Le créateur de la théorie chromosomique de l'hérédité est un scientifique. Théorie chromosomique de l'hérédité
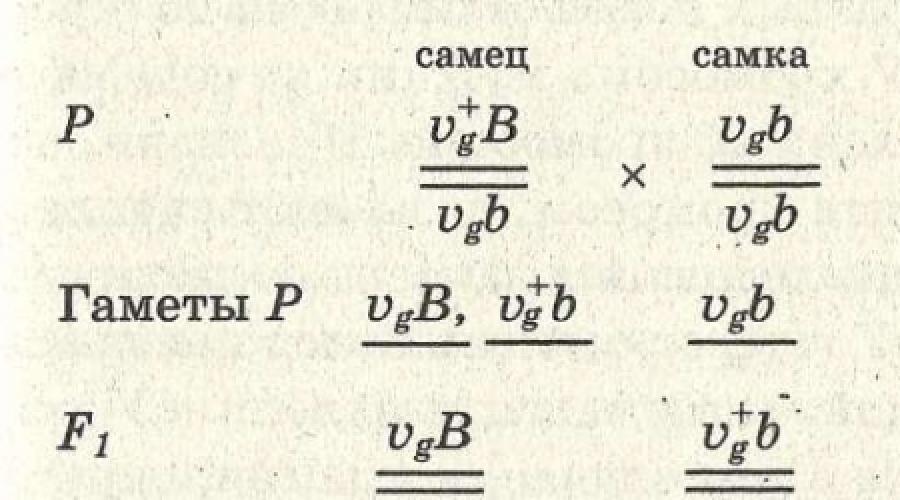
Lire aussi
Les cellules enfermées dans le noyau sont porteuses de gènes et représentent la base matérielle de l'hérédité, c'est-à-dire la continuité des propriétés des organismes dans un certain nombre de générations est déterminée par la continuité de leurs chromosomes. H. t. n. apparu au début du 20e siècle. sur la base de la théorie cellulaire et l'utilisation de l'analyse hybridologique pour étudier les propriétés héréditaires des organismes.
En 1902, W. Setton aux États-Unis, qui a attiré l'attention sur le parallélisme dans le comportement des chromosomes et des soi-disant mendéliens. "facteurs héréditaires", et T. Boveri en Allemagne a avancé l'hypothèse chromosomique de l'hérédité, selon laquelle les facteurs héréditaires mendéliens (appelés plus tard gènes) sont localisés dans les chromosomes. La première confirmation de cette hypothèse a été obtenue dans l'étude du mécanisme génétique de la détermination du sexe chez les animaux, lorsqu'il a été constaté que ce mécanisme est basé sur la distribution des chromosomes sexuels parmi la progéniture. Une justification supplémentaire de H. t. appartient au généticien américain T. H. Morgan, qui a remarqué que le transfert de certains gènes (par exemple, le gène qui cause la drosophile femelle aux yeux blancs lorsqu'il est croisé avec des mâles aux yeux rouges) est associé au transfert du chromosome X sexuel, c'est-à-dire que traits liés au sexe (une personne présente plusieurs dizaines de ces signes, dont certains défauts héréditaires - daltonisme, hémophilie, etc.).
Preuve de H. t. a été obtenu en 1913 par le généticien américain C. Bridges, qui a découvert la non-disjonction des chromosomes lors de la méiose chez les femelles drosophiles et a noté qu'une perturbation de la distribution des chromosomes sexuels s'accompagnait de modifications de l'hérédité des traits liés au sexe.
Avec le développement de H. t. il a été constaté que les gènes situés sur le même chromosome constituent un groupe de liaison et doivent être hérités ensemble ; le nombre de groupes de liaison est égal au nombre de paires de chromosomes, constant pour chaque type d'organisme ; les traits qui dépendent de gènes liés sont également hérités ensemble. En conséquence, la loi de la combinaison indépendante de caractéristiques devrait avoir une application limitée ; les traits dont les gènes sont situés sur des chromosomes différents (non homologues) doivent être hérités indépendamment. Le phénomène de liaison incomplète des gènes (quand, avec les combinaisons parentales de traits, de nouvelles combinaisons recombinantes de ceux-ci sont trouvées dans la progéniture des croisements) a été étudié en détail par Morgan et ses collègues (A. G. Sturtevant et autres) et a servi de justification de l'arrangement linéaire des gènes dans les chromosomes. Morgan a suggéré que les gènes liés des chromosomes homologues, qui sont en combinaisons et chez les parents, peuvent changer de place pendant la méiose sous la forme hétérozygote ®, à la suite de quoi, avec les gamètes AB et ab, les gamètes Ab et aB se forment. De telles recombinaisons se produisent en raison de ruptures de chromosomes homologues dans la zone entre les gènes et de la connexion ultérieure des extrémités cassées dans une nouvelle combinaison : La réalité de ce processus, appelé croisement chromosomique, ou crossing over, a été prouvée en 1933 par le scientifique K. Stern dans des expériences avec la drosophile et les scientifiques américains X. Creightonomi B. McClintock - avec du maïs. Plus les gènes liés sont éloignés, plus ils sont susceptibles de se croiser. La dépendance de la fréquence de croisement sur les distances entre les gènes liés a été utilisée pour construire des cartes génétiques des chromosomes. Dans les années 30. 20ième siècle F. Dobzhansky a montré que l'ordre de placement des gènes sur les cartes génétiques et cytologiques des chromosomes coïncide.
Selon les idées de l'école de Morgan, les gènes sont des porteurs discrets et indivisibles d'informations héréditaires. Cependant, la découverte en 1925 par les scientifiques soviétiques G. A. Nadson et G. S. Filippov, et en 1927 par le scientifique américain G. Möller, de l'influence des rayons X sur la survenue de changements héréditaires (mutations) chez la drosophile, ainsi que la utilisation des rayons X pour accélérer le processus de mutation chez la drosophile En 1928-1930, les scientifiques soviétiques A. S. Serebrovsky, N. P. Dubinin et d'autres ont permis de formuler des idées sur la divisibilité d'un gène en unités plus petites disposées en une séquence linéaire et capables de changements mutationnels. En 1957, ces idées ont été prouvées par les travaux du scientifique américain S. Benzer avec le bactériophage T4. L'utilisation des rayons X pour stimuler les réarrangements chromosomiques a permis à N. P. Dubinin et B. N. Sidorov de découvrir en 1934 l'effet de la position d'un gène (découvert en 1925 par Sturtevant), c'est-à-dire la dépendance de la manifestation d'un gène sur sa localisation sur le chromosome. Il y avait une idée de l'unité de la discrétion et de la continuité dans la structure du chromosome.
H. t. n. se développe dans le sens d'un approfondissement des connaissances sur les porteurs universels d'informations héréditaires - les molécules d'acide désoxyribonucléique (ADN). Il a été établi qu'une séquence continue de bases puriques et pyrimidiques le long de la chaîne d'ADN forme des gènes, des intervalles intergéniques, des signes de début et de fin de lecture d'informations au sein d'un gène ; détermine le caractère héréditaire de la synthèse de protéines cellulaires spécifiques et, par conséquent, le caractère héréditaire du métabolisme. L'ADN constitue la base matérielle du groupe de liaison dans les bactéries et de nombreux virus (dans certains virus, l'acide ribonucléique est le support d'informations héréditaires) ; Les molécules d'ADN qui composent les mitochondries, les plastes et d'autres organites cellulaires servent de supports matériels de l'hérédité cytoplasmique.
H. t. n., expliquant les modèles d'hérédité des traits chez les animaux et les organismes végétaux, joue un rôle important dans la page - x. science et pratique. Il fournit aux éleveurs des méthodes de sélection de races animales et de variétés végétales aux propriétés souhaitées. Certaines dispositions de H. t. permettent de mener plus rationnellement page - x. production. Ainsi, le phénomène de l'héritage d'un certain nombre de signes liés à un étage à la page - x. animaux autorisés avant l'invention des méthodes de régulation artificielle du sexe dans ver à soie abattre les cocons du sexe le moins productif, avant de développer une méthode de séparation des poulets par sexe en examinant le cloaque - abattre les coqs, etc. La valeur principale pour l'augmentation de la productivité de plusieurs pages - x. cultures a l'usage de la polyploïdie. L'étude des maladies héréditaires humaines est basée sur la connaissance des modèles de réarrangements chromosomiques.
motifs, découvertes par l'école de Morgan, puis confirmées et approfondies sur de nombreux objets, sont connues sous le nom général de théorie chromosomique de l'hérédité.
Ses principales dispositions sont les suivantes :
1. Les gènes sont sur les chromosomes ; chaque chromosome est un groupe de liaison de gènes ; le nombre de groupes de liaison dans chaque espèce est égal au nombre de paires de chromosomes.
2. Chaque gène du chromosome occupe une certaine place (locus); les gènes sont disposés linéairement sur les chromosomes.
3. Les gènes alléliques sont échangés entre chromosomes homologues.
4. La distance entre les gènes (locus) dans le chromosome est proportionnelle au nombre de croisements entre eux.
Le fondateur de la théorie, Thomas Gent Morgan, un généticien américain, lauréat du prix Nobel, a émis une hypothèse sur la limitation des lois de Mendel.
Dans ses expériences, il a utilisé la mouche des fruits Drosophila, qui possède des qualités importantes pour les expériences génétiques : simplicité, fertilité, un petit nombre de chromosomes (quatre paires) et de nombreux traits alternatifs distincts.
Morgan et ses étudiants ont établi ce qui suit :
- Les gènes situés sur le même chromosome sont hérités ensemble ou liés.
- Des groupes de gènes situés sur le même chromosome forment des groupes de liaison. Le nombre de groupes de liaison est égal à l'ensemble haploïde de chromosomes chez les individus homogamétiques et n + 1 chez les individus hétérogamétiques.
- Entre chromosomes homologues, un échange de sites (crossing over) peut se produire ; à la suite du croisement, des gamètes apparaissent, dont les chromosomes contiennent de nouvelles combinaisons de gènes.
- La fréquence des croisements entre chromosomes homologues dépend de la distance entre les gènes situés sur le même chromosome. Plus cette distance est grande, plus la fréquence de coupure est élevée. Pour une unité de distance entre gènes, on prend 1 morganide (1% de croisement) ou le pourcentage d'occurrence d'individus croisés. Avec une valeur de cette valeur de 10 morganides, on peut affirmer que la fréquence de croisement chromosomique aux points de localisation de ces gènes est de 10 % et que de nouvelles combinaisons génétiques seront révélées chez 10 % de la progéniture.
- Pour déterminer la nature de la localisation des gènes dans les chromosomes et déterminer la fréquence de croisement entre eux, des cartes génétiques sont construites. La carte reflète l'ordre des gènes sur le chromosome et la distance entre les gènes sur le même chromosome. Ces conclusions de Morgan et de ses collaborateurs sont appelées théorie chromosomique de l'hérédité. Les conséquences les plus importantes de cette théorie sont les idées modernes sur le gène en tant qu'unité fonctionnelle de l'hérédité, sa divisibilité et sa capacité à interagir avec d'autres gènes.
Exemple d'héritage lié :
- Vg - ailes de drosophile normales ;
- vg - ailes rudimentaires;
- BB- coloration grise corps;
- bb - couleur de corps sombre.
Enregistrement dans l'expression chromosomique :
Dans ce cas, la règle d'uniformité des hybrides de la première génération est observée. Conformément aux deuxième et troisième lois de Mendel, 25 % de chacun des phénotypes possibles (mouches grises à longues ailes, mouches grises à ailes courtes, mouches noires à longues ailes et mouches noires à ailes courtes) seraient attendus lors d'analyses ultérieures. des croix. Cependant, les expériences de Morgan n'ont pas donné de tels résultats. Lorsqu'une femelle VgVgbb, récessive dans les deux traits, a été croisée avec un mâle hybride F1, 50% de mouches grises à ailes courtes et 50% de mouches à corps noir et ailes longues se sont formées :

Si une femelle dihybride est croisée avec un mâle récessif homozygote, une progéniture se forme: 41,5% - gris avec des ailes courtes, 41,5% - noir avec de longues ailes, 8,5% - gris avec de longues ailes, 8,5% - noir avec des ailes courtes. 
Ces résultats indiquent la présence d'une liaison génique et d'un croisement entre eux. Puisque 17 % des individus recombinants sont issus du second croisement, la distance entre les gènes Vg et B est de 17 %, soit 17 morganides.
héritage lié au sexe
Les ensembles de chromosomes de sexes différents diffèrent par la structure des chromosomes sexuels. Le chromosome Y mâle ne contient pas beaucoup des allèles trouvés sur le chromosome X. Les signes déterminés par les gènes des chromosomes sexuels sont dits liés au sexe. La nature de l'hérédité dépend de la distribution des chromosomes dans la méiose. Chez les sexes hétérogamétiques, des traits liés au chromosome X et n'ayant pas d'allèle sur le chromosome Y apparaissent même lorsque le gène qui détermine le développement de ces traits est récessif. Chez l'homme, le chromosome Y est transmis de père en fils et le chromosome X est transmis aux filles. Les enfants reçoivent le deuxième chromosome de leur mère. C'est toujours le chromosome X. Si la mère porte un gène pathologique récessif sur l'un des chromosomes X (par exemple, le gène du daltonisme ou de l'hémophilie), mais qu'elle-même n'est pas malade, alors elle est porteuse. Si ce gène est transmis aux fils, ils peuvent naître avec cette maladie, car il n'y a pas d'allèle sur le chromosome Y qui supprime le gène pathologique. Le sexe de l'organisme est déterminé au moment de la fécondation et dépend de l'ensemble de chromosomes du zygote résultant. Chez les oiseaux, les femelles sont hétérogamétiques et les mâles sont homogamétiques. Les abeilles n'ont pas de chromosomes sexuels. Les mâles sont haploïdes. Les abeilles femelles sont diploïdes.

Les principales dispositions de la théorie chromosomique de l'hérédité:
- chaque gène a un locus (place) spécifique dans le chromosome ;
- les gènes du chromosome sont situés dans une certaine séquence;
- les gènes d'un chromosome sont liés, ils sont donc hérités principalement ensemble ;
- la fréquence de croisement entre gènes est égale à la distance qui les sépare ;
- l'ensemble des chromosomes dans les cellules d'un type donné (caryotype) est un trait caractéristique de l'espèce.
Article pour le concours "bio/mol/text": 2015 marque les 100 ans théorie chromosomique de l'hérédité. Ses principales dispositions ont été formulées par T. Morgan, A. Sturtevant, G. Möller et K. Bridges dans le livre "The Mechanism of Mendelian Inheritance", publié à New York en 1915. Et plus tard, Thomas Morgan a reçu le premier prix Nobel "génétique" - pour avoir découvert le rôle des chromosomes dans l'hérédité. La conférence internationale "Chromosome 2015", qui s'est tenue en août 2015 à l'Académie de Novosibirsk, était consacrée à l'anniversaire de la théorie des chromosomes. Le texte suivant est le commentaire de l'auteur sur affiche sur l'histoire de la recherche sur les chromosomes, présenté à la conférence, et maintenant à la "Biomolécule" - dans la nomination compétitive la plus "vivante" " Visuellement sur la bien-aimée».
Noter!
Des informations plus complètes peuvent être trouvées dans le livre - Koryakov D.E., Zhimulev I.F. . Novossibirsk : Maison d'édition de la branche sibérienne de l'Académie russe des sciences, 2009 - 258 p., ISBN 978-5-7692-1045-7
Le sponsor de la nomination "Le meilleur article sur les mécanismes du vieillissement et de la longévité" est la Science for Life Extension Foundation. Le prix du choix du public a été sponsorisé par Helicon.
Commanditaires du concours : Laboratoire de solutions de bioimpression 3D pour la recherche en biotechnologie et Studio de sciences visuelles pour le graphisme scientifique, l'animation et la modélisation.
Le texte ci-dessous est un bref commentaire sur l'affiche, et des informations plus complètes peuvent être trouvées dans le livre: Koryakov D.E., Zhimulev I.F. Chromosomes. Structure et fonctions. Novossibirsk : Maison d'édition de la branche sibérienne de l'Académie russe des sciences, 2009 - 258 p., ISBN 978-5-7692-1045-7.
Cliquez sur l'image pour l'agrandir (s'ouvre dans une fenêtre séparée).
Le rôle génétique des chromosomes
Chaque organisme ne reproduit que sa propre espèce, et même dans les moindres traits de l'apparence et du comportement des enfants, on peut voir une ressemblance avec leurs parents. La première étape pour comprendre pourquoi cela se produit a été faite par un moine de la ville autrichienne de Brunn (maintenant c'est le tchèque Brno) G. Mendel ( G.Mendel). En 1865, lors d'une réunion de la Société des naturalistes de Brünn, il fait un rapport intitulé " Expériences sur des hybrides de plantes» ( Versuche über Pflanzen-Hybriden), et en 1866 il le publia dans le recueil des oeuvres de cette société. Le moine-naturaliste a décrit les résultats du croisement de différentes formes de pois et a suggéré la présence de facteurs spéciaux dont dépendent les signes extérieurs de la plante. Les modèles d'hérédité de ces facteurs ont été appelés plus tard Les lois de Mendel. Cependant, les contemporains n'ont pas compris la signification de cette découverte et l'ont oubliée, et ce n'est qu'en 1900 que G. de Vries ( H. de Vries, Pays-Bas), K. Korrens ( C. Correns, Allemagne) et E. Chermak ( E. Tschermak, Autriche) a redécouvert indépendamment les lois de Mendel.
Bien avant toutes ces études, qui s'appelleraient désormais analyses génétiques, les scientifiques impliqués dans la botanique, la zoologie, l'embryologie, l'histologie et la physiologie ont jeté les bases cytogénétique- la science des chromosomes. Dans divers articles et livres, la priorité de la découverte des chromosomes est donnée à personnes différentes, mais le plus souvent l'année de leur découverte s'appelle 1882, et leur découvreur est l'anatomiste allemand W. Flemming ( W. Flemming). Cependant, il serait plus juste de dire qu'il n'a pas découvert les chromosomes, mais seulement collectés et rationalisés dans son livre fondamental " Substance cellulaire, noyau et division cellulaire» ( Zellsubstanz, Kern et Zellteilung) tout ce que l'on savait d'eux à l'époque. Le terme même de "chromosome" a été introduit dans la science par l'histologue allemand H. Waldeyer ( H. Waldeyer) en 1888, et littéralement traduit, le terme signifie « corps peint ».
Maintenant, il est difficile de dire qui a fait la première description des chromosomes. En 1842, le botaniste suisse K. Naegeli ( C.Nageli) a publié un ouvrage dans lequel il dépeint quelques petits corps qui apparaissent à la place du noyau lors de la division cellulaire lors de la formation du pollen chez les lys et les tradescantia. C'étaient peut-être les premiers dessins de chromosomes. Première (1873) description détaillée mitose dans un ver plat Mésostome ehrenbergii appartient, comme on dit, au zoologiste allemand A. Schneider ( FA. Schneider). Il a décrit non seulement les étapes individuelles de la mitose, qui avaient été vues avant lui, mais toute la séquence de changements complexes dans le noyau : l'apparition de corps filamenteux à sa place, leur divergence dans des directions opposées et la formation de nouveaux noyaux dans les cellules filles. . Un autre type de division est méiose- décrit pour la première fois en détail par E. van Beneden ( E. van Beneden, Belgique) en 1883, observant la formation de gamètes chez les vers ronds. Il a découvert que dans la méiose, le nombre de chromosomes est divisé par deux et restauré pendant la fécondation et, malgré la différence de taille, les gamètes mâles et femelles apportent un nombre égal de chromosomes au zygote.
* - Un peu sur la place et le but des éléments génétiques mobiles dans les génomes pro- et eucaryotes : " Eléments génétiques mobiles des procaryotes : stratification de la « société » des vagabonds et des casaniers», « Le génome humain : un livre utile ou un magazine sur papier glacé ?», « L'ADN « indésirable » est-il le moteur de l'évolution des mammifères ?» - Éd.
Une autre option d'échange est échange de chromatides sœurs(SHO). Si les chromatides sont échangées lors du croisement différent chromosomes, puis dans le cas de CHO, les chromatides sont échangées à l'intérieur une chromosomes. Pour la première fois, le généticien américain D. Taylor a vu CHO ( J.Taylor) en 1958.
Le croisement, bien qu'ambigu, est associé à la formation dans la prophase de la méiose d'une structure spéciale à partir d'une paire de chromosomes homologues - complexe synaptonémique. Il a été découvert en 1956 indépendamment par deux cytologistes américains : M. Moses ( M. Moïse) chez les écrevisses et D. Focett ( D. Fawcett) chez une souris.
Diversité des chromosomes
Si nous entendons par chromosomes tous les porteurs d'informations héréditaires, alors ils sont extrêmement divers en taille, forme, apparence, composition et nombre. Les chromosomes des virus et des bactéries peuvent être circulaires ou linéaires. Les chromosomes des chloroplastes et des mitochondries sont de forme circulaire. Les chromosomes nucléaires des eucaryotes ont une forme linéaire, et ce sont eux sous la forme de corps X- et V-environ formes différentes viennent généralement à l'esprit lorsque l'on mentionne les chromosomes. Elles sont appelées mitotique ou métaphase, car ils ont cet aspect lors de la division - mitose (et la métaphase est l'une de ses étapes).
En 1912, le botaniste et cytologiste russe S.G. Navashin a montré que les chromosomes en métaphase ont un ensemble individuel de caractéristiques, notamment la taille, le rapport des longueurs de bras, la présence de satellites et de constrictions. En utilisant la position du centromère ou le rapport des longueurs des bras, S.G. Navashin a proposé une classification des chromosomes mitotiques, qui est encore utilisée aujourd'hui : métacentriques, sous-métacentriques, acrocentriques et télocentriques.
Le nombre de chromosomes dans différents types les organismes peuvent varier considérablement: de deux (dans une paire d'espèces végétales et une des fourmis australiennes) à 1440 dans une fougère Ophioglossum reticulatum et même 1600 chez les radiolaires marins Aulacantha scolymantha. Chez l'homme, le nombre de chromosomes est de 46, et il n'a été déterminé qu'en 1955, et publié en 1956 par le cytogénéticien d'origine chinoise D. Chio ( J. Tjio) en collaboration avec son leader A. Levan ( A. Levan) en Suède. Quelques mois plus tard, le chiffre est confirmé par le britannique C. Ford ( C.Ford) et D. Hamerton ( J. Hamerton). Il y a eu des tentatives pour déterminer le nombre de chromosomes humains depuis la fin du 19e siècle. Dans différents cas, il y avait différentes significations: 18, 24, 47 ou 48, - et seulement en 1955, ils étaient convaincus qu'une personne avait des chromosomes 46. En l'honneur de cet événement, une plaque commémorative avec l'image de la même plaque métaphasique, selon laquelle les chromosomes ont été comptés. Il est curieux que le nombre de chromosomes du chimpanzé (48) ait été découvert 15 ans plus tôt.
Il est généralement admis que le nombre de chromosomes dans chaque type d'organisme vivant est constant, et dans la grande majorité des cas, il l'est. Cependant, certains animaux et plantes ont des soi-disant surnuméraire, ou Additionnel, chromosomes. Tous les chromosomes de l'ensemble principal sont appelés Un chromosome. Ils sont toujours présents et la perte ou l'ajout d'un seul d'entre eux entraîne de graves conséquences. Les chromosomes supplémentaires sont appelés Chromosomes B, et leurs principales caractéristiques sont la présence facultative et la volatilité du nombre. Pour la première fois, des chromosomes surnuméraires ont été trouvés par E. Wilson ( E.Wilson, USA) en 1906 au bogue Metapodius terminalis.
Un type particulier de chromosome appelé chromosomes lampbrush, peut être observée dans la prophase de la première division de la méiose lors de la formation des ovocytes chez les oiseaux, les poissons, les reptiles et les amphibiens. Ils ont été mentionnés pour la première fois dans son livre fondamental (1882) par V. Fleming, qui a découvert ces chromosomes dans l'axolotl. Ils tirent leur nom de leur ressemblance avec une brosse pour nettoyer les lampes à pétrole.
Une place très spéciale parmi tous les types de chromosomes est occupée par chromosomes polytènes, qui ressemblent à de longs cordons épais avec des rayures transversales. Ils ont été découverts par l'embryologiste français E. Balbiani ( E. balbiani) en 1881 dans les noyaux cellulaires des glandes salivaires des larves de moustiques Chironomus plumeux. Les chromosomes polytènes ont joué un rôle remarquable dans le développement de la génétique, de la cytogénétique et de la biologie moléculaire. Avec leur aide, la linéarité de l'arrangement des gènes a été démontrée et le rôle génétique des chromosomes a été prouvé sans équivoque. Sur les chromosomes polytènes de la Drosophile, le polymorphisme chromosomique des populations sauvages a été décrit pour la première fois. C'est sur des chromosomes polytènes qu'ont été découverts les gènes des protéines de choc thermique, composants d'un système qui protège les cellules de tous les organismes des influences stressantes. Les chromosomes polytènes ont joué un rôle clé dans l'étude du système de compensation de dosage chez la drosophile.
Évolution des chromosomes et des génomes
Dans les études cytogénétiques modernes, un rôle important est joué par coloration différentielle. Pour la première fois, la capacité des chromosomes à se colorer différemment (c'est-à-dire de longueur inégale) a été démontrée par le britannique S. Darlington ( C.Darlington) et L. La Cour ( L. La Cour) en 1938. Une autre méthode de recherche importante est hybridation in situ, qui vous permet de déterminer la position de n'importe quel fragment d'ADN sur le chromosome. La méthode est basée sur la capacité acides nucléiques forment des molécules double brin, à la fois ADN-ADN et ARN-ADN. Cette méthode a été inventée en 1969 par D. Goll ( J. Gall) et M. Pardue ( M.Pardue) des États-Unis et H. John ( H.Jean), M. Birnstiel ( M. Birnstiel) et C. Jones ( K.Jones) du Royaume-Uni.
La combinaison de ces méthodes permet d'étudier en détail l'évolution des chromosomes et des génomes*, et les compagnons constants du processus évolutif sont réarrangements chromosomiques. Au fur et à mesure qu'une espèce évolue, des réarrangements se produisent inévitablement dans ses chromosomes qui modifient l'ordre des gènes par rapport à l'espèce ancestrale. Plus les espèces s'éloignent les unes des autres, plus les réarrangements chromosomiques les distinguent, et plus l'ordre des gènes change. connu différents types réarrangements : délétions (perte), duplications (doublement) et translocations (mouvement) de sections de chromosomes, qui ont été découvertes par C. Bridges en 1916, 1919 et 1923, respectivement. Un autre type est celui des inversions (rotation d'un segment chromosomique de 180°), décrites par A. Sturtevant en 1921. De plus, il existe un type spécial de réarrangement appelé translocation Robertsonienne (ou fusion centrée). Il a été décrit pour la première fois par l'Américain W. Robertson ( W.Robertson) en 1916, comparant les ensembles de chromosomes d'espèces de criquets étroitement apparentées. L'essence de ce réarrangement est réduite à la fusion de deux chromosomes acrocentriques en un seul métacentrique ou sous-métacentrique. Il existe également un processus inverse - séparation centrée. Dans ce cas, le chromosome méta- ou sous-métacentrique est divisé en deux chromosomes acrocentriques.
* - Sur la biomolécule, vous pouvez trouver une sélection impressionnante d'articles qui, d'une manière ou d'une autre, affectent l'évolution des génomes et des changements code génétique: « Les génomes viraux dans le système de l'évolution», « Sous le "gène harmonica"», « L'allopolyploïdie, ou comment des génomes différents ont appris à vivre sous le même toit», « Les génomes complets des pinsons des Galapagos ont enfin révélé les mécanismes de leur évolution», « Composition du génome eucaryote : endosymbiose VS. transfert horizontal continu»; « Le mystérieux code de notre génome», « Évolution du code génétique», « Aux origines du code génétique : les âmes sœurs», « Des synonymes si différents" et etc. - Éd.
Position des chromosomes dans le noyau
À fin XIX siècle T. Boveri a avancé l'idée que les chromosomes du noyau interphase ne sont pas mélangés au hasard, mais chacun d'eux occupe son propre espace. En 1909, il a inventé le terme " territoire chromosomique". La première preuve de l'existence de territoires chromosomiques n'a été obtenue qu'en 1982 par le chercheur allemand T. Kremer ( T. Cremer) avec des co-auteurs. Plus tard, ils ont visualisé ces territoires à l'aide de colorants fluorescents de différentes couleurs. Il s'est avéré que les grands chromosomes sont beaucoup plus susceptibles de se trouver dans la partie périphérique du noyau, tandis que les petits sont concentrés principalement dans la partie centrale. De plus, à la périphérie du noyau, il existe des régions de chromosomes appauvries en gènes. Les régions enrichies en gènes, au contraire, sont situées plus près du centre du noyau.
La composition des chromosomes. ADN
Les chromosomes sont des structures constituées d'un complexe complexe d'ADN, d'ARN et de protéines. Un tel complexe s'appelle chromatine.
L'ADN en tant que substance chimique a été découvert et isolé dans forme pure jeune chercheur suisse F. Miescher ( F.Miescher), travaillant en 1868-1869 à l'Université de la ville allemande de Tübingen. Il a étudié composition chimique leucocytes, dont la source était le pus des bandages de la clinique chirurgicale locale. F. Miescher a développé une méthode pour séparer les noyaux et le cytoplasme des cellules et a analysé la composition des noyaux. En plus des protéines et des lipides, il a découvert une substance qu'il a appelée nucléine(du mot noyau- noyau), et maintenant il est connu sous le nom d'ADN. Le fait que l'ADN soit porteur d'informations héréditaires a été établi pour la première fois en 1944 par les Américains O. Avery ( O. Avery), C. Mac Leod ( C. Mac Leod) et M. McCarthy ( M. McCarty) dans des expériences sur l'infection de souris par des pneumocoques.
La structure de la molécule d'ADN sous forme de double hélice a été déchiffrée en 1953 par F. Crick ( F.Crick), D. Watson ( J.Watson), M. Wilkins ( M.Wilkins) et R.Franklin ( R.Franklin) travaillant au Royaume-Uni. Pour cette découverte, les trois premiers chercheurs ont reçu le prix Nobel en 1962 (l'histoire de la découverte a été décrite de manière fascinante dans le livre " double hélice» James Watson hautement recommandé - Éd.). Rosalind Franklin ne fait pas partie des récipiendaires, car elle était décédée d'un cancer quatre ans plus tôt. On sait que la molécule d'ADN est constituée d'une séquence de quatre types de nucléotides : l'adénine, la thymine, la guanine et la cytosine*. Pour développer une méthode de détermination de leur séquence ( séquençage) en 1980, le prix Nobel a été décerné à P. Berg ( P. Berg, États-Unis), W. Gilbert ( W.Gilbert, États-Unis) et F. Sanger ( F. Sanger, Grande Bretagne).
* - En plus des quatre nucléotides "classiques" de l'ADN, on retrouve également leurs variants épigénétiquement modifiés : la méthylcytosine et la méthyladénine (" La sixième fondation ADN : de la découverte à la reconnaissance"). Et pour certains bactériophages Bacillus subtilis décrit l'inclusion dans l'ADN de l'uracile "ARN-nouveau" - Rouge.
Si le séquençage était initialement processus laborieux, qui ne permettait de "lire" qu'un petit fragment à la fois, puis à mesure que la technologie se développait, il devenait possible de déterminer, par exemple, la séquence complète ADN mitochondrial homme (1981). En 1990, un projet ambitieux est lancé avec pour objectif le séquençage complet du génome humain, et le premier résultat est présenté en 2001 (biomolécule : " Le génome humain : comment c'était et comment ce sera"). Parallèlement, le séquençage une le génome a coûté une somme colossale - des centaines de millions de dollars. Mais la technologie ne s'arrête pas là et l'émergence de nouvelles méthodes a réduit les coûts des milliers de fois*. Le séquençage du génome entier est désormais monnaie courante et, en 2009, le projet Genome 10K a été lancé. Son objectif est le séquençage et « l'assemblage » complet de 10 000 génomes animaux en chromosomes.
* - La "loi" de Moore est carrément vouée à atteindre des points limites dans diverses sciences (partout où il était possible de la tirer). La biologie a même dépassé l'électronique : la baisse progressive du coût du séquençage en 2007 a connu un pic abrupt, rapprochant l'ère de la lecture systématique du génome dans les points feldsher ruraux dans le cadre des polices d'assurance maladie obligatoires. Certes, dans un avenir prévisible, vous devrez toujours débourser - 1000 dollars plus les frais de transport: " Technologie : 1 000 $ par génome". Mais même cela ne pouvait qu'être rêvé avant l'avènement des nouvelles méthodes de séquençage de l'ADN : « Séquençage 454 (pyroséquençage d'ADN à haut débit)". Et pour comprendre les processus de base (au niveau cellulaire) du développement de l'organisme et de la victoire sur le cancer, il y a autre chose à rêver : " Séquençage unicellulaire (version - Metazoa)» - Éd.
Les nouvelles technologies ont permis le développement d'une direction telle que l'étude de l'ADN ancien (biomolécule : " ADN ancien : bonjour du passé"). Il est devenu possible d'isoler l'ADN d'os vieux de plusieurs dizaines de milliers d'années, et en 2008, par exemple, le génome mitochondrial de Néandertal a été séquencé. L'étude de l'ADN ancien, et en fait de toute la biologie moléculaire moderne, ne peut être imaginée sans l'utilisation de PCR - réaction en chaîne par polymérase. Pour sa découverte, l'américain K. Mullis ( K. Mullis) a reçu le prix Nobel 1993.
La composition des chromosomes. Écureuils
L'ADN dans les chromosomes subit plusieurs niveaux successifs d'encapsidation, et au tout premier niveau double hélice L'ADN s'enroule autour d'un globule de protéine nucléosome(biomolécule : " Roule, roule jusqu'à l'histone d'ADN"). Le globule contient quatre types de protéines appelées histones. En 1982, le biologiste moléculaire anglais A. Klug ( A.Klug) a reçu le prix Nobel pour avoir déchiffré la structure tridimensionnelle des nucléosomes. Indirectement, les nucléosomes ont reçu un autre prix Nobel - en 1910, il a été reçu par le biochimiste allemand A. Kossel ( A.Kossel) pour l'étude de la composition chimique des substances qui forment le noyau cellulaire, y compris la découverte des histones.
Les parties C-terminales des molécules d'histone sont étroitement repliées, tandis que les parties N-terminales n'ont pas de structure définie et divergent librement sur les côtés. En 1963-1964, on a découvert que certains résidus d'acides aminés dans les histones pouvaient être modifiés de manière covalente, c'est-à-dire acétylés ou méthylés. Maintenant, la liste des modifications s'est considérablement allongée, à la fois des groupes relativement simples - méthyle, acétyle, phosphate - et de grandes molécules complexes : la biotine, les oligopeptides ou les chaînes ADP-ribose peuvent être attachées aux résidus d'acides aminés. Les modifications apparaissent principalement sur les parties N- et, dans une bien moindre mesure, sur les parties C-terminales des molécules d'histone.
Selon théorie du code des histones, les modifications présentes sur les nucléosomes dans une région de chromatine donnée ne sont pas aléatoires, mais "codent" un processus. Ce point de vue a été formulé en 2000-2001 par B. Shtral ( B. Strahl, États-Unis), S. Ellis ( C.Allis, États-Unis) et T. Yenuwein ( T. Jenuwein, L'Autriche). Schématiquement, le processus du code histone peut être composé de trois étapes. Au premier stade, des enzymes agissent en modifiant certains résidus dans les histones. Au deuxième stade, les protéines qui ont des domaines spéciaux à cet effet se lient aux acides aminés modifiés. Chacun des domaines ne convient que pour "sa" modification. Au dernier stade, ces protéines liées attirent d'autres complexes protéiques, déclenchant ainsi une sorte de processus.
* - À propos de perspectives brillantes et de doutes qui donnent à réfléchir dans le domaine d'application de l'IPSC : " Des chercheurs français ont réussi à rajeunir les cellules de personnes centenaires», « Boule de neige des problèmes de pluripotence». - Éd.
Hétérochromatine
L'un des objets d'étude des divers processus épigénétiques est hétérochromatine. Il a été découvert en tant que régions plus sombres des chromosomes en 1907 par le cytologiste allemand S. Guthertz ( S. Gutherz), et les termes "hétérochromatine" et "euchromatine" ont été introduits en 1928 par un autre cytologiste allemand E. Heitz ( E. Heitz). En un mot, l'euchromatine est la partie des chromosomes dans laquelle se trouvent la grande majorité des gènes, tandis que l'hétérochromatine est principalement constituée de régions contenant de l'ADN non codant, constituées de séquences courtes et répétées. De plus, l'eu- et l'hétérochromatine diffèrent par le temps de réplication pendant la phase S du cycle cellulaire. Cette différence a été décrite pour la première fois en 1959 par A. Lima de Faria ( A. Lima-de-Faria, USA), étudiant le processus de réplication de l'ADN dans les testicules de criquets Melanoplus differentalis. Il a montré que l'hétérochromatine commence et finit de répliquer son ADN plus tard que l'euchromatine.
Une propriété importante de l'hétérochromatine est la capacité d'inactiver les gènes d'euchromatine qui y sont placés. Ce phénomène est appelé effet de position de type mosaïque. Il a été découvert en 1930 par G. Möller chez la drosophile. À la suite d'un réarrangement chromosomique, le gène blanche entré dans l'hétérochromatine. Ce gène est responsable de la couleur rouge des yeux, et s'il ne fonctionne pas, les yeux deviennent blancs. G. Möller, d'autre part, a produit des mouches dont les yeux n'étaient ni rouges ni blancs, mais tachetés, et chez différentes mouches, les taches étaient de formes et de tailles différentes. Cela est dû au fait que le gène lui-même reste intact, mais n'est inactivé que de manière aléatoire dans certaines cellules de l'œil et fonctionne dans d'autres.
Malgré de nombreuses années de recherche, le processus de formation de l'hétérochromatine est encore largement flou, en particulier sa toute première étape. On suppose qu'un processus similaire à Interférence ARN(biomolécule : " À propos de tous les ARN du monde, petits et grands"). Pour la découverte de ce phénomène, deux américains E. Fire ( Un feu) et K. Mello ( C. Mello) a reçu le prix Nobel en 2006. Le processus d'interférence est complexe et en plusieurs étapes, mais sans entrer dans les détails, l'introduction d'un ARN double brin homologue à un gène dans une cellule conduit à l'inactivation de ce gène.
Télomères
Une étude intensive des télomères a commencé après les américains E. Blackburn en 1978 ( E. Blackburn) et D. Goll ont séquencé le télomère chez les ciliés Tetrahymena thermophila. Il s'est avéré que les télomères contiennent une séquence de six nucléotides, répétée de 20 à 70 fois. En 1985, K. Greider ( C.greider) et E. Blackburn ont découvert une enzyme appelée télomérase, dont la tâche est d'achever la construction des télomères. En 2009, E. Blackburn, K. Greider et D. Shostak ( J. Szostak, USA) a reçu le prix Nobel pour l'étude des télomères et la découverte de l'enzyme télomérase (biomolécule : " Prix Nobel « sans âge » : en 2009, des travaux sur les télomères et la télomérase ont été reconnus», « Vieillissement - le prix à payer pour la suppression des tumeurs cancéreuses ?»).
Compensation de dose
Un grand nombre d'espèces d'organismes vivants, y compris les humains, ont des chromosomes sexuels non homologues, par exemple, X et Y. Cela nécessite un processus appelé compensation de dose. Son essence est la suivante : puisque le nombre d'autosomes est le même chez les mâles et les femelles, le nombre de gènes autosomiques, et donc le nombre de leurs produits, sera également le même. Mais le nombre de produits synthétisés à partir de gènes situés sur le chromosome sexuel sera 2 fois plus important chez un sexe que chez l'autre. Il s'avère une disproportion qu'il faut réguler d'une manière ou d'une autre, c'est-à-dire égaliser la « dose de gènes ». Le système de compensation de dose est appelé à résoudre ce problème (biomolécule : " , USA) ont émis l'hypothèse que chez les mammifères femelles l'un des deux chromosomes X est inactivé, et son choix est aléatoire. Ainsi, le système de compensation de dose chez les mammifères égalise le nombre de chromosomes X actifs dans les différents sexes : chez les mâles, il n'y a qu'un seul chromosome X, et chez les femelles, sur deux, un seul fonctionne.
Chez la drosophile, la nature a inventé un autre mécanisme, essentiellement à l'opposé de celui des mammifères : le seul Le chromosome X mâle est hyperactivé et fonctionne comme deux chromosomes X féminins. Le fait que l'activité totale de deux copies de n'importe quel gène du chromosome X chez les femmes et d'une copie chez les hommes de la drosophile soit la même a été découvert à l'aube du développement de la génétique. Cela a été fait par K. Stern en 1929 et G. Möller en 1931, de sorte que la drosophile est le premier organisme dans lequel une compensation de dose a été trouvée.
Et enfin...
Quelques mots sur la découverte, qui n'est pas directement liée aux chromosomes, mais qui est très activement utilisée, notamment pour étudier différents aspects de la vie des chromosomes. En 2008 O. Shimomura ( O. Shimomura), M. Chalfie ( M. Chalfie) et R. Tsien ( R.Tsien) des États-Unis a reçu le prix Nobel pour la découverte, l'isolement et l'application de protéine fluorescente verte (GFP) méduse Aequorea victoria. À l'aide de manipulations moléculaires, il est possible de connecter le gène de la protéine GFP au gène de n'importe quelle autre protéine et d'obtenir une protéine chimérique qui remplira à la fois sa fonction d'origine et son éclat. en vert. Cela permet de voir dans quelles cellules la protéine agit, dans le noyau ou le cytoplasme, dans quelles parties des chromosomes. En plus des protéines* fluorescentes vertes (GFP), rouges (RFP) et jaunes (YFP) sont maintenant connues.
* - À propos de la variété des protéines fluorescentes et de leur application dans recherche biologique les matériaux disent : Prix Nobel de chimie fluorescent», « Protéines fluorescentes : plus diversifiées que vous ne le pensez !», « "Dessinons" une cellule vivante". Et sur la bioluminescence dans les organismes terrestres et marins et le travail du système luciférine-luciférase - articles: " Bioluminescence : Résurgence», « lueur microscopique de l'échelle cosmique». - Éd.
Chaque cellule contient un certain nombre de chromosomes. Il y a beaucoup de gènes en eux. Une personne possède 23 paires (46) de chromosomes, soit environ 100 000 gènes, situés sur les chromosomes. De nombreux gènes sont situés sur un chromosome. Un chromosome avec tous les gènes qu'il contient forme un groupe de liaison. Le nombre de groupes de liaison est égal à l'ensemble haploïde de chromosomes. Une personne a 23 groupes de liaison. Les gènes situés sur le même chromosome ne sont pas absolument liés. Au cours de la méiose, lorsque les chromosomes sont conjugués, les chromosomes homologues échangent des parties. Ce phénomène est appelé crossing-over, qui peut se produire n'importe où sur le chromosome. Plus les loci sont éloignés les uns des autres sur le même chromosome, plus souvent un échange de sites peut se produire entre eux (Fig. 76).
Chez la mouche drosophile, les gènes de la longueur des ailes (V - long et v - court) et de la couleur du corps (B - gris et b - noir) se trouvent dans la même paire de chromosomes homologues, c'est-à-dire appartiennent au même groupe de liaison. Si vous croisez une mouche avec une couleur de corps grise et de longues ailes avec une mouche noire avec des ailes courtes, alors dans la première génération toutes les mouches auront une couleur de corps grise et de longues ailes (Fig. 77).
À la suite du croisement d'un mâle dihétérozygote avec une femelle récessive homozygote, les mouches ressembleront à leurs parents. En effet, les gènes d'un même chromosome sont hérités de manière liée. Chez la mouche Drosophile mâle, la couvée est complète. Si vous croisez une femelle dihétérozygote avec un mâle homozygote récessif, alors certaines mouches ressembleront à leurs parents, et en
Riz. 76. Traverser.
1 - deux chromosomes homologues ; 2 - leur décussation pendant la conjugaison; 3 - deux nouvelles combinaisons de chromosomes.
d'autre part, une recombinaison de caractéristiques se produira. Un tel héritage a lieu pour les gènes du même groupe de liaison, entre lesquels un croisement peut se produire. Ceci est un exemple de liaison incomplète de gènes.
Les principales dispositions de la théorie chromosomique de l'hérédité
. Les gènes sont situés sur les chromosomes.
. Les gènes sont disposés linéairement sur un chromosome.

Riz. 77.Héritage lié des gènes de la couleur du corps et de l'état des ailes chez la mouche des fruits.
Le gène gris (B) domine le gène de la couleur du corps noir (b), le gène des ailes longues (V) domine le gène des ailes courtes (v). B et V sont sur le même chromosome.
a - liaison complète des gènes en raison de l'absence de croisement chromosomique chez les mâles de Drosophile : PP - une femelle grise à longues ailes (BBVV) est croisée avec un mâle noir à ailes courtes (bbvv) ; F 1 - mâle gris aux longues ailes (BbVv) croisé avec une femelle noire aux ailes courtes (bbvv); F 2 - puisque le mâle ne se croise pas, deux types de progéniture apparaîtront : 50 % - noir à ailes courtes et 50 % - gris avec des ailes normales ; b - liaison incomplète (partielle) des traits due au croisement chromosomique chez les femelles de Drosophile : PP - une femelle aux longues ailes (BBVV) est croisée avec un mâle noir aux ailes courtes (bbvv) ; F 1 - une femelle grise aux longues ailes (BbVv) est croisée avec un mâle noir aux ailes courtes (bbvv). F 2 - puisque le croisement de chromosomes homologues se produit chez la femelle, quatre types de gamètes se forment et quatre types de progéniture apparaîtront: non croisés - gris à longues ailes (BbVv) et noir à ailes courtes (bbvv), croisés - noir à longues ailes (bbVv), gris à ailes courtes (Bbvv).
. Chaque gène occupe une place spécifique - un locus.
. Chaque chromosome est un groupe de liaison. Le nombre de groupes de liaison est égal au nombre haploïde de chromosomes.
Les gènes alléliques sont échangés entre chromosomes homologues. La distance entre les gènes est proportionnelle au pourcentage de croisement entre eux.
Questions pour la maîtrise de soi
1. Où se trouvent les gènes ?
2. Qu'est-ce qu'un groupe d'embrayage ?
3. Quel est le nombre de groupes de liaison ?
4. Comment les gènes sont-ils liés dans les chromosomes ?
5. Comment le trait de la longueur des ailes et de la couleur du corps est-il hérité chez les mouches drosophiles ?
6. Quelle progéniture apparaîtra lors du croisement d'une femelle homozygote avec de longues ailes et en gris corps avec un mâle noir homozygote aux ailes courtes ?
7. Quelle progéniture apparaîtra lorsqu'un mâle dihétérozygote est croisé avec une femelle homozygote récessive ?
8. Quel type de liaison génétique la drosophile mâle possède-t-elle ?
9. Quelle sera la progéniture lorsqu'une femelle dihétérozygote est croisée avec un mâle homozygote récessif ?
10. Quel type de liaison génétique la femelle Drosophile possède-t-elle ?
11. Quelles sont les principales dispositions de la théorie chromosomique de l'hérédité ?
Mots clés du sujet " Théorie chromosomique hérédité"
gènes
groupe d'embrayage
longueur
cellules
conjugaison
traverser
ailes
mouche locus linéaire
hérédité
échanger
coloration
couple organisme
recombinaison
génération
position
descendance
distance
résultat
Parents
Masculin
femelle
traversée
corps
la théorie
terrain
chromosomes
Couleur
partie
Humain
Numéro
Mécanisme de détermination du sexe chromosomique
Les différences phénotypiques entre individus de sexes différents sont dues au génotype. Les gènes sont situés sur les chromosomes. Il existe des règles d'individualité, de constance, d'appariement des chromosomes. L'ensemble diploïde des chromosomes est appelé caryotype. Il y a 23 paires (46) de chromosomes dans les caryotypes féminin et masculin (Fig. 78).
22 paires de chromosomes sont identiques. Elles sont appelées autosomes. 23ème paire de chromosomes - chromosomes sexuels. Dans le caryotype féminin, un

Riz. 78.Caryotypes de divers organismes.1 - une personne; 2 - moustique; 3 plantes skerda.
les chromosomes sexuels XX. Dans le caryotype masculin, les chromosomes sexuels sont XY. Le chromosome Y est très petit et contient peu de gènes. La combinaison des chromosomes sexuels dans le zygote détermine le sexe du futur organisme.
Au cours de la maturation des cellules germinales, à la suite de la méiose, les gamètes reçoivent un ensemble haploïde de chromosomes. Chaque œuf contient 22 autosomes + un chromosome X. Le sexe qui produit des gamètes identiques sur le chromosome sexuel est appelé sexe homogamétique. La moitié des spermatozoïdes contiennent - 22 autosomes + chromosome X, et la moitié 22 autosomes + Y. Le sexe qui forme des gamètes différents sur le chromosome sexuel est appelé hétérogamétique. Le sexe de l'enfant à naître est déterminé au moment de la fécondation. Si un ovule est fécondé par un spermatozoïde du chromosome X, il se développe corps féminin si le chromosome Y est mâle (Fig. 79).

Riz. 79.Mécanisme chromosomique de formation du sexe.
La probabilité d'avoir un garçon ou une fille est de 1:1 soit 50%:50%. Cette définition du sexe est typique pour les humains et les mammifères. Certains insectes (sauterelles et cafards) n'ont pas de chromosome Y. Les mâles ont un chromosome X (X0) et les femelles en ont deux (XX). Chez les abeilles, les femelles ont 2n ensembles de chromosomes (32 chromosomes), tandis que les mâles ont n (16 chromosomes). Les femmes ont deux chromosomes X sexuels dans leurs cellules somatiques. L'un d'eux forme un morceau de chromatine, qui peut être vu dans les noyaux d'interphase lorsqu'il est traité avec un réactif. Cette masse est le corps de Barr. Les mâles n'ont pas de corps de Barr car ils n'ont qu'un seul chromosome X. Si, au cours de la méiose, deux chromosomes XX pénètrent dans l'ovule en même temps et qu'un tel ovule est fécondé par un spermatozoïde, le zygote aura alors un plus grand nombre de chromosomes.

Par exemple, un organisme avec un ensemble de chromosomes XXX (trisomie sur le chromosome X) le phénotype est une fille. Elle a des gonades sous-développées. Il existe deux corps de Barr dans les noyaux des cellules somatiques.
Un organisme avec un ensemble de chromosomes XXY (syndrome de Klinefelter) le phénotype est un garçon. Ses testicules sont sous-développés, un retard physique et mental est noté. Il y a un corps de Barr.
Chromosomes XO (monosomie sur le chromosome X)- déterminer Syndrome de Shereshevsky-Turner. Un organisme avec un tel ensemble est une fille. Elle a des gonades sous-développées, une petite taille. Pas de corps de Barr. Un organisme qui n'a pas de chromosome X, mais qui ne contient qu'un chromosome Y, n'est pas viable.
L'hérédité des traits dont les gènes sont situés sur les chromosomes X ou Y est appelée hérédité liée au sexe. Si les gènes se trouvent sur les chromosomes sexuels, ils sont hérités liés au sexe.
Une personne possède un gène sur le chromosome X qui détermine le signe de la coagulation du sang. Le gène récessif provoque le développement de l'hémophilie. Le chromosome X possède un gène (récessif) qui est responsable de la manifestation du daltonisme. Les femmes ont deux chromosomes X. Un trait récessif (hémophilie, daltonisme) n'apparaît que si les gènes qui en sont responsables sont localisés sur deux chromosomes X : X h X h ; X ré X ré . Si un chromosome X a un gène H ou D dominant et que l'autre a un gène h ou d récessif, il n'y aura pas d'hémophilie ni de daltonisme. Les hommes ont un chromosome X. S'il a un gène H ou h, alors ces gènes montreront certainement leur effet, car le chromosome Y ne porte pas ces gènes.
Une femme peut être homozygote ou hétérozygote pour les gènes situés sur le chromosome X, mais les gènes récessifs n'apparaissent qu'à l'état homozygote.

Si les gènes sont sur le chromosome Y (héritage hollandais), puis les signes conditionnés par eux se transmettent de père en fils. Par exemple, la pilosité des oreilles est héritée du chromosome Y. Les hommes ont un chromosome X. Tous les gènes qu'il contient, y compris les gènes récessifs, apparaissent dans le phénotype. Chez le sexe hétérogamétique (mâle), la plupart des gènes situés sur le chromosome X sont situés sur hémizygoteétat, c'est-à-dire qu'ils n'ont pas de paire allélique.
Le chromosome Y contient certains gènes homologues aux gènes du chromosome X, par exemple les gènes de la diathèse hémorragique, du daltonisme général, etc. Ces gènes sont hérités à la fois par le chromosome X et par le chromosome Y.
Questions pour la maîtrise de soi
1. Quelles sont les règles des chromosomes ?
2. Qu'est-ce qu'un caryotype ?
3. Combien d'autosomes une personne possède-t-elle ?
4. Quels chromosomes humains sont responsables du développement du sexe ?
5. Quelle est la probabilité d'avoir un garçon ou une fille ?
6. Comment le sexe est-il déterminé chez les sauterelles et les cafards ?
7. Comment le sexe est-il déterminé chez les abeilles ?
8. Comment le sexe est-il déterminé chez les papillons et les oiseaux ?
9. Qu'est-ce qu'un corps de Barr ?
10. Comment pouvez-vous déterminer la présence d'un corps de Barr ?
11. Qu'est-ce qui peut expliquer l'apparition de plus ou moins de chromosomes dans le caryotype ?
12. Qu'est-ce que l'héritage lié au sexe ?
13. Quels gènes sont liés au sexe chez l'homme ?
14. Comment et pourquoi les gènes récessifs liés au sexe montrent-ils leur effet chez les femmes ?
15. Comment et pourquoi les gènes récessifs liés au chromosome X chez l'homme montrent-ils leur effet ?
Mots clés du thème "Détermination chromosomique du sexe"
autosomes
papillons
probabilité
pilosité des oreilles
gamètes
génotype
gènes
sexe hétérogamétique
morceau de chromatine
sexe homogamétique
daltonisme
fille
action
femme
zygote
individualité
caryotype
sauterelles
garçon
méiose
mammifère
moment
monosomie
le mâle
trousse
insectes
héritage
transporteur
traitement réactif de fécondation
organisme
individuel
jumelage
des couples
sol
cellules sexuelles
progéniture
règlements
pancarte
des oiseaux
les abeilles
développement
différences
naissance
croissance
syndrome de coagulation du sang des testicules
syndrome de Klinefelter
Syndrome de Shershevsky-Turner
cécité
maturation
condition
combinaison
spermatozoïdes
fils
les cafards
Corps de barre
trisomie
Chromosome Y
phénotype
chromosome
Chromosome X
Humain
noyau
Oeuf
§ 5. T. G. Morgan et sa théorie des chromosomes
Thomas Gent Morgan est né en 1866 dans le Kentucky (États-Unis). Après avoir obtenu son diplôme universitaire à vingt ans, Morgan a reçu le titre de docteur en sciences à vingt-quatre ans et à vingt-cinq ans, il est devenu professeur.
Depuis 1890, Morgan s'est engagé dans l'embryologie expérimentale. Dans la première décennie du XXe siècle, il affectionne les questions d'hérédité.

Cela semble paradoxal, mais au début de son activité, Morgan était un ardent opposant aux enseignements de Mendel et allait réfuter ses lois sur les objets animaux - les lapins. Cependant, les administrateurs de l'Université de Columbia ont trouvé l'expérience trop coûteuse. Alors Morgan a commencé ses recherches sur un objet moins cher - la mouche des fruits Drosophila, et non seulement n'a pas nié les lois de Mendel, mais est également devenu un digne successeur de ses enseignements.

Un chercheur dans des expériences avec la drosophile crée théorie chromosomique de l'hérédité- la plus grande découverte, occupant, par expression N. K. Koltsova, "la même place en biologie que la théorie moléculaire en chimie et la théorie des structures atomiques en physique."
En 1909-1911. Morgan et ses élèves tout aussi illustres A. Sturtevant, G. Moeller, C. Ponts ont montré que la troisième loi de Mendel nécessite des ajouts importants : les inclinations héréditaires ne sont pas toujours héritées indépendamment ; parfois, ils sont transmis en groupes entiers - liés les uns aux autres. De tels groupes situés sur le chromosome correspondant peuvent se déplacer vers un autre chromosome homologue lors de la conjugaison des chromosomes au cours de la méiose (prophase I).
La théorie du chromosome complet a été formulée TG Morgan dans la période de 1911 à 1926. Avec son apparence et la poursuite du développement cette théorie est redevable non seulement à Morgan et à son école, mais aussi au travail d'un nombre important de scientifiques, étrangers et nationaux, parmi lesquels, tout d'abord, il convient de mentionner N. K. Koltsova et A. S. Serebrovsky (1872-1940).
Selon la théorie des chromosomes, la transmission de l'information héréditaire est associée aux chromosomes, dans lequel linéairement, à un certain lieu (de lat. lieu- lieu), les gènes mentent. Puisque les chromosomes sont appariés, chaque gène sur un chromosome correspond à un gène apparié sur l'autre chromosome (homologue) situé dans le même locus. Ces gènes peuvent être identiques (chez les homozygotes) ou différents (chez les hétérozygotes). Formes variées les gènes qui résultent d'une mutation de l'original sont appelés allèles, ou allélomorphes(du grec allo - différent, morph - forme). Les allèles affectent la manifestation d'un trait de différentes manières. Si un gène existe dans plus de deux états alléliques, alors ces allèles dans populations* forment une série d'allèles dits multiples. Chaque individu d'une population peut contenir deux (mais pas plus) allèles dans son génotype, et chaque gamète ne peut contenir qu'un seul allèle, respectivement. Dans le même temps, les individus avec n'importe quel allèle de cette série peuvent faire partie de la population. Les allèles de l'hémoglobine sont un exemple d'allèles multiples (voir Chapitre I, § 5).
* (Une population (du latin popularus - population) est un groupe d'individus de la même espèce, unis par croisement mutuel, dans une certaine mesure isolés des autres groupes d'individus de cette espèce.)
Le degré de dominance dans une série d'allèles peut augmenter du gène récessif extrême à l'extrême dominant. peut mener grand nombre exemples de ce type. Ainsi, chez le lapin, la série de gènes récessifs allèles multiples est le gène c qui détermine le développement de l'albinisme*. Le gène c h de coloration himalayenne (hermine) (yeux roses, corps blanc, bouts du nez, des oreilles, de la queue et des membres foncés) sera dominant par rapport à ce gène ; sur ce gène, ainsi que sur le gène c, le gène de couleur gris clair (chinchilla) c ch domine. Une étape encore plus dominante est le gène agouti - c a (domine sur les gènes c, c h et c ch). Le plus dominant de toute la série, le gène de couleur noire C domine toutes les "étapes inférieures des allèles" - gènes c, c h, c ch, c a.
* (Absence de pigmentation (voir chapitre VII, § 5).)
La dominance, comme la récessivité des allèles, n'est pas un absolu, mais leur propriété relative. Le degré de dominance et de récessivité peut être différent. Le même trait peut être hérité de manière dominante ou récessive.
Ainsi, par exemple, le pli au-dessus du coin interne de l'œil (epicanthus) est hérité de manière dominante chez les Mongoloïdes et de manière récessive chez les Négroïdes (Bushmen, Hottentots).
En règle générale, les allèles nouvellement émergents sont récessifs, au contraire, les allèles d'anciennes variétés de plantes ou de races animales (encore plus d'espèces sauvages) sont dominants.
Chaque paire de chromosomes est caractérisée par un certain ensemble de gènes qui composent le groupe de liaison. C'est pourquoi les groupes différents signes parfois hérités ensemble.
Étant donné que les cellules somatiques de la drosophile contiennent quatre paires de chromosomes (2n = 8) et que les cellules sexuelles en contiennent deux fois moins (1n = 4), la mouche des fruits a quatre groupes Embrayage; de même, chez l'homme, le nombre de groupes de liaison est égal au nombre de chromosomes de l'ensemble haploïde (23).
Pour un certain nombre d'organismes (drosophile, maïs) et certains chromosomes humains *, des cartes chromosomiques ou génétiques ont été compilées, qui sont un arrangement schématique des gènes dans les chromosomes.
* (Jusqu'à présent, il n'a été possible d'établir la localisation exacte des gènes humains (si l'on tient compte du nombre total de gènes) que dans un petit nombre et relativement Cas rares, par exemple, pour les traits liés aux chromosomes sexuels.)

A titre d'exemple, donnons une carte chromosomique d'une partie du chromosome X de la Drosophile (Fig. 24). Avec plus ou moins de précision, cette carte reflète la séquence des gènes et la distance qui les sépare. Il a été possible de déterminer la distance entre les gènes à l'aide d'analyses génétiques et cytologiques du croisement, qui se produit lors de la conjugaison des chromosomes homologues au cours du zygonème de la prophase I de la méiose (voir Chapitre II, § 7).

Le mouvement des gènes d'un chromosome à l'autre se produit avec une certaine fréquence, qui est inversement proportionnel à la distance entre les gènes : plus la distance est courte, plus pourcentage de croisement(l'unité de distance entre les gènes porte le nom de Morgan Morgane et est égal à la distance minimale dans le chromosome qui peut être mesurée par croisement). Le croisement est illustré à la Fig. 25.
À l'heure actuelle, la liaison étroite de certains loci de gènes est connue et le pourcentage de croisement a été calculé pour eux. Les gènes liés déterminent, par exemple, l'expression Facteur Rh et les gènes du système MN du sang (sur l'hérédité des propriétés du sang, voir Chapitre VII, § 3). Dans certaines familles, il a été possible de retracer le lien du facteur Rh avec ovalocytose(la présence d'environ 80 à 90% d'érythrocytes de forme ovale - l'anomalie se déroule, en règle générale, sans manifestations cliniques), ce qui donne environ 3% du croisement. On observe jusqu'à 9 % de croisement entre les gènes qui contrôlent les manifestations des groupes sanguins ABO et le facteur Lu. On sait que le gène qui affecte l'anomalie de la structure des ongles et du genou est également lié aux loci du système ABO ; le pourcentage de croisement entre eux est d'environ 10. Les groupes de liaison (et, par conséquent, les cartes chromosomiques) des chromosomes humains X et Y sont beaucoup mieux étudiés (voir Chapitre VII, § 6). On sait, par exemple, que les gènes qui déterminent le développement de daltonisme(daltonisme) et hémophilie(saignement); le pourcentage de chevauchement entre eux est de 10.
L'exactitude de l'hypothèse de Morgan a été confirmée au début du siècle par Kurt Stern (études cytologiques) et les collaborateurs de Morgan Theophilus Painter (cytologiste) et Calvin Bridges (généticien) sur les chromosomes géants des glandes salivaires des larves de Drosophila (semblables au géant chromosomes d'autres diptères). Sur la fig. 26 montre une partie du chromosome géant de la glande salivaire de la larve de chironomus (ver de vase).

Lors de l'étude de chromosomes géants avec un microscope optique conventionnel, la strie transversale est clairement visible, formée par l'alternance de bandes de disques claires et plus foncées - chromomères; ils sont formés par des zones fortement spiralées et densément adjacentes.
La formation de ces chromosomes géants est appelée polythénie, c'est-à-dire la reduplication des chromosomes sans augmenter leur nombre. Dans le même temps, les chromatides redoublées restent côte à côte, étroitement jointives les unes aux autres.
Si un chromosome constitué d'une paire de chromatides double consécutivement neuf fois, le nombre de brins (chromonèmes) dans un tel chromosome polytène sera de 1024. En raison de la déspiralisation partielle des chromonèmes, la longueur d'un tel chromosome augmente par rapport à la longueur habituelle. un par 150-200 fois.
En 1925, Sturtevant montra la présence croisement inégal : dans l'un des chromosomes homologues, il peut y avoir deux locus identiques, dans lesquels, par exemple, des gènes qui affectent la forme de l'œil de la drosophile - Bar sont situés, et dans l'autre - pas un seul locus. C'est ainsi que vole avec un signe prononcé d'yeux rayés étroits (gène barre ultra)(voir figure 31).
En plus des preuves cytologiques de l'exactitude de la théorie des chromosomes, des expériences génétiques ont été menées - en croisant différentes races de drosophiles. Ainsi, parmi les nombreux gènes liés chez la mouche des fruits, il existe deux gènes récessifs : le gène de la couleur noire du corps ( le noir) et le gène des ailes rudimentaires ( rudimentaire).
Appelons-les gènes a et b. Ils correspondent à deux allèles dominants : le gène du corps gris et des ailes normalement développées (A et B). Lors du croisement de mouches de race pure aabb et AABB, toute la première génération d'hybrides aura le génotype AaBb. Théoriquement parlant, les résultats suivants devraient être attendus dans la deuxième génération (F 2).

Cependant, dans un pourcentage faible mais constant de cas, des descendants inhabituels de gamètes inhabituels ont été rencontrés. Environ 18 % de ces gamètes ont été observés à chaque croisement (9 % Ab et 9 % aB).

La survenance de telles exceptions est bien expliquée par le processus de croisement. Ainsi, des études génétiques ont également permis d'établir que le trouble de l'embrayage est le croisement, entraînant une augmentation de la variabilité de la forme, est statistiquement constant.
En conclusion, notons qu'un certain nombre de dispositions de la génétique classique ont subi aujourd'hui un certain nombre de modifications.
Nous avons utilisé à plusieurs reprises les termes gènes (allèles) et traits "dominants" et "récessifs". Cependant, des études récentes ont montré que les gènes dits récessifs peuvent en fait ne pas être récessifs du tout. Il est plus correct de dire que les gènes récessifs donnent une très faible manifestation visible ou invisible dans le phénotype. Mais dans ce dernier cas, des allèles récessifs, invisibles de l'extérieur dans le phénotype, peuvent être détectés à l'aide de techniques biochimiques spéciales. De plus, le même gène dans certaines conditions environnementales peut se comporter comme dominant, dans d'autres - comme récessif.
Étant donné que le développement de tous les organismes se produit en fonction et sous l'influence de l'environnement extérieur, la manifestation du génotype dans un certain phénotype est également influencée par des facteurs environnementaux (température, nourriture, humidité et composition gazeuse de l'atmosphère, sa pression, la présence de formes pathogènes pour un organisme donné, la composition chimique de l'eau, du sol, etc., mais pour l'homme et les phénomènes l'ordre social). Le phénotype ne montre jamais toutes les possibilités génotypiques. Par conséquent, dans conditions différentes les manifestations phénotypiques de génotypes étroitement apparentés peuvent être très différentes les unes des autres. Ainsi, à la fois le génotype et l'environnement sont impliqués (dans une plus ou moins grande mesure) dans la manifestation d'un trait.