biosinteza peretelui celular. Celuloza în nutriție Biosinteza celulozei în plante
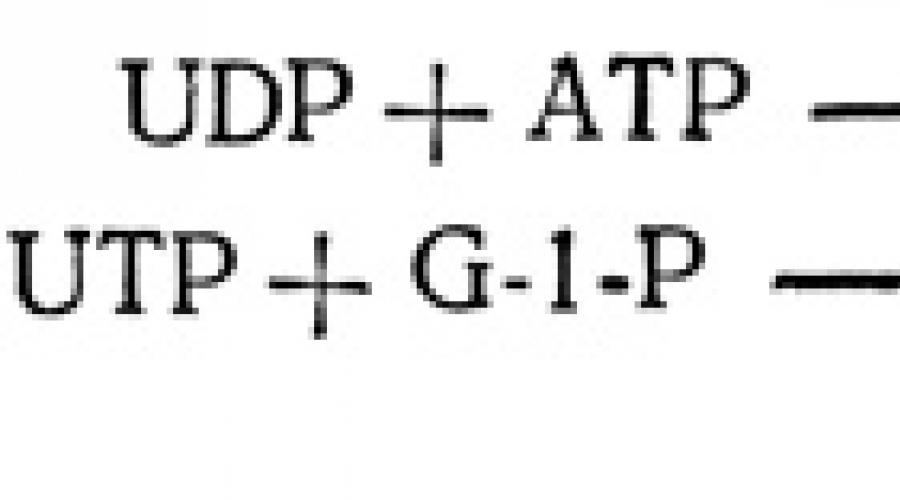
Citeste si
În ciclul Calvin-Benson, după cum sa menționat mai sus, se formează fructoză-6-fosfat (F-6-P). Acest hexoză fosfat poate fi transformat, sub acțiunea unor enzime specifice, în alte hexoze fosforilate și anume glucoză-6-fosfat (G-6-P) și glucoză-1-fosfat (G-1-P). Transformarea inversă se produce, de asemenea, cu ușurință.
Acești trei hexoză fosfați sunt apoi utilizați pentru a construi lanțuri de molecule de carbohidrați utilizate pentru transport, depozitare și în reacții de sinteză. Pentru ca astfel de transformări să apară, hexozofosfații trebuie mai întâi activați. Acest lucru se realizează de obicei prin atașarea lor la nucleotide, structuri inelare complexe similare cu acidul adenilic ATP. Produsul unei astfel de reacții de adiție este derivați de nucleotide ai monozaharidelor sau zaharuri nucleotidice. Cea mai frecventă este uridin difosfoglucoza (UDPG), formată în reacția dintre uridin trifosfat (UTP) și glucoză-1-fosfat (G-1-P). UTP în sine se formează indirect, ca urmare a transferului unei grupări fosfat de la ATP la UDP (uridin difosfat).

Nucleotidele ATP și UTP sunt prezente în toate celulele deoarece sunt utilizate împreună cu alte nucleotide în sinteza ADN și ARN.
Zaharurile sunt transportate în întreaga plantă sub formă de zaharoză, o dizaharidă formată din reziduuri de glucoză și fructoză (Fig. 5.2). Zaharoza se formează în reacția dintre UDPG și F-6-P:


Echilibrul acestei reacții este puternic deplasat spre sinteza zaharozei, ceea ce asigură posibilitatea acumulării acestei dizaharide în concentrații semnificative. Pentru utilizarea ulterioară, zaharoza trebuie mai întâi să sufere divizarea: enzima invertaza își catalizează hidroliza cu formarea de glucoză și fructoză liberă.

Energia legăturii glicozidice într-o astfel de reacție este irosită, fiind distribuită între două molecule. Prin urmare, dacă glucoza și fructoza urmează să fie descompuse în timpul respirației sau să participe (ca materii prime) la sinteza polizaharidelor, atunci ele trebuie mai întâi să sufere din nou fosforilarea datorită ATP. Procesele de sinteză și de descompunere a zaharozei arată în mod clar că reacțiile anabolice și catabolice (reacții de sinteză și de descompunere) urmează adesea căi diferite.
Sinteza amidonului și celulozei
Lanțurile polimerice lungi de amidon și celuloză sunt construite din aceleași unități elementare - reziduuri de glucoză, conectate doar în moduri diferite. Această diferență structurală este responsabilă pentru faptul că cei doi considerați polimeri ai glucozei (glucanii) sunt semnificativ diferiți ca natură; amidonul, de exemplu, este ușor de digerat în corpul uman „și celuloza nu este deloc digerată. Principala lor diferență este că atomii de carbon 1 și 4 ai două resturi de glucoză adiacente sunt legați în amidon prin legături α și în celuloză ( Legături β (Fig. 5.3). Amidonul este reprezentat de două forme: un polimer liniar, sau amiloză, care nu conține alte legături cu excepția α-1,4-glicozidice și un polimer ramificat, sau amilopectină, în care, împreună cu α Legăturile -1,4 -glicozidice sunt, de asemenea, legături 1,6. Diferența de natură a legăturilor determină aranjarea spațială inegală a lanțurilor polimerice. Amidonul este principala polizaharidă de rezervă a plantei. Este insolubil în apă și se depune strat cu strat în boabele de amidon conținute în cloroplaste (vezi Fig. .2.20) sau în leucoplaste lipsite de clorofilă a țesuturilor de depozitare ale tulpinii, rădăcinilor și semințelor. Uneori celulele țesutului de depozitare sunt literalmente înfundate cu boabe de amidon, care sunt ușor de identificat în ele, deoarece pot fi colorate în albastru cu iod. Fiind insolubil în apă, amidonul, spre deosebire de zaharoză și hexoze, nu provoacă un efect osmotic în celule (vezi cap. 6). Prin urmare, formarea amidonului în celulele frunzelor în perioadele de fotosinteză intensă împiedică suprimarea acesteia din urmă, care are loc ca urmare a acumulării de produse de fotosinteză. În întuneric, amidonul este hidrolizat treptat din nou pentru a forma glucoză fosfat, care este apoi transformat în zaharoză, care este transportată în alte părți ale plantei.

Orez. 5.3. Structura amidonului (A) și celulozei (B) (Modificat de la J. Bonner, A. W. Galston. 1952. Principles of Plant Physiology, San Francisco, W. H. Freeman and Co.) Rețineți că formulele chimice ale amidonului și celulozei sunt aceleași, dar se deosebesc aceste polizaharide prin orientarea spațială a punților lor de oxigen. A. Amidonul, principala polizaharidă de depozitare a plantei, este construit din două componente distincte: amiloză, cu lanțurile sale lungi neramificate de unități de glucoză, și amilopectina, constând din un numar mare lanțuri scurte ramificate. B. Celuloza, componenta principală a peretelui celular primar, există sub formă de lanțuri polimerice lungi. Lanțurile sunt combinate în fire micelare, iar acestea din urmă în microfibrile. Microfibrilele suficient de mari pentru a fi văzute cu un microscop electronic alcătuiesc urzeala și bătătura peretelui celular.
Produsul de pornire pentru sinteza amidonului este adenozin difosfoglucoza (ADPG), format din ATP și G-1-P:

Molecula de amidon este construită prin adăugarea treptată a unui reziduu de glucoză după altul într-o reacție ADPG cu un lanț de glucoză preformat:
Cu un conținut scăzut de zaharoză, amidonul este descompus și. transformat în zaharoză. Cu toate acestea, mai întâi este scindat la reziduuri de glucoză și la fiecare dintre ele se adaugă un reziduu de acid fosforic, adică se formează G-1-P, ceea ce asigură conservarea energiei de legătură:
Acest G-1-P poate fi apoi utilizat pentru sinteza zaharozei, pe care am descris-o mai sus. În semințe și în alte organe, în care există o divizare simultană a unor cantități mari de amidon, se descompune în maltoză dizaharidă (G-G) sub acțiunea a-amilazei. Maltoza se descompune apoi în glucoză, din care (pentru transport) zaharoza este din nou sintetizată. Această a doua cale, spre deosebire de prima, nu conservă energia de legare, așa că aici este necesar ATP pentru a converti glucoza în glucoză-6-P.

Celuloza, cel mai abundent carbohidrat de pe Pământ, este componenta principală a peretelui celular primar. Moleculele sale sunt construite în același mod ca și moleculele de amidon, cu diferența că un alt derivat nucleotidic al monozaharidei, guanozin difosfoglucoza (GDPG), joacă rolul unui donor de glucoză și că legătura dintre unitățile monomerice nu aparține α-, dar de tip β.

În unele cazuri, UDPG poate fi și un donor de glucoză pentru sinteza celulozei.
În corpul plantelor superioare, celuloza este rareori descompusă (cu excepția defalcării cauzate de activitatea microbilor). Două excepții notabile de la această regulă se referă la celulele din zona de separare a frunzei, care se formează înainte de a vărsa frunzele, și la vasele de xilem, în care pereții transversali se dizolvă. În zona de separare a frunzei, enzima celulază distruge pereții celulari, împărțind celuloza conținută în ei în unități monomerice individuale, adică în glucoză. Pereții celulari slăbiți de acest proces se rupe în cele din urmă și frunza se năruie.
Microfibrilele de celuloză din peretele celular sunt ținute împreună printr-o matrice de lanțuri polizaharide mixte, în principal xiloglucani și arabinogalactani (vezi Figura 2.31). (Xiloza și arabinoza sunt zaharuri cu cinci atomi de carbon (pentoze), iar galactoza este o hexoză înrudită cu glucoza.) Aceste polizaharide sunt sintetizate și din precursori, zaharuri nucleotidice, în principal în dictiozomi. Veziculele desprinse din dictiozomi se contopesc în cele din urmă cu plasmalema și în acest fel își transferă conținutul în peretele celular emergent.
Deci, toate polizaharidele trec cu ușurință una în alta, dar sinteza lor trece întotdeauna prin stadiul zaharurilor nucleotidice, în timp ce descompunerea are loc într-un mod mai direct.
În majoritatea produselor origine vegetală conține așa-numitele substanțe de balast care nu sunt digerate în tractul gastrointestinal - fibre și pectină. Cu toate acestea, ele sunt necesare pentru o persoană. Dacă hrana este săracă în ele, apar atonia intestinală și constipația. Astfel, fibrele sunt un reglator al motilității intestinale.
Fibrele îmbunătățesc funcția intestinală, încetinesc procesele de putrefacție și formarea gazelor și reduc absorbția anumitor substanțe nocive. Deci, de exemplu, pentru prevenirea bolilor profesionale la lucrătorii cu săruri metalice și substanțe radioactive, se recomandă consumul de coacăze roșii, care conține multe pectine.
Polizaharidă: Fibră
Fibre (celuloză)- o polizaharidă care formează cea mai mare parte a pereților celulari ai plantelor. Fibrele sunt insolubile în apă, doar se umflă în ea. Fibrele reprezintă mai mult de 50% din lemn. În fibrele de bumbac, este mai mare de 90%. Când sunt fierte cu acid sulfuric puternic, fibrele sunt complet transformate în glucoză. Cu hidroliză mai slabă, celobioza se obține din celuloză.
Într-o moleculă de celuloză, reziduurile de celobioză sunt legate prin legături glicozidice sub forma unui lanț lung. Greutatea moleculară a fibrei nu a fost determinată cu precizie. Se crede că fibra nu este o substanță individuală, ci este un amestec de substanțe omoloage. Greutățile moleculare ale fibrelor derivate din diverse surse, fluctuează destul de puternic: bumbac - 330.000 (2020 reziduuri glicozidice în lanț); ramie - 430.000 (2660 reziduuri), lemn de molid - 220.000 (1360 reziduuri). Folosind analiza de difracție cu raze X, s-a constatat că moleculele de fibre au o formă filamentoasă. Aceste molecule filamentoase sunt conectate în mănunchiuri - micele. Fiecare micelă constă din aproximativ 40-60 de molecule de fibre.
Conectarea moleculelor individuale de fibre în micelii are loc datorită legăturilor de hidrogen, care se realizează atât datorită atomilor de hidrogen ai grupărilor hidroxil ale fibrei, cât și datorită moleculelor de apă adsorbite de fibră. În pereții celulelor vegetale, miceliile din fibre sunt legate de hidrogen de diferite heteropolizaharide. De exemplu, la arțarul alb, aceștia sunt xiloglucan, interconectați prin legături glicozidice, constând din reziduuri de glucoză, xiloză, galactoză și fucoză; arabinogalactan, construit din reziduurile de arabinoză și galactoză; ramnogalacturonan, format din reziduuri de acid galacturonic și ramnoză. În plus, există dovezi că construcția peretelui celular al plantei, în special în stadiile incipiente ale formării sale, implică și o glicoproteină specială bogată în hidroxiprolină extensină. Când pereții celulari devin lignificați, lignina se acumulează în ei.
Fibrele nu sunt digerate în tractul gastrointestinal uman. Este digerat doar de rumegătoare, în stomacul cărora se află bacterii speciale care hidrolizează fibrele cu ajutorul enzimei celulaze secretate de acestea.
Hemiceluloze (semifibre). Sub această denumire, ele combină un grup mare de polizaharide cu molecul mare care sunt insolubile în apă, dar solubile în soluții alcaline. Hemicelulozele se gasesc in cantitati semnificative in partile lignificate ale plantelor: paie, seminte, nuci, lemn, stiuleti de porumb. O cantitate mare de hemiceluloze se găsește în tărâțe. Hemicelulozele sunt hidrolizate de acizi mai ușor decât celuloza. În același timp, ele formează manoză, galactoză, arabinoză sau xiloză și de aceea, respectiv, se numesc manani, galactani și pentozani (araban sau xilan).
La plante, în procesul de fotosinteză, se formează nu numai esteri fosfatici ai zaharurilor sau zaharuri simple, ci și forme mai complexe de carbohidrați - zaharoză, amidon, fibre. Defalcarea formelor complexe de carbohidrați în forme simple decurge, de asemenea, foarte rapid. Acest lucru se observă, de exemplu, în timpul germinării semințelor, îmbătrânirii organelor vegetative etc. Zaharurile simple sau esterii lor de fosfor formați în timpul dezintegrarii curg în organele de reproducere, unde carbohidrații complecși sunt din nou sintetizați din ele. Și, în sfârșit, în plante, procesele de transformări reciproce ale carbohidraților decurg foarte ușor.
Interconversia monozaharidelor trece prin esterii fosforici ai zaharurilor sau prin derivații lor de uridin difosfat (derivați UDP). Derivații UDP ai zaharurilor sunt unul sau altul zahăr conectat prin două resturi de acid fosforic cu uridină, de exemplu:
Fig.1. Uridin difosfat glucoză
Sinteza zaharozei
Zaharoza este cea mai importantă ligozaharidă produsă de plante, sub forma căreia carbonul legat și energia sunt transportate în întreaga plantă. Constă dintr-un reziduu de α-D-glucoză în forma sa de piranoză legat printr-o legătură glicozidică de β-D-fructoză în forma sa de furanoză. Deoarece atomul de carbon anomeric al ambelor monozaharide este implicat în formarea unei legături glicozidice, grupările lor hemiacetale sunt blocate și niciun inel nu se poate deschide. Astfel, zaharoza este un zahăr nereducător (nu reduce reactivii Fehling și Benedict) și, cu excepția sensibilității sale extreme la hidroliza acidă, este inert din punct de vedere chimic. Când este încălzită cu acizi, sau sub acțiunea zaharazei (invertază), zaharoza este hidrolizată, formând zahăr invertit - un amestec de glucoză și fructoză. Zaharoza este foarte solubilă în apă și are un gust dulce.
Zaharoza este folosită ca produs alimentar, precum și în producția de surfactanți (esteri de zaharoză cu acizi mai mari). Principala sursă de zaharoză este sfecla de zahăr, care conține până la 23% zaharoză, și trestia de zahăr, ale cărei tulpini conțin 10-18% zaharoză. Acum s-a stabilit că zaharoza este sintetizată nu numai în cloroplaste, ci și în citoplasma celulelor fotosintetice din UDP-glucoză și fructoză-6-fosfat, care au apărut din
dihidroxiacetonă fosfat. Această substanță se formează în timpul fotosintezei în cloroplaste și apoi intră în citoplasmă. În țesuturile nefotosintetice (de exemplu, în endospermul boabelor de ricin germinate), formarea zaharozei din UDP-glucoză și fructoză-6-fosfat are loc și în citoplasma celulară.
Zaharoza (trestie de zahăr, zahăr din sfeclă) este cea mai comună dizaharidă în natură. În plante, se formează din glucoză și fructoză. Primul pas este fosforilarea glucozei:
Glucoză + ATP → glucoză-6-fosfat + ADP,
apoi se izolează glucoza-6-fosfat la glucoză-1-fosfat. Glucoza-1-fosfat se combină cu UTP, în urma căreia acidul pirofosfat este scindat și se formează un compus de glucoză cu acid uridin difosforic (UDP) - glucoză uridin difosfat.
În același timp, fructoza este fosforilată sub acțiunea enzimei fructokinaze cu participarea ATP:
fructoză + ATP → fructoză-6-fosfat + ADP.
După aceea, interacțiunea UDP-glucoză cu fructoză-6-fosfat are loc cu participarea enzimei zaharoză-fosfat-UDP-glucoziltransferaza. În cele din urmă, zaharoza-6-fosfatul rezultat este hidrolizat prin acțiunea enzimei fosfatazei pentru a forma zaharoză liberă. Astfel, pentru biosinteza unei molecule de zaharoză sunt necesare 3 legături fosfat de înaltă energie, această reacție este ireversibilă. În țesuturile nefotosintetice ale unor plante, de exemplu, în rădăcinile de sfeclă de zahăr, tuberculi de cartofi și altele, zaharoza poate fi formată din fructoză liberă:
UDP-glucoza + fructoza ↔ zaharoză + UDP.
Reacția este catalizată de enzima zaharoză-UDP-glucoziltransferază și, în funcție de condiții, poate fi direcționată atât spre sinteza, cât și spre descompunerea zaharozei.
Sinteza amidonului
Amidonul este o polizaharidă de rezervă a plantelor și poate fi păstrat în plantă pentru o perioadă lungă de timp, sau consumat suficient de repede. Pe perioadă lungă de timp este depozitat în multe semințe, tuberculi și rizomi și se folosește numai atunci când aceste organe germinează. Pentru o perioadă scurtă de timp, amidonul se formează în cloroplaste în timpul perioadei de fotosinteză rapidă, iar în perioada de întuneric ulterioară, este consumat și curge din frunze sub formă de zaharoză. Amidonul se formează și se depozitează întotdeauna sub formă de boabe de amidon situate în plastide - cloroplaste sau amiloplaste. Boabele de amidon sunt structuri foarte organizate, a căror formă și dimensiune sunt foarte diverse, dar adesea caracteristice unei anumite specii de plante. Forma boabelor poate fi sferică, ovoidă, lenticulară sau neregulată; dimensiunea poate varia de la 1 la 100 de microni. Cele mai mari boabe de amidon sunt în cartofi, iar cele mai mici în orez și hrișcă. Boabele de amidon conțin până la 20% apă (din care 10% este legat chimic de amidon) și un număr de straturi concentrice de amidon. Boabele de amidon se formează prin stratificarea unor straturi nou formate pe cele existente anterior.
Conținutul de minerale din amidon este foarte scăzut - 0,2-0,7%, acestea fiind reprezentate în principal de acid fosforic. În amidon s-au găsit unii acizi grași cu molecul mare (palmitic, stearic etc.), al cărui conținut ajunge la 0,6%. Amidonul este un amestec de două polizaharide, amiloză și amilopectină. Moleculele de amiloză sunt lanțuri lungi neramificate care conțin de la 100 la câteva mii de reziduuri de glucopiranoză conectate prin legături glicozidice. Un studiu de difracție cu raze X a arătat că moleculele de amiloză au o structură elicoidală cu diametrul de 1,3 nm cu șase reziduuri succesive de glucoză pe tură a helixului.
Moleculele de amiloză sunt solubile în apă fierbinte, dar soluția rezultată este instabilă și apoi are loc precipitarea spontană a amilozei, cunoscută sub numele de retrogradare. Acest lucru se datorează tendinței moleculelor de amiloză lungi și subțiri de a se alinia unele lângă altele și de a forma agregate insolubile prin legături de hidrogen. Uridin difosfat glucoză-(UDPA) poate servi ca donator de reziduuri de glucoză în biosinteza amilozei. Pentru formarea sa în mediul de reacție este necesară prezența unei sămânțe, care pot fi polizaharide construite din doar 3-4 resturi de glucoză legate prin legături α(1-4).
Reziduurile de glucoză sunt transferate la acceptor (sămânță), unde are loc alungirea lanțului. Reacția se desfășoară conform schemei:
UDFG + acceptor (G)k -- UDP + acceptor (G)k + 1,
unde G sunt reziduuri de glucoză.
Enzima care catalizează această reacție se numește UDPG amidon glucozil transferază.
La majoritatea plantelor, donorul activ de glucoză nu este UDP-glucoza, ci adenozin difosfat glicoza α (ADPG). Reacția de adăugare a resturilor de glucoză de la ADPG la un acceptor cu greutate moleculară mică se desfășoară într-un mod similar și este catalizată de enzima ADPG amidon glucoziltransferază. Sinteza unei molecule de amilopectină ramificată având legături α (1-6) are loc cu ajutorul enzimei α-glucan transferază (enzima Q). D-enzima sau glucoziltransferaza, care formează legături α(1-4) și participă la formarea semințelor, este, de asemenea, implicată în sinteza amidonului.
Descompunerea amidonului are loc cu participarea a două procese - hidroliza și fosforoliza.Descompunerea hidrolitică a amidonului se realizează sub acțiunea a patru enzime din clasa hidrolizei α-amilazei, catalizează scindarea legăturilor α (1-4) și legăturile sunt rupte aleatoriu. Produsul final al acestei degradari este maltoza, glucoza, dextrinele. Sub acțiunea β-amilazei, legăturile α (1-4) sunt scindate cu formarea de resturi de maltoză. Enzima glucoamilaza catalizează scindarea secvenţială a resturilor de glucoză din molecula de amidon. Amilopectin-1,6-glucozidaza sau enzima R catalizează scindarea legăturilor α (1-6) din molecula de amilopectină, adică acționează asupra punctelor de ramificație.
Fosforoliza este adăugarea de acid fosforic la locul de ruptură a legăturii glucozidice dintre resturile de monozaharide din lanțul polizaharidic și se formează glucoză-1-fosfat. Această reacție este catalizată de enzima magglucon fosforilază, care aparține clasei de transferaze. Amidonul din plantă poate fi descompus foarte repede, deoarece enzimele de descompunere se găsesc în toate organele plantei.
Sinteza celulozei
Celuloza este insolubilă în apă, doar se umflă în ea. Cu hidroliza acidă (fierbe în acid sulfuric) se transformă în glucoză, cu una mai slabă - în celobioză. Folosind analiza de difracție cu raze X, s-a constatat că molecula de celuloză are o formă filamentoasă. Aceste molecule filamentoase, datorită legăturilor de hidrogen, sunt conectate în mănunchiuri de 40-60 de bucăți per micelă. În pereții celulelor vegetale, miceliile de celuloză sunt legate prin legături de hidrogen de diferite heteropolizaharide. De exemplu, la arțarul alb sunt xiloglucan, arabinogalactan, ramnogalacturon conectat prin legături glicozidice. În plus, există dovezi că o glicoproteină specială bogată în oxiprolină, extensina, este implicată în construcția peretelui celular al plantei.
Celuloza este construită din reziduuri de β-glucoză. Nu este glucoza liberă care participă la biosinteza celulozei, ci derivatul său GDP - guanozin difosfat glucoză cu participarea enzimei celulozo sintetazei conform schemei:
GDF - glucoză + (glucoză) c → GDF + (glucoză) c + 1
Descompunerea celulozei are loc predominant pe cale hidrolitică sub acțiunea enzimei celulazei până la celobioza dizaharidă.
Carbohidrații sunt transportați sub formă de zaharoză. În procesul de fotosinteză, se formează o mulțime de carbohidrați, și în acest sens mare importanță are un flux de asimilați către alte părți ale celulei din cloroplaste. Pătrunderea hexozelor fosforilate și a zaharozei prin membrana cloroplastică este dificilă; fosfații de trioză (PHA și FDA) pătrund cel mai ușor în membranele cloroplastice. Se presupune că carbohidrații complecși rezultați se descompun în trioză fosfați și, în această formă, se deplasează în citoplasmă, unde pot servi ca material pentru resinteza hexozelor, zaharozei și amidonului.
Transportul parenchimatos intercelular se realizează în două moduri - prin plasmodesmate (simplast) sau prin spațiul liber (apoplast). Zaharoza formată în celulele mezofilei frunzelor este desorbită în apoplast. Lăsând celulele parenchimatoase în apoplast, zaharoza este scindată de invertază în hexoze. Hexozele se deplasează de-a lungul apoplastului către celulele de transfer ale fasciculelor vasculare de-a lungul unui gradient de concentrație. Când sunt în contact cu celulele de transfer ale floemului, acestea se transformă din nou în zaharoză. Apoi, tuburile de sită sunt încărcate, zaharoza curge împotriva gradientului de concentrație și este necesar un consum de energie (ATP).
Se presupune că zaharoza traversează membrana cu ajutorul unui purtător în complex cu un proton. În același timp, datorită activității H + -ATP-azei, ionii H + sunt pompați din celulele floemului și apoi revin de-a lungul gradientului de pH, trăgând zaharoza împreună cu ea împotriva gradientului său de concentrație. Principala formă de transport a carbohidraților în floem este zaharoza (C 12 H 22 O 11). La unele specii, împreună cu zaharoza, oligozaharidele (rafinoza, stahioza), precum și unii alcooli, servesc ca formă de transport a carbohidraților.