 etc.). Încălcarea contactelor intercelulare are loc, deși cu mare dificultate, în capilare și arteriole și chiar în vasele mai mari. Ușurința formării golurilor de histamină este direct proporțională cu slăbiciunea structurală inițială a conexiunilor intercelulare, marginea crește în timpul trecerii de la arteriole la capilare și de la capilare la venule, atingând un maxim la nivelul venulelor postcapilare (pericitice).
Ineficacitatea histaminei în perturbarea P. vasculară a unor organe este bine explicată tocmai din punctul de vedere al dezvoltării joncțiunilor strânse în endoteliul microvaselor acestor organe, de exemplu. creier.
În termeni teoretici și practici, problema mecanismelor efectoare care stau la baza formării defectelor structurale precum golurile de histamină este importantă. Aceste schimbări ultrastructurale sunt tipice pentru faza initiala inflamația acută (vezi), când, conform lui I. I. Mechnikov (1891), o creștere a P. vasculară este adecvată din punct de vedere biologic, deoarece aceasta asigură o ieșire crescută a fagocitelor la locul leziunii. Se poate adăuga că o producție de plasmă crescută în astfel de cazuri este, de asemenea, recomandabilă, deoarece în acest caz anticorpii și agenții de protecție nespecifici sunt livrați în focar. Astfel, o creștere a P. vasculară în focarul inflamației poate fi considerată ca o stare specifică a funcției de barieră-transport a pereților microvaselor, adecvată noilor condiții de existență a țesutului, și o modificare a P. vasculară. în timpul inflamației și situațiilor similare nu este o încălcare, ci ca o nouă stare funcțională care contribuie la restabilirea homeostaziei tisulare perturbate. Trebuie avut în vedere faptul că în unele organe (ficat, splină, măduvă osoasă), unde, în conformitate cu caracteristicile funcțiilor organelor, există un flux metabolic continuu de celule și macromolecule, „scurgerile” intercelulare sunt formațiuni normale și permanente. , care sunt lacune de histamină exagerate, dar spre deosebire de golurile de histamină adevărate sunt capabile de existență pe termen lung. Adevăratele goluri de histamină se formează chiar în primele secunde după expunerea la mediatorii inflamației acute pe endoteliu și, în cea mai mare parte, după 10-15 minute. sunt inchise. Mecanismul de formare a golurilor de histamină are o natură protectoare, determinată filogenetic și este asociat cu o reacție stereotipată la nivel celular, declanșată de stimularea diferitelor tipuri de receptori.
Natura acestei reacții stereotipe a rămas neexplorată multă vreme. I. I. Mechnikov credea că o creștere a P. vasculară în timpul inflamației este asociată cu o reducere a celulelor endoteliale. Cu toate acestea, mai târziu s-a constatat că endoteliocitele din vasele animalelor cu sânge cald nu aparțin categoriei de celule care își schimbă în mod activ forma ca celulele musculare. Rowley (D. A. Rowley, 1964) a sugerat că divergența endoteliocitelor este o consecință a creșterii presiunii intravasculare și a supraîntinderii asociate a endoteliului. Măsurătorile directe au dovedit inacceptabilitatea acestei ipoteze în raport cu venule și capilare, totuși, pentru vasele arteriale ea are o anumită valoare, deoarece dacă activitatea tonică a membranei musculare este perturbată, presiunea intravasculară ridicată poate provoca într-adevăr supraîntinderea endoteliului și deteriorarea contactelor intercelulare. Dar, în acest caz, apariția golurilor de histamină în intime nu este întotdeauna asociată cu acțiunea presiunii transmurale. Robertson și Kairallah (A. L. Robertson, P. A. Khairallah, 1972) în experimente pe un segment izolat al aortei abdominale a unui iepure au arătat că se formează goluri largi în endoteliu sub influența angiotensinei II în locurile de rotunjire și scurtare a endoteliocitelor. Morfol similar. De asemenea, s-au găsit schimbări în endoteliul microvaselor metabolice ale pielii cu aplicare topică a angiotensinei II, prostaglandinei E1 și trigliceridelor serice.
O. V. Alekseev și A. M. Chernukh (1977) au descoperit în endoteliocitele microvaselor metabolice capacitatea de a crește rapid conținutul din citoplasmă a structurilor microfibrilare similare în morfolul lor. caracteristici cu microfilamente de actină. Acest fenomen reversibil (așa-numitul fenomen de structuralizare operațională a aparatului microfibrilar) se dezvoltă sub influența factorilor care provoacă formarea de goluri intercelulare largi. Reversibilitatea fenomenului în cazul utilizării histaminei îngreunează depistarea și explică bine durata scurtă și reversibilitatea existenței lacunelor histaminei. Cu ajutorul citocalazinei-B, care blochează formarea microfibrilelor de actină, se dezvăluie semnificația patogenetică a acestui fenomen în mecanismul de formare a golurilor histaminice intercelulare. Aceste fapte indică faptul că endoteliocitele au o capacitate latentă de a se contracta, care se realizează în condițiile în care nivelul anterior de P. vasculară este inadecvat și este necesară o modificare relativ rapidă și reversibilă. Schimbarea P. vasculară acţionează, astfel, ca un act special de biol. reglare, care asigură adaptarea funcției de barieră-transport a endoteliului vascular în concordanță cu noile nevoi locale care au apărut brusc în legătură cu modificările condițiilor activității vitale a țesuturilor.
Prezența în țesuturi a mecanismului de schimbare în P. vasculară poate fi atribuită așa-numitului. factori de risc, deoarece funcționarea acestui mecanism în condiții inadecvate poate provoca o încălcare a homeostaziei țesuturilor și a funcției organelor, și nu o manifestare a acțiunii mecanismelor adaptiv-protectoare. Principalele moduri de perturbare a P. vasculară sunt prezentate pe schemă. Modificările P. vasculare se bazează pe mecanisme care nu numai că duc la formarea canalelor intercelulare (goluri de histamină), dar afectează și activitatea suprafeței celulare (adică microvezicularea și transportul microvezicular, vacuolizarea și formarea de microbule). Rezultatul poate fi perforarea endoteliocitelor cu formarea de canale transcelulare mai mult sau mai puțin extinse și pe termen lung.
O mare importanță în mecanismele de perturbare a P. vasculară este atașată modificărilor locale ale sarcinii electrice de suprafață, în special pe membranele care închid porii din capilarele fenestrate (de exemplu, glomeruli renali). Potrivit unor date, schimbarea în sarcină singură poate fi baza pentru creșterea randamentului de proteine din capilarele glomerulare. Acea. se demonstrează limitarea teoriei porilor; În condiții de patologie, efectul de creștere a porozității endoteliului poate fi realizat în diferite moduri: prin formarea de canale intercelulare precum golurile de histamină; transport microvezicular și intravacuolar crescut; perforarea celulelor endoteliale pe baza microveziculării crescute, vacuolizării sau formării de microbule în endoteliu; distrugerea microfocală a endoteliocitelor; descuamarea endoteliocitelor; schimbare fiz.-chim. proprietățile suprafeței endoteliocitelor etc. (vezi Microcirculația ]]). Același efect poate fi obținut și datorită mecanismelor extra-perete, în special, datorită unei modificări a capacității de legare a macromoleculelor sanguine, cu care aproape toți indicatorii cunoscuți utilizați pentru a evalua starea P. vasculară interacționează.mecanismele enumerate. Deci, de exemplu, histamina crește porozitatea peretelui vascular datorită formării de goluri de histamină în endoteliul venulelor, precum și prin influențarea suprafeței endoteliocitelor și a proceselor de transport asociate activității sale și transformărilor ultrastructurale (formarea de transcelulare). pori, fenestrari, microtubuli etc.). Trebuie luat în considerare faptul că acest lucru modifică adesea grosimea endoteliocitelor și adâncimea golurilor intercelulare, ceea ce poate afecta semnificativ permeabilitatea peretelui vascular ca barieră de difuzie. Problema comportamentului în condiții de patologie biochimică nu a fost deloc studiată. mecanisme care împiedică sau, dimpotrivă, favorizează pătrunderea substanţelor prin peretele vascular, în special a celor biologic active. Se știe, de exemplu, că endoteliocitele capilarelor cerebrale au în mod normal o activitate enzimatică care distruge serotonina și astfel împiedică pătrunderea acesteia atât din sânge în creier, cât și în direcția opusă. Endoteliul capilarelor pulmonare contine kininaza II, care este localizata in veziculele micropinocitare si asigura distrugerea bradikininei si, in acelasi timp, conversia angiotensinei I in angiotensină II (hipertensiune). Astfel, endoteliul exercită un fel de control asupra echilibrului bioregulatorilor umorali și influențează activ metabolismul histohematic al acestor agenți.
Intervenția direcționată se realizează la trei niveluri (vezi diagrama). Primul nivel - impactul asupra procesului de formare a factorilor cauzali (receptibili) - practic nu este utilizat, deși există medicamente separate care pot acționa la acest nivel. De exemplu, reserpina afectează depunerea factorilor de perturbare ai P. în mastocite, care sunt principala sursă de mediatori ai inflamației acute (histamină și serotonina); agenții antiprostaglandine inhibă sinteza prostaglandinelor - acid acetilsalicilic etc.
Al doilea nivel este cel principal în practica dezvoltării mijloacelor de prevenire și tratare a tulburărilor P vasculare. Corespunde procesului de recepție a factorului cauzal. Un număr semnificativ de medicamente antihistaminice, antiserotoninice și antibradikinine sunt utilizate pentru a preveni tulburările P. vasculare cauzate de mediatorii corespunzători. Avantajul și în același timp dezavantajul acestor medicamente, acționând prin blocarea receptorilor specifici, este specificitatea lor ridicată. O astfel de specificitate le face ineficiente în condițiile multiplicității etiol. factori care acţionează simultan sau secvenţial, ceea ce se observă de obicei într-o pană. practică. De asemenea, este important ca excluderea acțiunii unuia sau mai multor factori care determină dezvoltarea unei faze a tulburării P. vasculare să nu excludă dezvoltarea fazelor ulterioare. Aceste neajunsuri pot fi depășite prin intervenție la al treilea nivel.
Al treilea nivel este efectul asupra mecanismelor efectoare intracelulare (subcelulare) prin care se realizează direct acțiunea factorilor lui P. și sunt aceleași pentru acțiunea diferiților agenți patogeni. Realitatea și eficacitatea acestei abordări pot fi demonstrate experimental prin utilizarea unei substanțe (citocalazina-B) care inhibă fenomenul de structuralizare operațională a aparatului microfibrilar în endoteliocite (formarea gelului de actină și a microfibrilelor de actină).
Într-o pană În practică, pentru a normaliza creșterea P. vasculară, se utilizează vitamina P (vezi Bioflavonoide) și săruri de calciu. Cu toate acestea, aceste medicamente nu pot fi considerate ca fiind specifice pentru a stabili. agenți care încalcă P. vasculară, deși au un efect general de întărire asupra barierelor histohematice, a membranelor și a peretelui vaselor de sânge în special.
Diferiți factori P. endogeni pot fi utilizați pentru a crește P. vascular, de exemplu. histamină sau substanțe care le eliberează din depozitele de țesuturi.
Bibliografie: Alekseev O. V. Homeostazia microcirculatorii, în cartea: Homeostasis, ed. P. D. Horizontova, p. 278, M., 1976; Antonov VF Lipidele și permeabilitatea ionică a membranelor, M., 1982; Membrane biologice, ed. D. S. Parsons, trad. din engleză, M., 1978; D e Robert tis E., Novinsky V. și S și e cu F. Biologia celulei, trad. din engleză, M., 1967; Celulă vie, trad. din engleză, ed. G. M. Frank, p. 130, Moscova, 1962; K a z-nacheevV.P. şi Dz şi z şi N cu to şi y A. A. Clinical pathology of transcapilary exchange, M., 1975; Picior ușor E. Fenomene de transfer în sistemele vii, trans. din engleză, M., 1977; Lakshminaraya nay și x N. Electrozi cu membrană, trans. din engleză, L., 1979; Lev A. A. Modeling of ionic selectivity of cellular membranes, L., 1976; Ovchinnikov Yu. A., Ivanov V. T. și III la r despre b A. M. Complexoni activi pe membrană, M., 1974; Structura și funcția celulei, trans. din engleză, ed. G. M. Frank, p. 173, M., 1964; Troshin A. S. Problema permeabilității celulare, M. - L., 1956; Chernukh A. M., Alexandrov P. N. și Alekseev O. V. Microcirculation, M., 1975; Di Rosa M., Giroud J. R. a. W 1 1-loughby D. A. Studii ale mediatorilor răspunsului inflamator acut indus la șobolani în diferite locuri de către carra-geenan și terebentină, J. Path., v. 104, p. 15, 1971; M a j n o G. a. P a 1 a-de G. E. Studies on inflammation, I. The effect of histamine and serotonin on vascu-lar permeability, an electron microscopic study, J. biophys. biochimie. Cytol., v. 11, p. 571, 1961; M a j n o G., S h e a S. M. a. Leventhal M. Contracția endotelială indusă de mediatori de tip histamină, J. Cell Biol., v. 42, p. 647, 1969: Shimamoto T. Contracția celulelor endoteliale ca mecanism cheie în aterogeneza și tratamentul aterosclerozei cu relaxanți ai celulelor endoteliale, în: Atherosclerosis III, ed. de G. Schettler a. A. Weizel, p. 64, V.-N. Y., 1974.
B. F. Antonov; O. V. Alekseev (cale. Fiz.).
Metodele de curățare a membranei se bazează pe o permeabilitate diferită a membranei pentru componentele amestecului de gaz care se curăță.[ ...]
Permeabilitatea selectivă a membranelor în procesul de ultrafiltrare se explică printr-un mecanism pur de separare prin sită - particulele de impurități care sunt mai mari decât dimensiunea porilor membranei nu trec prin membrană, doar apa este filtrată prin ea.[ .. .]
Selectivitatea și permeabilitatea membranelor trebuie luate în considerare în raport cu costurile de obținere a aerului îmbogățit cu oxigen. Costurile de separare a aerului depind de permeabilitate, selectivitate, parametri geometrici ai membranelor, performanța modulului, costul energiei electrice și alți factori. Costul aerului îmbogățit cu oxigen este estimat în raport cu oxigenul echivalent pur, definit ca cantitatea de oxigen pur necesară amestecării cu aerul (21% oxigen) pentru a obține aceeași cantitate și procent de oxigen care se obține în procesul de separare a gazelor. în cauză.[ ...]
Ultrafiltrarea este un proces membranar pentru separarea solutiilor a caror presiune osmotica este scazuta. Această metodă este utilizată în separarea substanțelor cu greutate moleculară relativ mare, a particulelor în suspensie, a coloizilor. Ultrafiltrarea în comparație cu osmoza inversă este un proces mai eficient, deoarece o permeabilitate mare a membranei se realizează la o presiune de 0,2-1 MPa.[ ...]
Spalare deseuri solide 434, 425 Permeabilitatea membranei 273 Strecurare 197 cl.[ ...]
Ionii de calciu au o mare influență asupra structurilor membranelor. Necesitatea ionilor de Ca2+ pentru a stabiliza membranele a fost subliniată de mult timp. S-a demonstrat că prezența ionilor de Ca2+ în soluția înconjurătoare este necesară pentru formarea unei membrane de suprafață pe o picătură endoplasmatică izolată din celulele interdistante ale carofitelor. Prezența Ca2+ la o concentrație de 10 4 M a favorizat formarea unei membrane de suprafață pe picătură, deși nu suficient de puternică; s-a format o membrană mai puternică la o concentrație de 10-3 M și în special 10 2 M. Când ionii de calciu sunt îndepărtați (de exemplu, atunci când sunt tratați cu chelați sau în absența Ca2 + în mediu), se notează mucilagiul firelor de păr radiculare. , și crește și permeabilitatea membranelor la alte substanțe.Ionii de Ca2 + se modifică și proprietățile electrice atât ale membranelor artificiale, cât și ale celor naturale, reducând densitatea de sarcină pe suprafața membranei.Lipsa de Ca duce la creșterea vacuolizării, modificări ale cromozomilor, ruptura membranelor ER și a altor compartimente intracelulare.[ ...]
Odată cu creșterea concentrației soluției separate, permeabilitatea membranelor scade, iar odată cu creșterea presiunii, crește. După procesul de purificare, se obține un filtrat, epuizat cu 90-99,5 ° / o în compușii inițiali și un concentrat trimis pentru prelucrare ulterioară.[ ...]
Răspunsul la acetilcolină și aminele biogene este de a modifica permeabilitatea membranelor la ioni și/sau de a induce sinteza mesagerilor secundari. Prezența cAMP, cGMP, Ca2+, precum și a enzimelor de sinteză și catabolism în celula vegetală și organitele acesteia, confirmă posibilitatea medierii locale.[ ...]
Deci, sub acțiunea EMR cu microunde (2,45 GHz), s-a constatat o creștere a permeabilității cationilor membranelor eritrocitare la temperatura camerei, în timp ce în absența EMR cu microunde, un efect similar se observă doar la o temperatură de 37 °C. [...]
Fondurile de metaboliți nu sunt distribuite uniform în întreaga celulă, ci separate prin membrane și localizate în compartimente separate (camere, compartimente). Compartimentele fondurilor metabolice ale celulei sunt interconectate prin fluxuri de transport. În conformitate cu permeabilitatea selectivă a membranelor, are loc o redistribuire spațială a intermediarilor și a produselor metabolice. De exemplu, într-o celulă, aprovizionarea cu ATP este menținută datorită legăturilor „orizontale” dintre procesele de formare a fosforului fotosintetic și oxidativ.[ ...]
concentrația soluției. Odată cu creșterea concentrației soluției separate, permeabilitatea membranelor scade din cauza creșterii presiunii osmotice a solventului și a efectului polarizării concentrației. Cu o valoare a criteriului Reynolds de 2000-3000, polarizarea concentrației este practic absentă, totuși, turbulizarea soluției este asociată cu recircularea sa multiplă, adică cu costuri energetice, și duce la acumularea de particule în suspensie în soluție și apariția murdărie biologică.[ ...]
O scădere a temperaturii apei, care duce la răcirea peștilor, duce, de asemenea, la o creștere a permeabilității membranelor, care își pierd capacitatea de a menține gradienții ionici. În acest caz, conjugarea reacțiilor enzimatice este perturbată, pompele ionice încetează să funcționeze, activitatea sistemului nervos central și periferic este întreruptă, activitatea aparatului cardiorespirator este inhibată, ceea ce în cele din urmă poate duce la dezvoltarea hipoxiei. În timpul supraîncălzirii sau răcirii peștilor care rezultă dintr-o schimbare bruscă a temperaturii într-un timp limitat, un anumit rol revine stresului osmotic din cauza unei încălcări a capacității organismului de a menține o anumită concentrație de ioni și proteine în sânge. De exemplu, o scădere a temperaturii de la 25 la 11 ° C provoacă dezvoltarea unei comei la tilapia ținută în apă dulce, însoțită de o scădere a concentrației de ioni de sodiu și clor și a proteinelor totale din sânge. Potrivit autorilor, moartea peștilor are loc din cauza dezvoltării colapsului osmoreglator și a inhibării funcției renale. O confirmare indirectă a acestei ipoteze poate fi prevenirea comei termice la peștii ținuți în apă de mare diluată, ceea ce este în concordanță cu observațiile anterioare privind creșterea rezistenței termice a peștilor datorită adăugării de ioni de sodiu, calciu și magneziu în apă. . Cu toate acestea, trebuie avut în vedere că cauzele morții peștilor la temperaturi ridicate sau scăzute sunt diferite și depind de durata și intensitatea efectului de temperatură.[ ...]
valoare PH. O modificare a pH-ului inițial are ca rezultat, de obicei, o scădere a permeabilității membranei. Efectul pH-ului asupra selectivității membranei este mic. Acizii volatili sunt slab reținuți de membrane, prin urmare, neutralizarea preliminară a acizilor volatili crește selectivitatea procesului de separare.[ ...]
La concentrații mari de sare într-un electrodializator cu trei camere cu membrane inerte, eficiența maximă a curentului nu depășește 20%.[ ...]
Rezultate pozitive de curățare obținute Ape uzate de la OP-7 prin osmoză inversă la o presiune de 5 MPa. Permeabilitatea membranei a fost de 5-20,8 l/(m2-h) la o concentrație de OP-7 în filtrat de 1-18 mg/l.[ ...]
Surfactanții (alchil sulfati) stimulează în cea mai mare măsură reproducerea bacteriilor. În plus, agenții tensioactivi, prin modificarea permeabilității membranelor celulelor vii (S. S. Stroev, 1965 etc.), pot contribui la o mai bună digestibilitate a nutrienților conținuti în apă de către microbi.[ ...]
Natura substanței dizolvate are un anumit efect asupra selectivității și, într-o măsură mai mică, asupra permeabilității membranei. Această influență constă în faptul că substanțele anorganice sunt reținute de membrane mai bine decât substanțele organice cu aceeași greutate moleculară; dintre compușii înrudiți, de exemplu, omologii, substanțele cu o greutate moleculară mai mare sunt reținute mai bine; substanțele care formează legături cu membrana, de exemplu, hidrogenul, sunt reținute de membrană, cu atât mai bine, cu atât această legătură este mai puțin puternică; selectivitatea reținerii compușilor macromoleculari prin ultrafiltrare este cu atât mai mare, cu atât greutatea moleculară a substanței dizolvate este mai mare.[ ...]
Membranele din acetat de celuloză pot funcționa în intervalul de pH 4,5-7, iar cele din polimeri rezistenți chimic pot funcționa la pH 1-14. Permeabilitatea membranelor este aleasă pentru a permite trecerea apei, a sărurilor solubile și a reține uleiurile. Dimensiunea porilor în membrane este de obicei în intervalul 2,5-10 nm. Instalația este dotată cu conducte auxiliare pentru spălarea membranelor cu apă filtrată sau demineralizată, echipată cu instrumente și dispozitive automate.[ ...]
Cu o scădere semnificativă a diferenței de potențial intracelular până la un anumit nivel de prag, se observă o schimbare bruscă a permeabilității membranei și inversarea (reversia) fluxurilor ionice. Ionii de calciu din mediul extern din jurul celulei intră în ea, în timp ce ionii de clorură și ionii de potasiu părăsesc celula în soluția de baie.[ ...]
toleranta este asociata cu factori interniși include procese metabolice precum captarea selectivă a ionilor, permeabilitatea redusă a membranei, imobilizarea ionilor în anumite părți ale plantelor, îndepărtarea ionilor din procesele metabolice prin formarea unei rezerve în forme insolubile în diferite organe, adaptarea la înlocuirea unui organism fiziologic. element cu unul toxic în enzimă, îndepărtarea ionilor din plante atunci când sunt spălate prin frunze, excreția de suc, căderea frunzelor, excreția prin rădăcini. Plantele tolerante pot fi stimulate la concentrații ridicate de metale, ceea ce indică faptul că acestea nevoie fiziologicăîn exces. Unele specii de plante sunt capabile să acumuleze o cantitate semnificativă de metale grele fără semne vizibile de oprimare. Alte plante nu au această capacitate (vezi tabel[ ...]
Presiunea este unul dintre principalii factori care determină performanța instalațiilor de osmoză inversă. Performanța membranelor crește odată cu creșterea presiunii în exces. Totuși, pornind de la o anumită presiune, permeabilitatea membranelor scade din cauza compactării materialului polimeric al membranei.[ ...]
De asemenea, s-a stabilit că scăzut ([ ...]
Deoarece polizaharidele hemicelulozice au o greutate moleculară medie numerică de cel mult 30.000, utilizarea osmometriei convenționale este dificilă din cauza permeabilității membranelor pentru fracțiile cu greutate moleculară mică. Metoda lui Hill de osmometrie în fază de vapori are o serie de avantaje față de alte metode. Această metodă se bazează pe măsurarea diferenței dintre presiunea de vapori a unei soluții și a unui solvent și este după cum urmează. O picătură de soluție și o picătură de solvent sunt plasate pe două joncțiuni de termocuplu și păstrate într-o atmosferă saturată cu vapori puri de solvent. Datorită presiunii reduse de vapori a soluției, o parte din vapori se va condensa pe picătura de soluție, ridicând temperatura picăturii și a termocuplului. Forța electromotoare rezultată este măsurată cu un galvanometru. Limita superioară a valorii măsurate a greutății moleculare este de aproximativ 20.000, precizia măsurării este de 1%.[ ...]
În cele din urmă, membranele reticulului endoplasmatic sunt suprafețele de-a lungul cărora se propagă biocurenții, care sunt semnale care modifică permeabilitatea selectivă a membranelor și, prin urmare, activitatea enzimelor. Datorită acestui fapt, unele reacții chimice sunt puse în mișcare, altele sunt inhibate - metabolismul este supus reglementării și decurge într-o manieră coordonată.[ ...]
Plasmalema reglează intrarea substanțelor în celulă și ieșirea lor din aceasta, asigură pătrunderea selectivă a substanțelor în și din celulă. Viteza de penetrare prin membrana a diferitelor substanțe este diferită. Apa și substanțele gazoase pătrund bine prin ea. Si substantele liposolubile patrund usor, probabil datorita faptului ca are un strat lipidic. Se presupune că stratul lipidic al membranei este pătruns cu pori. Acest lucru permite substanțelor care sunt insolubile în grăsimi să treacă prin membrană. Porii poartă o sarcină electrică, astfel încât pătrunderea ionilor prin ei nu este complet liberă. În anumite condiții, încărcarea porilor se modifică, iar aceasta reglează permeabilitatea membranelor pentru ioni. Cu toate acestea, membrana nu este la fel de permeabilă pentru diferiți ioni cu aceeași sarcină și pentru diferite molecule neîncărcate de dimensiuni similare. Aceasta arată cea mai importantă proprietate a membranei - selectivitatea permeabilității sale: pentru unele molecule și ioni, este mai bine permeabilă, pentru altele mai rău.[ ...]
În prezent, mecanismul de acțiune al mediatorilor în celulele animale și vegetale, care se bazează pe reglarea fluxurilor de ioni, este în general recunoscut. Modificările potențialelor de membrană se datorează schimbărilor în permeabilitatea ionică a membranelor prin deschiderea sau închiderea canalelor ionice. Acest fenomen este asociat cu mecanismele de apariție și propagare a AP în celulele animale și vegetale. În celulele animale, acestea sunt canale N7K+ controlate de canalele de acetilcolină și Ca2+, mai des dependente de aminele biogene. În celulele vegetale, apariția și răspândirea AP este asociată cu canalele de calciu, potasiu și clorură.[ ...]
Cu reproductibilitate și stabilitate mai mari, un flux stabil de gaze și vapori poate fi obținut prin metode bazate pe difuzia gazelor sau vaporilor lichizi printr-un capilar (Fig. 10) sau o membrană permeabilă (Fig. 11) în fluxul de gaz diluant. În astfel de metode se observă un echilibru între faza gazoasă și suprafețele adsorbante ale echipamentului, ceea ce asigură stabilitatea microfluxului.[ ...]
O creștere a temperaturii duce la scăderea vâscozității și a densității soluției și, simultan, la creșterea presiunii osmotice a acesteia. Reducerea vâscozității și a densității soluției crește permeabilitatea membranelor, iar o creștere a presiunii osmotice reduce forța motrice a procesului și reduce permeabilitatea.[ ...]
În orice sistem viu, există un REB și ar fi surprinzător dacă nu ar fi. Aceasta ar însemna egalitatea absolută a concentrațiilor de electroliți în toate celulele, organele, soluțiile externe sau coincidența completă a permeabilității membranei la toți cationii și anionii.[ ...]
În experimentul 6, similar cu experimentul 1, cantitatea de potasiu eliberată și materie organică solubilă în apă a fost determinată la diferite concentrații de atrazină. Judecând după rezultatele obținute, se poate spune că atrazina nu crește permeabilitatea membranelor pentru substanțele organice cu greutate moleculară mică și crește pentru potasiu. Acest efect a fost proporțional cu concentrația de atrazină.[ ...]
La examinarea persoanelor expuse la radiații de nivel scăzut în timpul muncii (de exemplu, radiologi și tehnicieni care lucrează cu raze X, ale căror doze au fost măsurate cu dozimetre individuale) folosind metoda atomilor marcați, au fost efectuate teste de sânge pentru permeabilitatea eritrocitelor. membrane în timpul trecerii cationilor monovalenți. S-a constatat că permeabilitatea membranelor eritrocitare la indivizii iradiați este semnificativ mai mare decât la cei care nu au fost iradiați. În plus, graficul de dependență a făcut posibilă stabilirea unei creșteri rapide a permeabilității la iradiere scăzută; la doze mari, curba devine plată, similar cu observația lui Stokke în studiile pe animale (vezi Fig. XIV-3). Aceste date sunt în concordanță cu rezultatele obținute de Petkau.[ ...]
La desalinizarea apelor uzate saline prin hiperfiltrare prin membrane semipermeabile, principalii parametri - concentrația de substanțe dizolvate în concentrat și filtrat trebuie să fie determinate pe unitatea de lățime a membranei la o lungime dată, capacitatea de separare, coeficientul de permeabilitate a membranei, presiunea, debitele de apă sursă, filtrat și concentrat.[ .. .]
Posibilitatea unei astfel de adaptări se datorează dependenței constantelor termodinamice, chimice și cinetice de temperatură. Această dependență, în general, determină direcția și viteza reacții chimice, tranziții conformaționale ale maodomoleculelor biologice, tranziții de fază ale lipidelor, modificări ale permeabilității membranei și alte procese, a căror funcționare asigură activitatea vitală a organismelor la temperaturi ridicate.[ ...]
Toate acestea sunt doar primii pași în domeniul aplicării apei magnetice în medicină. Cu toate acestea, informațiile deja disponibile indică perspectivele de utilizare a magnetizării sistemelor de apă în acest domeniu. O serie de manifestări medicale sunt posibil (ipotetic) legate de faptul că magnetizarea sistemelor de apă crește permeabilitatea membranelor.[ ...]
S-a stabilit că filmele polimerice produse de industrie pentru ultrafiltrare, schimb ionic, precum și membranele din colodion, gelatină, celuloză și alte materiale, au o selectivitate bună, dar o permeabilitate scăzută (0,4 l/m h la o presiune de 40 am). ). Membranele preparate conform unei prescripții speciale dintr-un amestec de acetat de celuloză, acetonă, apă, perclorat de magneziu și acid clorhidric (respectiv 22,2; 66,7; 10,0; 1,1 și 0,1 procente în greutate) fac posibilă desalinizarea apei de la 5, 25 la 0,05% NaCl si au o permeabilitate de 8,5-18,7 l!m2 ■ h la o presiune de functionare de 100-140 am, durata lor de viata este de cel putin 6 luni. Studii microscopice electronice ale acestor membrane, deoarece, conform calculelor preliminare 1192], osmoza inversă poate deveni competitivă cu alte metode de desalinizare a apei cu o creștere a permeabilității membranei până la 5 m31 mg pe zi.[ ...]
Potențialul de repaus al peretelui celular. Peretele celular (înveliș) are o sarcină de suprafață negativă. Prezența acestei sarcini conferă peretelui celular proprietăți distincte de schimb de cationi. Peretele celular se caracterizează prin selectivitate predominantă pentru ionii de Ca2+, care joacă un rol important în reglarea permeabilității membranei în raport cu ionii K și Na+.[ ...]
Astfel, efectele observate indică faptul că fluidul de cultură al micromicetului Fusarium oxysporum conține, pe lângă acidul fusaric, și alte componente cu activitate biologică ridicată. Gradul de patogenitate a diferitelor izolate de ciuperci fitopatogeni poate fi evaluat pe baza determinării modificărilor permeabilității membranei celule vegetale la amoniac.[ ...]
Ca urmare, formarea de ATP este redusă sau oprită, ceea ce duce la suprimarea proceselor care depind de energia respirației. Structura și permeabilitatea selectivă a membranelor sunt de asemenea perturbate, ceea ce necesită cheltuirea energiei respiratorii pentru a se menține. Aceste modificări duc la scăderea capacității celulelor de a absorbi și reține apa.[ ...]
Pe de altă parte, stabilizarea structurii spațiale a proteinei și a altor biopolimeri se realizează în mare măsură datorită interacțiunii: biopolimer - apă. Complexul apă-proteină-nucleic este considerat a fi baza funcționării sistemelor vii, deoarece numai în prezența acestor trei componente este posibilă funcționarea normală a membranelor. Permeabilitatea selectivă a membranelor depinde de starea apei. Extrapolând modelul cluster al apei la sistemele biologice, se poate demonstra că atunci când clusterul este distrus în anumite zone ale membranei, se deschide o cale de transport preferenţial. Apa fără structură, de exemplu, împiedică comportamentul protonilor în apropierea membranei, în timp ce protonii se propagă rapid de-a lungul unui cadru structurat.[ ...]
Este descrisă o schemă pentru analiza continuă a gazelor folosind un electrod ion-selectiv, care poate fi utilizată pentru a determina conținutul de NH3, HCl și HP în gaze. În revizuirea activității BNS din SUA, printre alte metode de certificare a gazelor (amestecurilor) de referință, este indicată și metoda de certificare folosind electrozi ion-selectivi pentru gazele NSI și NR. Dintre toate modelele de electrozi ion-selectivi, se utilizează de obicei următoarele: o membrană ion-selectivă separă două soluții - interne și externe (testate). Pentru contactul electric, în soluția internă este plasat un electrod auxiliar, reversibil la ionii soluției interne, a cărui activitate este constantă, drept urmare și potențialul este constant. Pe suprafețele interioare și exterioare ale membranei apare o diferență de potențial, care depinde de diferența de activitate a ionilor din soluțiile externe și interne. Teoria apariției potențialului de membrană este descrisă în lucrare. Practic, apariția potențialului se explică prin permeabilitatea membranelor fie numai pentru cationi (cation-selectiv), fie doar pentru anioni (anion-selectiv).
PERMEABILITATEA ŞI TRANSPORTUL SUBSTANŢELOR ÎN MEMBRANELE BIOLOGICE
Permeabilitatea membranelor biologice
Una dintre cele mai importante funcții ale unei membrane biologice este aceea de a asigura schimbul de ioni și molecule între celulă și mediu. Capacitatea membranelor biologice de a trece diferite substanțe prin ea însăși se numește permeabilitate. Studiul permeabilității celulare este de mare importanță, deoarece aproape toate procesele activității vitale celulare sunt asociate cu această funcție: metabolismul, generarea și conducerea biopotențialelor, secreția, recepția etc.
Transport de substante
Orice celulă își menține foarte precis substanța chimică, inclusiv compoziția ionică. Constanța compoziției ionice se numește homeostază ionică și se menține prin transportul de substanțe.
Transportul de substanțe în organism poate fi împărțit în trei categorii:
Transport pe distanțe lungi - transferul de substanțe între organe, lungimea lui în plante poate ajunge la câteva zeci de metri;
Transport în apropiere - transfer între celulele vecine;
Transportul pe membrană este transportul de substanțe prin membrană.
Din punct de vedere al costurilor energetice, transportul cu membrane poate fi împărțit în 2 clase cele mai importante: activ și pasiv.
Transport pasiv nu are legătură directă cu consumul de energie chimică; se realizează ca urmare a difuzării substanțelor spre potenţial electrochimic mai mic(unde concentrația de substanțe și sarcina este mai mică). transport activ merge mai departe cu cheltuirea energiei chimice ATP sau transferul de electroni de-a lungul lanțului respirator.
Tipuri de transport pasiv al substanțelor în celule și țesuturi: difuzie, osmoză.
Difuzie- mecanismul principal de transport pasiv al substanțelor, datorită prezenței unui gradient de concentrație. Există mai multe tipuri de difuzie:
1) difuzie simplă, substanța care difuzează se deplasează de-a lungul unui gradient prin membrană fără a forma un complex sau a pătrunde prin canal;
2) difuzie facilitată efectuată cu ajutorul purtătorilor – proteine sau complexe moleculare.
difuzie simplă - acesta este un proces fizic spontan de pătrundere a unei substanțe dintr-o zonă de concentrație mare într-o zonă de concentrație mai mică, ca urmare a mișcării haotice termice (brownie) a moleculelor.
Cu ajutorul difuziei simple, substanțele solubile în lipide sunt transferate prin membrană, în special astfel de substanțe care sunt foarte solubile în lipide și sunt esențiale pentru metabolismul metabolic, cum ar fi oxigenul și dioxid de carbon. Otrăvurile și medicamentele pot pătrunde și în celulă prin faza lipidică a membranelor.
Fundamentarea matematică a procesului de difuzie a fost dată mai întâi de A. Fick. Conform primei legi a lui Fick, fluxul de materie (J) este direct proporțional cu gradientul de concentrație dC/dx:
https://pandia.ru/text/78/170/images/image002_95.gif" width="433" height="56">
Luând în considerare acest lucru, formula pentru curgerea unei substanțe printr-o membrană poate fi scrisă după cum urmează:
https://pandia.ru/text/78/170/images/image004_68.gif" width="128" height="56">
Coeficient de permeabilitate R are dimensiunea cm/s și este o caracteristică cantitativă a capacității unei anumite substanțe de a pătrunde în membrană. Este asemănător
coeficientul de difuzie (D), dar depinde nu numai de natura substanței și de temperatură, ci și de proprietățile membranei.
DIV_ADBLOCK54">
Penetrarea în celulă a glucozei, glicerolului, aminoacizilor și a altor substanțe nu are o dependență liniară de concentrația lor. Mai mult, la anumite concentrații, rata de penetrare a acestora este mult mai mare decât la difuzia simplă. Această caracteristică se explică prin faptul că, în acest caz, nu se observă difuzie simplă, ci facilitată. Substanța difuzează prin membrană singură, dar viteza de difuzie este mult crescută dacă moleculele acestei substanțe formează un complex cu moleculele purtătorului, care este foarte solubil în lipide. Moleculele purtătoare pot fi atât mobile, cât și fixate în membrană - canale.
Difuzia cu participarea purtătorului, ca și difuzia simplă, are loc până când concentrația pe ambele părți ale membranei devine aceeași.
Difuzarea facilitată este așa-numita difuzia schimbului, în care purtătorul formează un compus cu o substanță care difuzează și se mișcă odată cu acesta de la o suprafață a membranei la alta, unde molecula purtătoare este eliberată, o altă moleculă a aceleiași substanțe îi ia locul, iar complexul este transferat înapoi. În timpul lucrului purtătorilor în cazul difuziei schimbătoare, concentrația de substanțe pe ambele părți ale membranei nu se modifică. Existența difuziei schimbătoare a fost dovedită prin metoda atomilor marcați pe eritrocite, mitocondrii etc.
Pătrunderea particulelor dizolvate cu sarcină electrică prin membrana celulară se realizează prin gradient electrochimic, nu în ceea ce privește concentrarea.
Potențial electrochimic- energia care trebuie cheltuită pentru a muta un mol dintr-o substanță dintr-un punct infinit îndepărtat din spațiu către punct dat cu conditii specifice.
Valoarea potențialului electrochimic este descrisă prin formula:
unde: - membru standard;
Activitatea ionului j
Lucrari chimice;
Volum molal parțial;
Lucru mecanic (lucrare pentru extinderea volumului)
sarcina ionică;
Potential electric;
numărul Faraday;
Munca Electrica.
Moleculele unei substanțe se deplasează întotdeauna spre potențialul lor electrochimic inferior.
Potenţialul Nernst
Luați în considerare un sistem simplu format din două compartimente care conțin o soluție de sare (de exemplu, KCl) și separate printr-o membrană care permite trecerea doar a cationului, în cazul nostru K+.

Soluțiile sunt în echilibru, ceea ce înseamnă că potențialele electrochimice ale ionilor din ambele compartimente sunt aceleași. Din punct de vedere matematic, aceasta va fi exprimată prin ecuația:
Această ecuație, ținând cont de faptul că presiunile și volumele molare ale substanței în ambele compartimente sunt aceleași, ne permite să calculăm diferența de potențiale electrice dintre compartimente:

Această ecuație se numește ecuația Nernst. Dacă luăm în considerare valorile constantelor înainte de semnul logaritmului și folosim logaritmi zecimal în loc de logaritmi naturali, atunci această formulă poate fi scrisă ca:
https://pandia.ru/text/78/170/images/image020_2.jpg" width="459" height="268">
Presiunea osmotică a unei soluții depinde de cantitatea de ioni dizolvați și de temperatură. Conform ecuației van't Hoff, presiunea osmotică (π) a unei soluții este direct proporțională cu concentrația (C) a substanței dizolvate și temperatura absolută a soluției (T):
π = iRTC,
unde i este coeficientul izotonic, care depinde de gradul de disociere a electrolitului și arată de câte ori crește numărul de particule dizolvate în timpul disocierii moleculelor; pentru neelectroliți i = l, pentru electroliți eu > 1; R este constanta gazului.
transport activ- mișcarea împotriva gradientului potențialului electrochimic, efectuată cu cheltuirea energiei chimice datorată hidrolizei ATP sau transferului de electroni prin lanțul respirator.
Alocați transportul activ primar și transportul activ secundar.
Orez. - Transferul activ al ionilor în timpul lucrului de transport ATPaze.
Transport se numește activ primar, dacă este realizat de proteine purtătoare (se mai numesc și pompe ionice) , sursa de energie pentru care sunt ATP, pirofosfat sau substraturi oxidate în lanțurile de transport de electroni ale mitocondriilor, cloroplaste.
Un exemplu tipic de transport activ primar este transportul activ al ionilor cu ajutorul ATPazelor.
secundar activtransport (asociat). numit procesul de transfer de ioni prin membrană împotriva gradientului de concentrație a acesteia datorită energiei gradientului electrochimic al altor ioni. Transportul asociat poate fi efectuat în modul simport(ambele ioni sunt transportați prin membrană în aceeași direcție) sau antiporta(ionii sunt transportați în direcții opuse). Transportul activ secundar asigură transferul membranar de monozaharide, zaharoză, aminoacizi, peptide, anioni și un număr de cationi. În acest scop, proteinele purtătoare membranare folosesc cel mai adesea gradientul electrochimic al ionilor de hidrogen creat de diverse pompe H+.

Orez. - Transportul conjugat de ioni și substanțe prin membrane (Δφ - diferența de potențial, ΔрН - diferența de concentrație de protoni)
Între fluxurile pasive și active în celulă există o interacțiune care vizează menținerea constantei compoziției ionice a celulei.
Când energia celulei este blocată (temperatură scăzută, inhibitori, întuneric), intensitatea transportului activ scade, afluxul de ioni din mediul extern scade. În același timp, se observă o creștere a rezistenței electrice a membranei cu un factor de zece. Aceasta indică o scădere a transportului ionilor prin canale pasive, ceea ce previne pierderea ionilor în celulă.
INTRODUCERE
Transportul pe membrană - transportul substanțelor prin membrana celulară în interiorul sau în afara celulei, efectuat folosind diverse mecanisme - difuzie simplă, difuzie facilitată și transport activ.
Cea mai importantă proprietate a unei membrane biologice este capacitatea sa de a trece diferite substanțe în și din celulă. Acest lucru este de mare importanță pentru autoreglare și menținerea unei compoziții constante a celulei. Această funcție a membranei celulare este îndeplinită datorită permeabilității selective, adică. capacitatea de a trece unele substanțe și de a nu trece altele. Cel mai simplu mod de a trece prin stratul dublu lipidic sunt moleculele nepolare cu o greutate moleculară mică (oxigen, azot, benzen). Molecule polare mici precum dioxidul de carbon, oxidul de azot, apa și ureea pătrund rapid prin stratul dublu lipidic. Etanolul și glicerolul, precum și steroizii și hormonii tiroidieni trec prin stratul dublu lipidic cu o viteză vizibilă. Pentru moleculele polare mai mari (glucoză, aminoacizi), precum și pentru ioni, stratul dublu lipidic este practic impermeabil, deoarece partea sa interioară este hidrofobă. Deci, pentru apă, coeficientul de permeabilitate (cm/s) este de aproximativ 10-2, pentru glicerol - 10-5, pentru glucoză - 10-7 și pentru ionii monovalenți - mai puțin de 10-10.
Transportul moleculelor și ionilor polari mari are loc datorită proteinelor canalului sau proteinelor purtătoare. Deci, în membranele celulare există canale pentru ionii de sodiu, potasiu și clor, în membranele multor celule există canale de apă ale acvaporinelor, precum și proteine purtătoare pentru glucoză, diferite grupuri de aminoacizi și mulți ioni. Transport activ și pasiv.
Membranele formează structura celulei și își îndeplinesc funcțiile. Încălcarea funcțiilor membranelor celulare și intracelulare stă la baza leziunilor celulare ireversibile și, ca urmare, dezvoltarea unor boli severe ale sistemului cardiovascular, nervos și endocrin.
1. Date de bază despre structura membranei celulare.
Membranele celulare includ plasmolema, caryolemma, membranele mitocondriale, EPS, aparatul Golgi, lizozomii, peroxizomii. O caracteristică comună a tuturor membranelor celulare este că acestea sunt straturi subțiri (6-10 nm) de natură lipoproteică (lipide în combinație cu proteine). Principal componente chimice membranele celulare sunt lipide (40%) și proteine (60%); in plus, carbohidrati (5-10%) au fost gasiti in multe membrane.
Membrana plasmatică înconjoară fiecare celulă, îi determină dimensiunea și menține diferențele dintre conținutul celulei și mediul extern. Membrana servește ca un filtru foarte selectiv și este responsabilă de transportul activ al substanțelor, adică de intrarea în celulă. nutriențiși eliminarea deșeurilor nocive. În cele din urmă, membrana este responsabilă de percepția semnalelor externe, permițând celulei să răspundă la schimbările externe. Toate membranele biologice sunt ansambluri de molecule de lipide și proteine ținute împreună prin interacțiuni non-covalente.
Baza oricărei membrane moleculare este formată din molecule de lipide care formează un strat dublu. Lipidele includ un grup mare de substanțe organice care au o solubilitate slabă în apă (hidrofobicitate) și o solubilitate bună în solvenți organici și grăsimi (lipofilitate). Compoziția lipidelor din diferite membrane nu este aceeași. De exemplu, membrana plasmatică, spre deosebire de membranele reticulului endoplasmatic și mitocondriilor, este îmbogățită cu colesterol. Reprezentanții caracteristici ai lipidelor găsite în membranele celulare sunt fosfolipidele (glicerofosfatide), sfingomielinele și colesterolul din lipidele steroizi.
O caracteristică a lipidelor este împărțirea moleculelor lor în două părți funcțional diferite: hidrofobe nepolare, nepurtătoare de sarcină ("cozi"), constând din acizi grași, și "capete" polare hidrofile, încărcate. Aceasta determină capacitatea lipidelor de a forma în mod spontan structuri membranare cu două straturi (bilipide) cu o grosime de 5-7 nm.
Primele experimente care au confirmat acest lucru au fost efectuate în 1925.
Formarea bistratului este o proprietate specială a moleculelor de lipide și se realizează chiar și în afara celulei. Cele mai importante proprietăți ale stratului dublu: capacitatea de auto-asamblare - fluiditate - asimetrie.
2. Idei generale despre permeabilitate.
Caracteristicile membranelor, pereților vaselor și celulelor epiteliale, reflectând capacitatea de a conduce substanțe chimice; distinge între activ (transport activ de substanţe) şi pasiv P. (fagocitoză
3. Transferul de molecule prin membrană.
Deoarece interiorul stratului lipidic este hidrofob, acesta oferă o barieră practic impenetrabilă pentru majoritatea moleculelor polare. Datorită prezenței acestei bariere, scurgerea conținutului celulelor este împiedicată, totuși, din această cauză, celula a fost nevoită să creeze mecanisme speciale pentru transportul substanțelor solubile în apă prin membrană. Transferul moleculelor mici solubile în apă se realizează folosind proteine de transport speciale. Acestea sunt proteine transmembranare speciale, fiecare dintre acestea fiind responsabilă de transportul anumitor molecule sau grupuri de molecule înrudite.
În celule, există și mecanisme pentru transferul de macromolecule (proteine) și chiar de particule mari prin membrană. Procesul de absorbție a macromoleculelor de către celulă se numește endocitoză. În termeni generali, mecanismul apariției sale este următorul: zonele locale ale membranei plasmatice se invaginează și se închid, formând o veziculă endocitară, apoi particula absorbită intră de obicei în lizozomi și suferă degradare.
3.1 Difuzie (latina difuzio - distribuție, răspândire, împrăștiere) - procesul de transfer de materie sau energie dintr-o zonă de concentrație mare într-o zonă de concentrație scăzută (împotriva gradientului de concentrație). Cel mai faimos exemplu de difuzie este amestecarea gazelor sau a lichidelor (dacă aruncați cerneală în apă, lichidul va deveni uniform colorat după un timp). Un alt exemplu este legat de un solid: dacă un capăt al tijei este încălzit sau încărcat electric, căldura (sau, respectiv, curentul electric) se răspândește din partea fierbinte (încărcată) în partea rece (neîncărcată). În cazul unei tije metalice, difuzia termică se dezvoltă rapid, iar curentul curge aproape instantaneu. Dacă tija este făcută din material sintetic, difuzia termică are loc lent, iar difuzia particulelor încărcate electric este foarte lentă. Difuzia moleculelor se desfășoară în general și mai lent. De exemplu, dacă o bucată de zahăr este coborâtă pe fundul unui pahar cu apă și apa nu este amestecată, va dura câteva săptămâni până când soluția devine omogenă. Și mai lentă este difuzarea unui solid în altul. De exemplu, dacă cuprul este acoperit cu aur, atunci aurul se va difuza în cupru, dar în condiții normale ( temperatura camereiși presiunea atmosferică) stratul purtător de aur va atinge o grosime de câțiva micrometri abia după câteva mii de ani.
Toate tipurile de difuzie se supun acelorași legi. Rata de difuzie este proporțională cu aria secțiunii transversale a probei, precum și cu diferența de concentrații, temperaturi sau încărcături (în cazul valorilor relativ mici ale acestor parametri). Astfel, căldura va călători de patru ori mai repede printr-o tijă de doi centimetri în diametru decât printr-o tijă de un centimetru în diametru. Această căldură se va răspândi mai repede dacă diferența de temperatură pe centimetru este de 10°C în loc de 5°C. Rata de difuzie este, de asemenea, proporțională cu parametrul care caracterizează un anumit material. În cazul difuziei termice, acest parametru se numește conductivitate termică, în cazul unui flux de sarcini electrice - conductivitate electrică. Cantitatea de substanță care difuzează într-un timp dat și distanța parcursă de substanța care difuzează sunt proporționale cu rădăcina pătrată a timpului de difuzie.
Difuzia este un proces la nivel molecular și este determinat de natura aleatorie a mișcării moleculelor individuale. Viteza de difuzie este deci proporțională cu viteza medie a moleculelor. În cazul gazelor viteza medie există mai multe molecule mici, și anume, este invers proporțional cu rădăcina pătrată a masei moleculei și crește odată cu creșterea temperaturii. Procesele de difuzie în solide la temperaturi ridicate sunt adesea întâlnite uz practic. De exemplu, anumite tipuri de tuburi catodice (CRT) folosesc toriu metalic difuzat prin wolfram metalic la 2000°C.
3.2 Ecuația lui Fick
În cele mai multe cazuri practice, în locul potențialului chimic se folosește concentrația C. Înlocuirea directă a lui µ cu C devine incorectă în cazul concentrațiilor mari, deoarece potențialul chimic este legat de concentrație conform unei legi logaritmice. Dacă nu luăm în considerare astfel de cazuri, atunci formula de mai sus poate fi înlocuită cu următoarea.