Quelle est la molécule d'ADN dans les mitochondries. Caractéristiques de la structure de l'ADN mitochondrial
Lire aussi
© G.M. Dymshits
Surprises du génome mitochondrial
G. M. Dymmerdes
Grigory Moiseevich Dymshits, Docteur en sciences biologiques, professeur du département de biologie moléculaire, Université d'État de Novosibirsk, chef du laboratoire de structure du génome, institut de cytologie et de génétique, branche sibérienne de l'Académie russe des sciences. Co-auteur et éditeur de quatre manuels scolaires de biologie générale.Un quart de siècle s'est écoulé depuis la découverte des molécules d'ADN dans les mitochondries avant qu'ils ne s'intéressent non seulement aux biologistes moléculaires et aux cytologistes, mais aussi à la génétique, aux évolutionnistes, ainsi qu'aux paléontologues et médecins légistes, historiens et linguistes. Un tel intérêt a été suscité par les travaux d'A. Wilson de l'Université de Californie. En 1987, il publie les résultats analyse comparative ADN de mitochondries prélevé sur 147 représentants de différents groupes ethniques de toutes les races humaines habitant les cinq continents. Selon le type, l'emplacement et le nombre de mutations individuelles, il a été établi que tout l'ADN mitochondrial provenait de la même séquence nucléotidique ancestrale par divergence. Dans la presse pseudo-scientifique, cette conclusion a été interprétée de manière extrêmement simplifiée - toute l'humanité est venue d'une seule femme, appelée Eve mitochondriale (les filles et les fils ne reçoivent des mitochondries que de leur mère), qui vivait en Afrique du Nord-Est il y a environ 200 000 ans. Après encore 10 ans, il a été possible de déchiffrer un fragment d'ADN mitochondrial isolé des restes d'un Néandertalien et d'estimer le temps d'existence du dernier ancêtre commun de l'homme et de Néandertal à 500 000 ans.
Aujourd'hui, la génétique mitochondriale humaine se développe intensivement tant dans la population que dans aspect médical. Un lien a été établi entre un certain nombre de maladies héréditaires graves et des défauts de l'ADN mitochondrial. Les changements génétiques associés au vieillissement sont plus prononcés dans les mitochondries. Qu'est-ce que le génome mitochondrial, qui diffère chez l'homme et les autres animaux de celui des plantes, des champignons et des protozoaires en termes de taille, de forme et de capacité génétique ? Comment cela fonctionne-t-il et comment le génome mitochondrial est-il originaire de différents taxons ? Cela sera discuté dans notre article.
Les mitochondries sont appelées les centrales électriques de la cellule. En plus de la membrane lisse externe, ils ont une membrane interne qui forme de nombreux plis - crêtes. Les composants protéiques de la chaîne respiratoire y sont intégrés - des enzymes impliquées dans la conversion de l'énergie des liaisons chimiques des nutriments oxydés en énergie des molécules d'acide adénosine triphosphorique (ATP). Avec une telle « monnaie convertible », la cellule paie tous ses besoins énergétiques. Dans les cellules des plantes vertes, en plus des mitochondries, il existe également d'autres stations énergétiques - les chloroplastes. Ils fonctionnent sur des "batteries solaires", mais ils forment également de l'ATP à partir d'ADP et de phosphate. Comme les mitochondries, les chloroplastes - organites à réplication autonome - ont également deux membranes et contiennent de l'ADN.
En plus de l'ADN, la matrice mitochondriale contient ses propres ribosomes, qui diffèrent par de nombreuses caractéristiques des ribosomes eucaryotes situés sur les membranes du réticulum endoplasmique. Cependant, les ribosomes mitochondriaux ne forment pas plus de 5% de toutes les protéines qui entrent dans leur composition. La plupart des protéines qui composent les composants structurels et fonctionnels des mitochondries sont codées par le génome nucléaire, synthétisées sur les ribosomes du réticulum endoplasmique et transportées par ses canaux jusqu'au site d'assemblage. Ainsi, les mitochondries sont le résultat des efforts conjugués de deux génomes et de deux appareils de transcription et de traduction. Certaines enzymes sous-unités de la chaîne respiratoire mitochondriale sont constituées de différents polypeptides, dont certains sont codés par le nucléaire et d'autres par le génome mitochondrial. Par exemple, l'enzyme clé de la phosphorylation oxydative, la cytochrome c oxydase, chez la levure se compose de trois sous-unités codées et synthétisées dans les mitochondries et de quatre sous-unités codées dans le noyau cellulaire et synthétisées dans le cytoplasme. L'expression de la plupart des gènes mitochondriaux est contrôlée par certains gènes nucléaires.
Tailles et formes des génomes mitochondriaux
À ce jour, plus de 100 génomes mitochondriaux différents ont été lus. L'ensemble et le nombre de leurs gènes dans l'ADN mitochondrial, dont la séquence nucléotidique est complètement déterminée, varient considérablement selon les espèces d'animaux, de plantes, de champignons et de protozoaires. Le plus grand nombre gènes trouvés dans le génome mitochondrial du protozoaire flagellé Rectinomonas americana- 97 gènes, y compris tous les gènes codant pour les protéines trouvés dans l'ADNmt d'autres organismes. Chez la plupart des animaux supérieurs, le génome mitochondrial contient 37 gènes : 13 pour les protéines de la chaîne respiratoire, 22 pour l'ARNt et deux pour l'ARNr (pour la grande sous-unité ribosomale ARNr 16S et pour le petit ARNr 12S). Chez les plantes et les protozoaires, contrairement aux animaux et à la plupart des champignons, le génome mitochondrial code également certaines protéines qui composent les ribosomes de ces organites. Les enzymes clés de la synthèse des polynucléotides matrices, telles que l'ADN polymérase (répliquant l'ADN mitochondrial) et l'ARN polymérase (transcrivant le génome mitochondrial), sont codées dans le noyau et synthétisées sur les ribosomes cytoplasmiques. Ce fait indique l'autonomie relative des mitochondries dans la hiérarchie complexe de la cellule eucaryote.
Les génomes mitochondriaux des différentes espèces diffèrent non seulement par l'ensemble des gènes, l'ordre de leur emplacement et de leur expression, mais aussi par la taille et la forme de l'ADN. La grande majorité des génomes mitochondriaux décrits aujourd'hui sont des molécules d'ADN double brin circulaires superenroulées. Dans certaines plantes, en plus des formes annulaires, il existe également des formes linéaires, et dans certains protozoaires, par exemple les ciliés, seul l'ADN linéaire a été trouvé dans les mitochondries.
Typiquement, chaque mitochondrie contient plusieurs copies de son génome. Ainsi, dans les cellules hépatiques humaines, il y a environ 2 000 mitochondries, et dans chacune d'elles, il y a 10 génomes identiques. Dans les fibroblastes de souris, il y a 500 mitochondries contenant deux génomes, et dans les cellules de levure S.cerevisiae- jusqu'à 22 mitochondries de quatre génomes chacune.
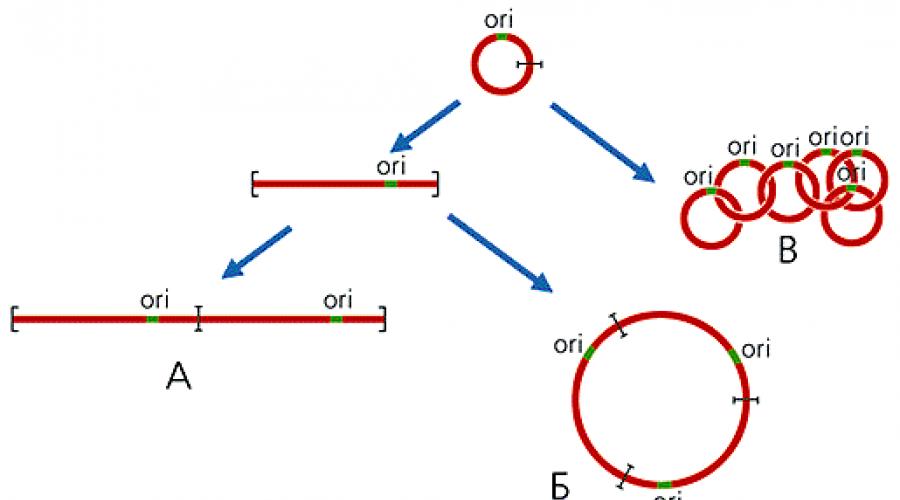
Le génome mitochondrial des plantes est généralement constitué de plusieurs molécules. Différentes tailles. L'un d'eux, le «chromosome principal», contient la plupart des gènes, et des formes annulaires de plus petite longueur, en équilibre dynamique à la fois entre elles et avec le chromosome principal, se forment à la suite d'une recombinaison intra- et intermoléculaire due à à la présence de séquences répétées (Fig. 1 ).
Fig. 1. Schéma de formation de molécules d'ADN circulaires de différentes tailles dans les mitochondries végétales.
La recombinaison se produit sur des sites répétés (indiqués en bleu).

Figue 2. Schéma de formation d'oligomères d'ADNmt linéaires (A), circulaires (B), à chaîne (C).
ori - la région d'origine de la réplication de l'ADN.
La taille du génome mitochondrial de différents organismes varie de moins de 6 000 paires de bases dans le plasmodium malarique (en plus de deux gènes d'ARNr, il ne contient que trois gènes codant pour des protéines) à des centaines de milliers de paires de bases dans Plantes terrestres(par exemple, à Arabidopsis thaliana de la famille des crucifères 366924 paires de bases). Dans le même temps, des différences de 7 à 8 fois dans les tailles d'ADNmt des plantes supérieures sont trouvées même au sein de la même famille. La longueur de l'ADNmt des vertébrés diffère légèrement: chez l'homme - 16569 paires de bases, chez les porcs - 16350, chez les dauphins - 16330, chez les grenouilles à griffes Xénope laevis- 17533, carpe - 16400. Ces génomes sont également similaires en termes de localisation des gènes, dont la plupart sont situés bout à bout ; dans certains cas, ils se chevauchent même, généralement d'un nucléotide, de sorte que le dernier nucléotide d'un gène est le premier du suivant. Contrairement aux vertébrés, chez les plantes, les champignons et les protozoaires, l'ADNmt contient jusqu'à 80 % de séquences non codantes. Dans différentes espèces, l'ordre des gènes dans les génomes des mitochondries est différent.
Une concentration élevée d'espèces réactives de l'oxygène dans les mitochondries et un système de réparation faible augmentent la fréquence des mutations de l'ADNmt par rapport au nucléaire d'un ordre de grandeur. Les radicaux oxygène provoquent des substitutions spécifiques de C®T (désamination de la cytosine) et de G®T (dommages oxydatifs à la guanine), ce qui peut entraîner une richesse de l'ADNmt en paires AT. De plus, tous les ADNmt ont une propriété intéressante - ils ne sont pas méthylés, contrairement à l'ADN nucléaire et procaryote. On sait que la méthylation (modification chimique temporaire de la séquence nucléotidique sans perturber la fonction codante de l'ADN) est l'un des mécanismes d'inactivation programmée des gènes.
Réplication et transcription de l'ADN dans les mitochondries de mammifères
Chez la plupart des animaux, les chaînes complémentaires de l'ADNmt diffèrent considérablement en termes de densité spécifique, car elles contiennent des quantités différentes de nucléotides puriques « lourds » et de nucléotides pyrimidiques « légers ». Ils sont donc appelés - chaîne H (lourde - lourde) et L (légère - légère). Au début de la réplication de la molécule d'ADNmt, la soi-disant boucle D (de la boucle de déplacement anglaise) est formée. Cette structure, visible au microscope électronique, est constituée d'un brin double et d'un brin simple (partie rétractée de la chaîne H). La région double brin est formée d'une partie de la chaîne L et d'un fragment d'ADN nouvellement synthétisé complémentaire de 450 à 650 (selon le type d'organisme) nucléotides de long, ayant une amorce ribonucléotidique à l'extrémité 5', qui correspond au point de départ de la synthèse de la chaîne H (ori H). Synthèse La chaîne L ne commence que lorsque la chaîne H fille atteint le point ori L. Cela est dû au fait que la région d'initiation de la réplication de la chaîne L est accessibles aux enzymes de synthèse d'ADN uniquement à l'état simple brin, et donc uniquement dans la double hélice non torsadée lors de la synthèse des brins H Ainsi, les brins filles d'ADNmt sont synthétisés de manière continue et asynchrone (Fig. 3).

Figue 3. Schéma de réplication de l'ADNmt chez les mammifères.
Tout d'abord, une boucle D est formée, puis un brin H fille est synthétisé,
puis la synthèse de la chaîne L fille commence.
Dans les mitochondries, le nombre total de molécules de boucle D dépasse de manière significative le nombre de molécules entièrement répliquantes. Cela est dû au fait que la boucle D a des fonctions supplémentaires - fixation de l'ADNmt à la membrane interne et initiation de la transcription, puisque les promoteurs de transcription des deux brins d'ADN sont localisés dans cette région.
Contrairement à la plupart des gènes eucaryotes, qui sont transcrits indépendamment les uns des autres, chacune des chaînes d'ADNmt de mammifère est réécrite pour former une molécule d'ARN commençant dans la région ori H. En plus de ces deux longues molécules d'ARN, complémentaires des chaînes H et L, plus de sections courtes de la chaîne H qui commencent au même point et se terminent à l'extrémité 3" du gène de l'ARNr 16S (Fig. 4). Il y a 10 fois plus de tels transcrits courts que de longs. À la suite de la maturation ( traitement), de l'ARNr 12S est formé à partir d'eux et de l'ARNr 16S impliqué dans la formation des ribosomes mitochondriaux, ainsi que des ARNt de phénylalanine et de valine. Les ARNt restants sont excisés à partir de longs transcrits et des ARNm traduits sont formés, aux extrémités 3 "dont polyadenyl les séquences sont jointes. Les extrémités 5 'de ces ARNm ne sont pas coiffées, ce qui est inhabituel pour les eucaryotes.L'épissage (fusion) ne se produit pas, car aucun des gènes mitochondriaux de mammifères ne contient d'introns.

Figue 4. Transcription de l'ADNmt humain contenant 37 gènes. Tous les transcrits commencent à être synthétisés dans la région ori H. Les ARN ribosomiques sont excisés des transcrits à chaîne H longue et courte. L'ARNt et l'ARNm sont formés à la suite du traitement à partir des transcrits des deux brins d'ADN. Les gènes d'ARNt sont représentés en vert clair.Surprises du génome mitochondrial
Malgré le fait que les génomes des mitochondries de mammifères et de levure contiennent approximativement le même nombre de gènes, la taille du génome de la levure est 4 à 5 fois plus grande - environ 80 000 paires de bases. Bien que les séquences codantes d'ADNmt de levure soient hautement homologues à celles de l'homme, les ARNm de levure ont en outre une région de tête en 5' et une région non codante en 3', comme la plupart des ARNm nucléaires. Certains gènes contiennent également des introns. Par exemple, le gène box codant pour la cytochrome oxydase b possède deux introns. Une copie de la majeure partie du premier intron est excisée de manière autocatalytique (sans la participation d'aucune protéine) du transcrit d'ARN primaire. L'ARN restant sert de matrice pour la formation de l'enzyme maturase impliquée dans l'épissage. Une partie de sa séquence d'acides aminés est codée dans les copies restantes des introns. La maturase les coupe, détruisant son propre ARNm, des copies des exons sont fusionnées et l'ARNm de la cytochrome oxydase b est formé (Fig. 5). La découverte d'un tel phénomène nous a obligés à reconsidérer le concept d'introns comme "rien ne codant des séquences". 
Figue 5. Traitement (maturation) de l'ARNm de la cytochrome oxydase b dans les mitochondries de levure.
Au premier stade de l'épissage, l'ARNm est formé, selon lequel la maturase est synthétisée,
nécessaire pour la deuxième étape de l'épissage.
Lors de l'étude de l'expression des gènes mitochondriaux Trypanosoma brucei ont trouvé un écart surprenant par rapport à l'un des axiomes de base de la biologie moléculaire, qui dit que la séquence de nucléotides dans l'ARNm correspond exactement à celle des régions codantes de l'ADN. Il s'est avéré que l'ARNm d'une des sous-unités de la cytochrome c oxydase est édité ; après transcription, sa structure primaire change - quatre uraciles sont insérés. En conséquence, un nouvel ARNm est formé, qui sert de matrice pour la synthèse d'une sous-unité supplémentaire de l'enzyme, dont la séquence d'acides aminés n'a rien à voir avec la séquence codée par l'ARNm non édité (voir tableau).

Découverte pour la première fois dans les mitochondries des trypanosomes, l'édition de l'ARN est largement répandue dans les chloroplastes et les mitochondries des plantes supérieures. Il a également été trouvé dans des cellules somatiques de mammifères, par exemple, dans l'épithélium intestinal humain, l'ARNm du gène de l'apolipoprotéine est modifié.
Les mitochondries ont présenté la plus grande surprise aux scientifiques en 1979. Jusque-là, on croyait que le code génétique était universel et que les mêmes triplets encodaient les mêmes acides aminés chez les bactéries, les virus, les champignons, les plantes et les animaux. Le chercheur anglais Burrell a comparé la structure d'un des gènes mitochondriaux du veau avec la séquence d'acides aminés dans la sous-unité de la cytochrome oxydase codée par ce gène. Il s'est avéré que le code génétique des grosses mitochondries bétail(comme une personne) n'est pas seulement différent de l'universel, il est "idéal", c'est-à-dire obéit à la règle suivante : « si deux codons ont deux nucléotides identiques, et que le troisième nucléotide appartient à la même classe (purine - A, G, ou pyrimidine - U, C), alors ils codent pour le même acide aminé ». Il existe deux exceptions à cette règle dans le code universel : le triplet AUA code pour l'isoleucine, et le codon AUG code pour la méthionine, tandis que dans le code mitochondrial idéal, ces deux triplets codent pour la méthionine ; le triplet UGG code uniquement le tryptophane, tandis que le triplet UGA code un codon stop. Dans le code universel, les deux déviations concernent les moments fondamentaux de la synthèse protéique : le codon AUG est initiateur, et le codon stop UGA stoppe la synthèse du polypeptide. Le code idéal n'est pas inhérent à toutes les mitochondries décrites, mais aucune d'entre elles n'a de code universel. On peut dire que les mitochondries parlent des langues différentes, mais jamais la langue du noyau.
Comme déjà mentionné, il existe 22 gènes d'ARNt dans le génome mitochondrial des vertébrés. Comment un tel ensemble incomplet sert-il les 60 codons pour les acides aminés (le code idéal de 64 triplets a quatre codons d'arrêt, tandis que le code universel en a trois) ? Le fait est que lors de la synthèse des protéines dans les mitochondries, les interactions codon-anticodon sont simplifiées - deux des trois nucléotides anticodon sont utilisés pour la reconnaissance. Ainsi, un ARNt reconnaît les quatre représentants de la famille des codons, qui ne diffèrent que par le troisième nucléotide. Par exemple, un ARNt de leucine avec l'anticodon GAU se dresse sur le ribosome en face des codons CUU, CUU, CUA et CUG, assurant l'inclusion sans équivoque de la leucine dans la chaîne polypeptidique. Les deux autres codons de leucine UUA et UUG sont reconnus par les ARNt avec l'anticodon AAU. Au total, huit molécules d'ARNt différentes reconnaissent huit familles de quatre codons chacune, et 14 ARNt reconnaissent différentes paires de codons, chacun codant pour un acide aminé.
Il est important que les enzymes aminoacyl-ARNt synthétase responsables de la fixation des acides aminés aux ARNt mitochondriaux correspondants soient codées dans le noyau cellulaire et synthétisées sur les ribosomes du réticulum endoplasmique. Ainsi, chez les vertébrés, tous les composants protéiques de la synthèse mitochondriale des polypeptides sont cryptés dans le noyau. Dans le même temps, la synthèse des protéines dans les mitochondries n'est pas inhibée par le cycloheximide, qui bloque le travail des ribosomes eucaryotes, mais est sensible aux antibiotiques érythromycine et chloramphénicol, qui inhibent la synthèse des protéines chez les bactéries. Ce fait est l'un des arguments en faveur de l'origine des mitochondries à partir de bactéries aérobies lors de la formation symbiotique des cellules eucaryotes.
Théorie symbiotique de l'origine des mitochondries
L'hypothèse sur l'origine des mitochondries et des plastes végétaux à partir de bactéries endosymbiontes intracellulaires a été proposée par R. Altman en 1890. Au cours du siècle de développement rapide de la biochimie, de la cytologie, de la génétique et de la biologie moléculaire apparu il y a un demi-siècle, l'hypothèse s'est développée dans une théorie basée sur une grande quantité de données factuelles. Son essence est la suivante: avec l'avènement des bactéries photosynthétiques dans l'atmosphère terrestre, l'oxygène s'est accumulé - un sous-produit de leur métabolisme. Avec une augmentation de sa concentration, la vie des hétérotrophes anaérobies est devenue plus compliquée et certains d'entre eux sont passés de la fermentation sans oxygène à la phosphorylation oxydative pour obtenir de l'énergie. De tels hétérotrophes aérobies pourraient, avec une efficacité supérieure à celle des bactéries anaérobies, décomposer les substances organiques formées à la suite de la photosynthèse. Une partie des aérobies libres a été capturée par les anaérobies, mais non "digérée", mais stockée sous forme de stations énergétiques, les mitochondries. Il ne faut pas considérer les mitochondries comme des esclaves capturés pour fournir des molécules d'ATP à des cellules incapables de respirer. Ce sont plutôt des « créatures » qui, même au Protérozoïque, ont trouvé pour elles-mêmes et leur progéniture le meilleur des abris, où elles peuvent fournir le moindre effort sans risquer d'être mangées.
De nombreux faits plaident en faveur de la théorie symbiotique :
- les tailles et formes des mitochondries et des bactéries aérobies libres coïncident ; les deux contiennent des molécules d'ADN circulaires non associées à des histones (contrairement à l'ADN nucléaire linéaire);Il y a une idée que différents royaumes d'eucaryotes avaient des ancêtres différents et que l'endosymbiose des bactéries est apparue à différents stades de l'évolution des organismes vivants. Ceci est également mis en évidence par des différences dans la structure des génomes mitochondriaux des protozoaires, des champignons, des plantes et des animaux supérieurs. Mais dans tous les cas, la majeure partie des gènes des promitochondries est entrée dans le noyau, éventuellement à l'aide d'éléments génétiques mobiles. Lorsqu'une partie du génome de l'un des symbiotes est incluse dans le génome de l'autre, l'intégration des symbiotes devient irréversible.Les ARN ribosomiques et de transfert des mitochondries diffèrent par les séquences nucléotidiques des ARN nucléaires, tout en démontrant une similitude surprenante avec des molécules similaires de certaines eubactéries aérobies à Gram négatif ;
Les ARN polymérases mitochondriales, bien que codées dans le noyau cellulaire, sont inhibées par la rifampicine, tout comme les bactériennes, et les ARN polymérases eucaryotes sont insensibles à cet antibiotique ;
La synthèse des protéines dans les mitochondries et les bactéries est inhibée par les mêmes antibiotiques qui n'affectent pas les ribosomes eucaryotes ;
La composition lipidique de la membrane mitochondriale interne et du plasmalemme bactérien est similaire, mais très différente de celle de la membrane mitochondriale externe, qui est homologue aux autres membranes des cellules eucaryotes ;
Les crêtes formées par la membrane mitochondriale interne sont des analogues évolutifs des membranes mésosomales de nombreux procaryotes ;
Jusqu'à présent, des organismes ont survécu qui imitent des formes intermédiaires sur la voie de la formation de mitochondries à partir de bactéries (amibe primitive Pelomyxa n'a pas de mitochondries, mais contient toujours des bactéries endosymbiotiques).
Le nouveau génome peut créer des voies métaboliques menant à la formation produits utiles, qui ne peut être synthétisé par aucun des partenaires séparément. Ainsi, la synthèse d'hormones stéroïdiennes par les cellules du cortex surrénalien est une chaîne complexe de réactions, dont certaines se produisent dans les mitochondries, et d'autres dans le réticulum endoplasmique. Après avoir capturé les gènes des promitochondries, le noyau a pu contrôler de manière fiable les fonctions du symbiote. Le noyau code pour toutes les protéines et la synthèse lipidique de la membrane externe des mitochondries, la plupart des protéines de la matrice et la membrane interne des organites. Plus important encore, le noyau code pour les enzymes de réplication, de transcription et de traduction de l'ADNmt, contrôlant ainsi la croissance et la reproduction des mitochondries. Le taux de croissance des partenaires en symbiose devrait être approximativement le même. Si l'hôte se développe plus rapidement, à chaque génération, le nombre de symbiotes par individu diminuera et, à la fin, des descendants apparaîtront sans mitochondries. Nous savons que chaque cellule d'un organisme à reproduction sexuée contient de nombreuses mitochondries qui répliquent leur ADN entre les divisions hôtes. Cela garantit que chacune des cellules filles reçoit au moins une copie du génome mitochondrial.
Héritage cytoplasmique
En plus de coder les composants clés de la chaîne respiratoire et son propre appareil de synthèse des protéines, le génome mitochondrial, dans certains cas, est impliqué dans la formation de certains traits morphologiques et physiologiques. Ces caractéristiques comprennent le syndrome NCS (rayure non chromosomique, tache foliaire non codée par chromosome) et la stérilité mâle cytoplasmique (CMS), qui entraîne une perturbation du développement normal du pollen, caractéristique d'un certain nombre d'espèces végétales supérieures. La manifestation des deux caractéristiques est due à des changements dans la structure de l'ADNmt. Dans le CMS, des réarrangements des génomes mitochondriaux sont observés à la suite d'événements de recombinaison conduisant à des délétions, des duplications, des inversions ou des insertions de certaines séquences nucléotidiques ou de gènes entiers. De tels changements peuvent endommager non seulement les gènes existants, mais aussi l'émergence de nouveaux gènes fonctionnels.
La transmission cytoplasmique, contrairement à la transmission nucléaire, n'obéit pas aux lois de Mendel. Cela est dû au fait que chez les animaux et les plantes supérieurs, les gamètes de sexes différents contiennent des quantités disparates de mitochondries. Ainsi, dans l'œuf de souris, il y a 90 000 mitochondries et dans le sperme - seulement quatre. Évidemment, dans un œuf fécondé, les mitochondries proviennent principalement ou uniquement de femelle, c'est à dire. l'héritage de tous les gènes mitochondriaux est maternel. L'analyse génétique de l'hérédité cytoplasmique est difficile en raison des interactions nucléaire-cytoplasmique. Dans le cas de la stérilité mâle cytoplasmique, le génome mitochondrial mutant interagit avec certains gènes nucléaires dont les allèles récessifs sont nécessaires au développement du trait. Les allèles dominants de ces gènes, tant à l'état homozygote qu'hétérozygote, restaurent la fertilité des plantes, quel que soit l'état du génome mitochondrial.
L'étude des génomes mitochondriaux, leur évolution, suivant les lois spécifiques de la génétique des populations, la relation entre les systèmes génétiques nucléaire et mitochondrial, est nécessaire pour comprendre l'organisation hiérarchique complexe de la cellule eucaryote et de l'organisme dans son ensemble.
Certaines maladies héréditaires et le vieillissement humain sont associés à certaines mutations de l'ADN mitochondrial ou de gènes nucléaires qui contrôlent le fonctionnement des mitochondries. Les données s'accumulent sur l'implication des défauts de l'ADNmt dans la cancérogenèse. Par conséquent, les mitochondries peuvent être une cible pour la chimiothérapie anticancéreuse. Il existe des faits sur l'interaction étroite des génomes nucléaires et mitochondriaux dans le développement d'un certain nombre de pathologies humaines. De multiples délétions d'ADNmt ont été trouvées chez des patients présentant une faiblesse musculaire sévère, une ataxie, une surdité, un retard mental, hérités de manière autosomique dominante. Le dimorphisme sexuel a été établi dans les manifestations cliniques de la maladie coronarienne, ce qui est très probablement dû à l'effet maternel - l'hérédité cytoplasmique. Le développement de la thérapie génique offre l'espoir de corriger les défauts des génomes mitochondriaux dans un avenir prévisible.
Ce travail a été soutenu par la Fondation russe pour la recherche fondamentale. Projet 01-04-48971.
L'auteur est reconnaissant à l'étudiant diplômé M.K. Ivanov, qui a créé les dessins de l'article.
Littérature
1. Yankovsky N.K., Borinskaya S.A. Notre histoire écrite dans l'ADN // Nature. 2001. N° 6. pp.10-18. 2. Minchenko AG, Dudareva N.A. Génome mitochondrial. Novossibirsk, 1990. 3. Gvozdev V.A.// Soro. éduquer. magazine 1999. N° 10. pp.11-17. 4. Margelis L. Le rôle de la symbiose dans l'évolution cellulaire. M., 1983. 5. Skoulatchev V.P.// Soro. éduquer. magazine 1998. N° 8. S.2-7. 6. Igamberdiev A.U.// Soro. éduquer. magazine 2000. N° 1. S.32-36.
La structure des acides nucléiques.
N. to. - sont une macromolécule informationnelle universelle de la cellule, dont la fonction principale est: 1) Stockage informations héréditaires sous la forme d'un code génétique. 2) Reproduction d'informations héréditaires par auto-duplication ou réplication de l'ADN. 3) Mise en œuvre de l'information héréditaire dans le processus de biosynthèse des protéines. N to étaient reçus pour la première fois des noyaux des carreaux du pus. Analyse chimique ont montré qu'il existe deux types de n à : 1) ADN, 2) ARN. L'ADN se trouve dans le noyau, les mitochondries et les centrioles. Les ARN se trouvent dans le noyau, les nucléoles, les ribosomes et les mitochondries. En termes chimiques, n to - polymères constitués de chaînes polynucléotidiques, monomère n. yav - nucléotide. Il existe 4 types d'ADN : A, T, G, C. Dans l'ARN, la thymine est remplacée par l'uracile. Selon la structure, l'adénine et la guanine sont des bases puriques
Propriétés et fonctions de l'ADN.
L'analyse chimique a montré qu'il existe deux types de n à : 1) ADN, 2) ARN. L'ADN se trouve dans le noyau, les mitochondries et les centrioles. Les ARN se trouvent dans le noyau, les nucléoles, les ribosomes et les mitochondries. En termes chimiques, n to - polymères constitués de chaînes polynucléotidiques, monomère n. yav - nucléotide. Il existe 4 types d'ADN : A, T, G, C. Dans l'ARN, la thymine est remplacée par l'uracile. Selon la structure, l'adénine et la guanine sont des bases puriques
1 noyau benzénique) T, C, U (bases pyrimidiques - 2 noyaux benzéniques). L'ADN est une hélice constituée de deux chaînes polynucléotidiques. Les chaînes polynucléotidiques sont constituées de nucléotides reliés entre eux par des liaisons hydrogène qui forment des paires complémentaires. Dans l'ADN A=T, G=C (Règle de Chargaff 1951). Les nucléotides de chaque brin d'ADN sont connectés les uns aux autres de sorte que le 5e carbone du sucre du nucléotide précédent est connecté au 3e carbone du sucre du suivant. En raison de ces liaisons, la molécule d'ADN a deux extrémités. Le diamètre de l'hélice LNC est de 2 nm. Un tour d'hélice correspond à 3,4 nm. La distance entre les nucléotides est de 0,34 nm. Chaque tour de l'hélice contient 10 paires de bases. Il existe plusieurs niveaux d'organisation dans l'ADN : 1) La structure primaire est l'ordre dans lequel les nucléotides sont disposés en chaînes complémentaires. 2) La structure secondaire est la double hélice de l'ADN. 3) Tertiaire - ADN dans la composition des chromosomes.
Caractéristiques structurelles de l'ADN mitochondrial.
ADN mitochondrial (ADNmt) - ADN localisé (contrairement à l'ADN nucléaire) dans les mitochondries, organelles des cellules eucaryotes.
Dans la plupart des organismes étudiés, les mitochondries ne contiennent que des molécules d'ADN circulaires, dans certaines plantes, des molécules circulaires et linéaires sont présentes simultanément, et dans un certain nombre de protistes (par exemple, les ciliés), il n'y a que des molécules linéaires. Les mitochondries de mammifères contiennent généralement deux à dix copies identiques de molécules d'ADN circulaires. Chez les plantes, chaque mitochondrie contient plusieurs molécules d'ADN de tailles différentes qui sont capables de se recombiner.Chez les protistes de l'ordre des kinétoplastides (par exemple, les trypanosomes), une section spéciale de la mitochondrie (cinétoplaste) contient deux types de molécules d'ADN - identiques maxi- anneaux (20-50 pièces) d'environ 21 ko de long. et mini-anneaux (20 000 à 55 000 pièces, environ 300 variétés, longueur moyenne d'environ 1 000 pb). Tous les anneaux sont connectés en un seul réseau (catenans), qui est détruit et restauré à chaque cycle de réplication. Les maxi-anneaux sont homologues à l'ADN mitochondrial d'autres organismes. Chaque mini-anneau contient quatre régions conservées similaires et quatre régions hypervariables uniques. Les minicercles codent pour de courtes molécules d'ARN guide (guideRNA) qui modifient l'ARN transcrit à partir des gènes des maxicercles. L'ADN mitochondrial (ADNmt) est le génome des organites cellulaires - les mitochondries. L'origine endosymbiotique de ces organites détermine l'existence semi-autonome du système génétique mitochondrial. Ainsi, la synthèse d'ADN dans les mitochondries se déroule indépendamment de la synthèse d'ADN nucléaire, et l'héritage de cette structure génétique cytoplasmique - le chromosome mitochondrial - se produit normalement strictement le long de la lignée maternelle. Cela donne aux auteurs une raison d'isoler conditionnellement la totalité des gènes mitochondriaux et tous les fragments d'ADNmt en cours de réplication dans une ressource génétique distincte de la population - le pool de gènes mitochondriaux. Des structures contenant de l'ADN dans les mitochondries ont été identifiées dans les années 60. Au cours du dernier quart de siècle, l'organisation structurelle et fonctionnelle du génome mitochondrial de l'homme et de nombreuses espèces animales a été étudiée en détail. Le chromosome mitochondrial est représenté par une molécule d'ADN double brin circulaire, qui est présente dans l'organite sous la forme d'une forme superenroulée fermée de manière covalente associée à la membrane mitochondriale interne. Chaque organite contient 1 à 8 molécules d'ADN, soit 1000 à 8000 copies par cellule. En règle générale, un organisme possède une seule forme d'ADNmt, c'est-à-dire un haplotype hérité de la lignée maternelle.
Types d'ARN dans les cellules.
Il existe trois types d'ARN dans les cellules : 1) ARN-I (messager ou ARN messager).
2) ARN-R (ARN ribosomal).
3) ARN-T (ARN de transfert)
ARN messager - synthétisé et transcrit en ADN et transporte des informations pour la synthèse des protéines. L'ARN-R et l'ARN-T sont synthétisés dans les nucléoles du noyau. Le nucléole est une région de chromosomes qui possède des satellites. L'ADN nucléolaire contient les gènes sur lesquels l'ARN-R et l'ARN-T sont synthétisés. Les ARNr se trouvent dans les ribosomes (dans la petite et la grande sous-unité). But : à travers la petite sous-unité d'AK, il se fixe à l'ARN-T via l'ATP. La différence entre l'ADN et l'ARN : 1) L'ARN est constitué d'une seule chaîne. 2) Dans l'ARN, le sucre est le ribose. 3) L'ARN est plus court que l'ADN. 4) L'ARN-T a la forme d'une structure tertiaire. ARN matriciel (informationnel) - ARN, qui sert d'intermédiaire dans le transfert d'informations codées dans l'ADN aux ribosomes, machines moléculaires qui synthétisent les protéines d'un organisme vivant. La séquence codante de l'ARNm détermine la séquence d'acides aminés de la chaîne polypeptidique d'une protéine.
Transport (ARNt) - petit, composé d'environ 80 nucléotides, molécules à structure tertiaire conservatrice. Ils transportent des acides aminés spécifiques vers le site de synthèse des liaisons peptidiques dans le ribosome. Chaque ARNt contient un site de fixation d'acides aminés et un anticodon pour la reconnaissance et la fixation aux codons d'ARNm. L'ARN ribosomique (ARNr) est le composant catalytique des ribosomes. Les ribosomes eucaryotes contiennent quatre types de molécules d'ARNr : 18S, 5.8S, 28S et 5S. Trois de quatre types L'ARNr est synthétisé dans le nucléole. Dans le cytoplasme, les ARN ribosomiques se combinent avec des protéines ribosomiques pour former une nucléoprotéine appelée ribosome. Le ribosome se fixe à l'ARNm et synthétise la protéine. L'ARNr représente jusqu'à 80% de l'ARN présent dans le cytoplasme d'une cellule eucaryote
Introduction
Un quart de siècle s'est écoulé depuis la découverte des molécules d'ADN dans les mitochondries avant qu'ils ne s'intéressent non seulement aux biologistes moléculaires et aux cytologistes, mais aussi à la génétique, aux évolutionnistes, ainsi qu'aux paléontologues et aux médecins légistes. Un tel intérêt a été suscité par les travaux d'A. Wilson de l'Université de Californie. En 1987, il publie les résultats d'une analyse comparative de l'ADN mitochondrial prélevé sur 147 représentants de différents groupes ethniques de toutes les races humaines habitant les cinq continents. Selon le type, l'emplacement et le nombre de mutations individuelles, il a été établi que tout l'ADN mitochondrial provenait d'une séquence nucléotidique ancestrale par divergence. Dans la presse pseudo-scientifique, cette conclusion a été interprétée de manière extrêmement simplifiée - toute l'humanité provenait d'une femme, appelée Eve mitochondriale (car les filles et les fils ne reçoivent des mitochondries que de leur mère), qui vivait en Afrique du Nord-Est il y a environ 200 000 ans . Après encore 10 ans, il a été possible de déchiffrer un fragment d'ADN mitochondrial isolé des restes d'un Néandertalien et d'estimer le temps d'existence du dernier ancêtre commun de l'homme et de Néandertal à 500 000 ans.
Aujourd'hui, la génétique mitochondriale humaine se développe intensivement tant dans la population que dans le domaine médical. Un lien a été établi entre un certain nombre de maladies héréditaires graves et des défauts de l'ADN mitochondrial. Les changements génétiques associés au vieillissement sont plus prononcés dans les mitochondries. Qu'est-ce que le génome mitochondrial, qui diffère chez l'homme et les autres animaux de celui des plantes, des champignons et des protozoaires en termes de taille, de forme et de capacité génétique ? Quel est le rôle, comment fonctionne-t-il et comment le génome mitochondrial est-il né dans différents taxons en général et chez l'homme en particulier ? À ce sujet et sera discuté dans mon résumé "petit et très modeste".
En plus de l'ADN, la matrice mitochondriale contient ses propres ribosomes, qui diffèrent par de nombreuses caractéristiques des ribosomes eucaryotes situés sur les membranes du réticulum endoplasmique. Cependant, les ribosomes mitochondriaux ne forment pas plus de 5% de toutes les protéines qui les composent. La plupart des protéines qui composent les composants structurels et fonctionnels des mitochondries sont codées par le génome nucléaire, synthétisées sur les ribosomes du réticulum endoplasmique et transportées par ses canaux jusqu'au site d'assemblage. Ainsi, les mitochondries sont le résultat des efforts conjugués de deux génomes et de deux appareils de transcription et de traduction. Certaines enzymes sous-unités de la chaîne respiratoire mitochondriale sont constituées de différents polypeptides, dont certains sont codés par le nucléaire et d'autres par le génome mitochondrial. Par exemple, l'enzyme clé de la phosphorylation oxydative, la cytochrome c oxydase, chez la levure se compose de trois sous-unités codées et synthétisées dans les mitochondries et de quatre sous-unités codées dans le noyau cellulaire et synthétisées dans le cytoplasme. L'expression de la plupart des gènes mitochondriaux est contrôlée par certains gènes nucléaires.
Théorie symbiotique de l'origine des mitochondries
L'hypothèse sur l'origine des mitochondries et des plastes végétaux à partir de bactéries endosymbiontes intracellulaires a été avancée par R. Altman en 1890. Au cours du siècle de développement rapide de la biochimie, de la cytologie, de la génétique et de la biologie moléculaire apparu il y a un demi-siècle, l'hypothèse s'est transformé en une théorie basée sur des éléments plus factuels. Son essence est la suivante: avec l'avènement des bactéries photosynthétiques, l'oxygène s'est accumulé dans l'atmosphère terrestre - un sous-produit de leur métabolisme. Avec une augmentation de sa concentration, la vie des hétérotrophes anaérobies s'est compliquée et certains d'entre eux sont passés de la fermentation anoxique à la phosphorylation oxydative pour obtenir de l'énergie. De tels hétérotrophes aérobies pourraient, avec une efficacité supérieure à celle des bactéries anaérobies, décomposer les substances organiques formées à la suite de la photosynthèse. Une partie des aérobies libres a été capturée par les anaérobies, mais non «digérée», mais stockée sous forme de stations énergétiques, les mitochondries. Il ne faut pas considérer les mitochondries comme des esclaves capturés pour fournir des molécules d'ATP à des cellules incapables de respirer. Ce sont plutôt des « créatures » qui, même au Protérozoïque, ont trouvé pour elles-mêmes et leur progéniture le meilleur des abris, où elles peuvent fournir le moindre effort sans risquer d'être mangées.
De nombreux faits plaident en faveur de la théorie symbiotique :
Les tailles et les formes des mitochondries et des bactéries aérobies libres coïncident ; les deux contiennent des molécules d'ADN circulaires non associées à des histones (contrairement à l'ADN nucléaire linéaire);
Les ARN ribosomiques et de transfert des mitochondries diffèrent par les séquences nucléotidiques des ARN nucléaires, tout en démontrant une similitude surprenante avec des molécules similaires de certaines eubactéries aérobies à Gram négatif ;
Les ARN polymérases mitochondriales, bien que codées dans le noyau cellulaire, sont inhibées par la rifampicine, tout comme les bactériennes, et les ARN polymérases eucaryotes sont insensibles à cet antibiotique ;
La synthèse des protéines dans les mitochondries et les bactéries est inhibée par les mêmes antibiotiques qui n'affectent pas les ribosomes eucaryotes ;
La composition lipidique de la membrane mitochondriale interne et du plasmalemme bactérien est similaire, mais très différente de celle de la membrane mitochondriale externe, qui est homologue aux autres membranes des cellules eucaryotes ;
Les crêtes formées par la membrane mitochondriale interne sont des analogues évolutifs des membranes mésosomales de nombreux procaryotes ;
Jusqu'à présent, des organismes ont survécu qui imitent des formes intermédiaires sur la voie de la formation de mitochondries à partir de bactéries (amibe primitive Pelomyxa n'a pas de mitochondries, mais contient toujours des bactéries endosymbiotiques).
Il y a une idée que différents royaumes d'eucaryotes avaient des ancêtres différents et que l'endosymbiose des bactéries est apparue à différents stades de l'évolution des organismes vivants. Ceci est également mis en évidence par des différences dans la structure des génomes mitochondriaux des protozoaires, des champignons, des plantes et des animaux supérieurs. Mais dans tous les cas, la majeure partie des gènes des promitochondries est entrée dans le noyau, éventuellement à l'aide d'éléments génétiques mobiles. Lorsqu'une partie du génome de l'un des symbiotes est incluse dans le génome de l'autre, l'intégration des symbiotes devient irréversible. Le nouveau génome peut créer des voies métaboliques conduisant à des produits utiles qui ne peuvent être synthétisés par l'un ou l'autre des partenaires seuls. Ainsi, la synthèse des hormones stéroïdes par les cellules du cortex surrénalien est une chaîne complexe de réactions, dont certaines se produisent dans les mitochondries, et d'autres dans le réticulum endoplasmique. Après avoir capturé les gènes des promitochondries, le noyau a pu contrôler de manière fiable les fonctions du symbiote. Le noyau code pour toutes les protéines et la synthèse lipidique de la membrane externe des mitochondries, la plupart des protéines de la matrice et la membrane interne des organites. Plus important encore, le noyau code pour les enzymes de réplication, de transcription et de traduction de l'ADNmt, contrôlant ainsi la croissance et la reproduction des mitochondries. Le taux de croissance des partenaires en symbiose devrait être approximativement le même. Si l'hôte se développe plus rapidement, à chaque génération, le nombre de symbiotes par individu diminuera et, à la fin, des descendants apparaîtront sans mitochondries. Nous savons que chaque cellule d'un organisme à reproduction sexuée contient de nombreuses mitochondries qui répliquent leur ADN entre les divisions hôtes. Cela garantit que chacune des cellules filles reçoit au moins une copie du génome mitochondrial.
Rôle du noyau cellulaire dans la biogenèse mitochondriale
La levure mutante d'un certain type présente une grande délétion dans l'ADN mitochondrial, ce qui conduit à un arrêt complet de synthèse des protéines dans les mitochondries ; en conséquence, ces organites ne sont pas en mesure d'accomplir leur fonction. Étant donné que ces mutants forment de petites colonies lorsqu'ils sont cultivés sur un milieu à faible teneur en glucose, ils sont appelés mu cytoplasmiquetantamimenue.
Bien que les petits mutants manquent de synthèse des protéines mitochondriales et ne forment donc pas de mitochondries normales, ces mutants contiennent néanmoins les promitochondries, qui, dans une certaine mesure, ressemblent aux mitochondries ordinaires, ont une membrane externe normale et une membrane interne avec des crêtes peu développées. Les promitochondries contiennent de nombreuses enzymes codées par des gènes nucléaires et synthétisées sur des ribosomes cytoplasmiques, notamment des ADN et des ARN polymérases, toutes les enzymes du cycle de l'acide citrique et de nombreuses protéines qui composent la membrane interne. Ceci démontre clairement le rôle prédominant du génome nucléaire dans la biogenèse mitochondriale.
Il est intéressant de noter que bien que les fragments d'ADN perdus représentent de 20 à plus de 99,9 % du génome mitochondrial, la quantité totale d'ADN mitochondrial chez les petits mutants reste toujours au même niveau que chez le type sauvage. Cela est dû au processus encore peu étudié d'amplification de l'ADN, qui aboutit à la formation d'une molécule d'ADN constituée de répétitions en tandem d'un même site et de taille égale à une molécule normale. Par exemple, l'ADN mitochondrial d'un petit mutant qui conserve 50 % de la séquence nucléotidique de l'ADN de type sauvage sera composé de deux répétitions, tandis qu'une molécule qui ne conserve que 0,1% le génome de type sauvage sera construit à partir de 1000 copies du fragment restant. Ainsi, les petits mutants peuvent être utilisés pour produire en grand nombre certaines sections d'ADN mitochondrial, qui, pourrait-on dire, sont clonées par la nature elle-même.
Bien que la biogenèse des organites soit principalement contrôlée par des gènes nucléaires, les organites eux-mêmes, à en juger par certaines données, exercent également une sorte d'effet de rétroaction réglementaire; c'est en tout cas le cas des mitochondries. Si la synthèse des protéines est bloquée dans les mitochondries des cellules intactes, les enzymes impliquées dans la synthèse mitochondriale de l'ADN, de l'ARN et des protéines commencent à se former en excès dans le cytoplasme, comme si la cellule essayait de surmonter l'effet de l'agent bloquant. Mais, bien que l'existence d'une sorte de signal provenant des mitochondries ne fasse aucun doute, sa nature n'est toujours pas connue.
Pour un certain nombre de raisons, les mécanismes de la biogenèse mitochondriale sont aujourd'hui étudiés le plus souvent en culture. Saccharomyces carlsbergensis(levure de bière et S. cerevisiae(La levure de boulanger). Tout d'abord, lorsqu'elles poussent sur glucose, ces levures présentent une capacité unique à n'exister que grâce à la glycolyse, c'est-à-dire à se passer de la fonction des mitochondries. Cela permet d'étudier les mutations de l'ADN mitochondrial et nucléaire qui empêchent le développement de ces organites. De telles mutations sont mortelles dans presque tous les autres organismes. Deuxièmement, la levure - de simples eucaryotes unicellulaires - est facile à cultiver et sujette à des recherches biochimiques. Enfin, la levure peut se reproduire à la fois en phases haploïde et diploïde, généralement de manière asexuée par bourgeonnement (mitose asymétrique). Mais chez la levure, il y a aussi un processus sexuel : de temps en temps, deux cellules haploïdes fusionnent, formant un zygote diploïde, qui soit se divise par mitose, soit subit une méiose et donne à nouveau naissance à des cellules haploïdes. En contrôlant l'alternance de la reproduction asexuée et sexuée au cours de l'expérience, on peut en apprendre beaucoup sur les gènes responsables de la fonction des mitochondries. Grâce à ces méthodes, on peut notamment savoir si de tels gènes sont localisés dans l'ADN nucléaire ou dans l'ADN mitochondrial, puisque les mutations dans les gènes mitochondriaux ne sont pas héritées selon les lois de Mendel, qui régissent l'hérédité des gènes nucléaires.
Systèmes de transport mitochondrial
La plupart des protéines contenues dans les mitochondries et les chloroplastes sont importées dans ces organites à partir du cytosol. Cela soulève deux questions : comment la cellule dirige-t-elle les protéines vers l'organite approprié, et comment ces protéines y pénètrent-elles ?
Une réponse partielle a été obtenue en étudiant le transport de la petite sous-unité (S) de l'enzyme dans le stroma du chloroplaste ribulose-1,5-bisphosphate-carboxytrous d'homme. Si l'ARNm isolé du cytoplasme d'une algue unicellulaire Chlamydomonas ou à partir de feuilles de pois, introduites comme matrice dans un système de synthèse de protéines in vitro, alors l'une des nombreuses protéines résultantes se liera à un anticorps anti-S spécifique. La protéine S synthétisée in vitro est appelée ppo-S car elle est plus grande que la protéine S normale d'environ 50 résidus d'acides aminés. Lors de l'incubation de la protéine pro-S avec des chloroplastes intacts, elle pénètre dans les organites et y est transformée sous l'action de la peptidase en protéine S. Ensuite, la protéine S se lie à la grande sous-unité de la ribulose-1,5-bisphosphate carboxylase, synthétisée sur les ribosomes du chloroplaste, et forme avec elle une enzyme active dans le stroma du chloroplaste.
Le mécanisme de transfert de la protéine S est inconnu. On pense que le pro-S se lie à une protéine réceptrice située sur la membrane externe du chloroplaste ou à la jonction des membranes externe et interne, puis est transféré au stroma par des canaux transmembranaires dans un processus consommateur d'énergie.
De même, les protéines sont transportées dans les mitochondries. Si des mitochondries de levure purifiées sont incubées avec un extrait cellulaire contenant des protéines de levure radioactives fraîchement synthétisées, on peut observer que les protéines mitochondriales codées par le génome nucléaire sont séparées des protéines cytoplasmiques non mitochondriales et incorporées sélectivement dans les mitochondries, tout comme cela se produit dans un génome intact. cellule. Dans le même temps, les protéines des membranes externe et interne, de la matrice et de l'espace intermembranaire se dirigent vers le compartiment mitochondrial correspondant.
De nombreuses protéines nouvellement synthétisées destinées à la membrane interne, à la matrice et à l'espace intermembranaire ont un peptide leader à leur extrémité N-terminale, qui est clivée pendant le transport par une protéase spécifique située dans la matrice. Le transfert des protéines vers ces trois compartiments mitochondriaux nécessite l'énergie d'un gradient électrochimique de protons créé à travers la membrane interne. Le mécanisme de transport des protéines pour la membrane externe est différent : dans ce cas, ni dépense énergétique ni clivage protéolytique de la protéine précurseur la plus longue ne sont nécessaires. Ces observations et d'autres suggèrent que les quatre groupes de protéines mitochondriales sont transportés dans l'organelle par le mécanisme suivant : on suppose que toutes les protéines, à l'exception de celles destinées à la membrane externe, sont incluses dans la membrane interne à la suite d'une énergie- processus consommateur qui se produit aux points de contact entre les membranes externe et interne. Apparemment, après cette première incorporation de la protéine dans la membrane, celle-ci subit un clivage protéolytique, ce qui entraîne une modification de sa conformation ; selon l'évolution de la conformation, la protéine est soit fixée dans la membrane, soit « poussée » dans la matrice ou dans l'espace intermembranaire.
Le transfert des protéines à travers les membranes des mitochondries et des chloroplastes est en principe similaire à leur transfert à travers les membranes du réticulum endoplasmique. Cependant, il existe plusieurs différences importantes ici. Premièrement, lorsqu'elle est transportée dans la matrice ou le stroma, la protéine traverse à la fois les membranes externe et interne de l'organite, tandis que lorsqu'elle est transférée dans la lumière du réticulum endoplasmique, les molécules ne traversent qu'une seule membrane. De plus, le transfert des protéines dans le réticulum s'effectue selon le mécanisme excrétion dirigée(décharge vectorielle) - elle commence lorsque la protéine n'est pas encore complètement descendue du ribosome (import cotraductionnel), et le transfert vers les mitochondries et les chloroplastes se produit après que la synthèse de la molécule de protéine est complètement terminée (importation post-traductionnelle).
Malgré ces différences, dans les deux cas, la cellule synthétise des protéines précurseurs contenant une séquence signal qui détermine à quelle membrane cette protéine ira. Apparemment, dans de nombreux cas, cette séquence est clivée de la molécule précurseur après l'achèvement du processus de transport. Cependant, certaines protéines sont immédiatement synthétisées sous leur forme finale. On pense que dans de tels cas, la séquence signal est contenue dans la chaîne polypeptidique de la protéine finie. Les séquences signal sont encore mal comprises, mais il devrait probablement exister plusieurs types de telles séquences, dont chacune détermine le transfert d'une molécule protéique vers une zone spécifique de la cellule. Par exemple, dans une cellule végétale, certaines des protéines dont la synthèse commence dans le cytosol sont ensuite transportées vers les mitochondries, d'autres vers les chloroplastes, d'autres vers les peroxysomes, d'autres encore vers le réticulum endoplasmique. Les processus complexes menant à la distribution intracellulaire correcte des protéines commencent seulement à se préciser.
En dehors de acides nucléiques et les protéines, les lipides sont nécessaires pour construire de nouvelles mitochondries. Contrairement aux chloroplastes, les mitochondries reçoivent la plupart de leurs lipides de l'extérieur. Dans les cellules animales, les phospholipides synthétisés dans le réticulum endoplasmique sont transportés vers la membrane mitochondriale externe par des protéines spécifiques puis incorporés dans la membrane interne ; on pense que cela se produit au point de contact entre les deux membranes. La principale réaction de la biosynthèse des lipides, catalysée par les mitochondries elles-mêmes, est la conversion de l'acide phosphatidique en phospholipide cardiolipine, qui est contenu principalement dans la membrane mitochondriale interne et représente environ 20 % de tous ses lipides.
Taille et forme des génomes mitochondriaux
À ce jour, plus de 100 génomes mitochondriaux différents ont été lus. L'ensemble et le nombre de leurs gènes dans l'ADN mitochondrial, dont la séquence nucléotidique est complètement déterminée, varient considérablement selon les espèces d'animaux, de plantes, de champignons et de protozoaires. Le plus grand nombre de gènes a été trouvé dans le génome mitochondrial du protozoaire flagellé Rectinomo-nas americana- 97 gènes, y compris tous les gènes codant pour les protéines trouvés dans l'ADNmt d'autres organismes. Chez la plupart des animaux supérieurs, le génome mitochondrial contient 37 gènes : 13 pour les protéines de la chaîne respiratoire, 22 pour l'ARNt et deux pour l'ARNr (pour la grande sous-unité ribosomale ARNr 16S et pour le petit ARNr 12S). Chez les plantes et les protozoaires, contrairement aux animaux et à la plupart des champignons, le génome mitochondrial code également certaines protéines qui composent les ribosomes de ces organites. Les enzymes clés de la synthèse des polynucléotides matrices, telles que l'ADN polymérase (répliquant l'ADN mitochondrial) et l'ARN polymérase (transcrivant le génome mitochondrial), sont codées dans le noyau et synthétisées sur les ribosomes cytoplasmiques. Ce fait indique l'autonomie relative des mitochondries dans la hiérarchie complexe de la cellule eucaryote.
Les génomes mitochondriaux de différentes espèces diffèrent non seulement par l'ensemble des gènes, l'ordre de leur localisation et de leur expression, mais aussi par la taille et la forme de l'ADN. La grande majorité des génomes mitochondriaux décrits aujourd'hui sont des molécules d'ADN double brin circulaires superenroulées. Dans certaines plantes, en plus des formes annulaires, il existe également des formes linéaires, et dans certains protozoaires, par exemple les ciliés, seul l'ADN linéaire a été trouvé dans les mitochondries.
En règle générale, chaque mitochondrie contient plusieurs copies de son génome. Ainsi, dans les cellules hépatiques humaines, il y a environ 2 000 mitochondries, et dans chacune d'elles, il y a 10 génomes identiques. Dans les fibroblastes de souris, il y a 500 mitochondries contenant chacune deux génomes, et dans les cellules de levure S.cerevisiae- jusqu'à 22 mitochondries de quatre génomes chacune.
DIV_ADBLOCK764">
 Figue 2. Schéma de formation d'oligomères d'ADNmt linéaires (A), circulaires (B), à chaîne (C). ori - la région d'origine de la réplication de l'ADN.
Figue 2. Schéma de formation d'oligomères d'ADNmt linéaires (A), circulaires (B), à chaîne (C). ori - la région d'origine de la réplication de l'ADN.
La taille du génome mitochondrial de différents organismes varie de moins de 6 000 paires de bases dans le plasmodium malarique (en plus de deux gènes d'ARNr, il ne contient que trois gènes codant pour des protéines) à des centaines de milliers de paires de bases chez les plantes terrestres (par exemple , dans Arabidopsis thaliana de la famille des crucifères 366924 paires de bases). Dans le même temps, des différences de 7 à 8 fois dans les tailles d'ADNmt des plantes supérieures sont trouvées même au sein de la même famille. La longueur de l'ADNmt des vertébrés diffère légèrement: chez l'homme - 16569 paires de bases, chez les porcs - 16350, chez les dauphins - 16330, chez les grenouilles à griffes Xénope laevis- 17533, carpe - 16400. Ces génomes sont également similaires en termes de localisation des gènes, dont la plupart sont situés bout à bout ; dans certains cas, ils se chevauchent même, généralement d'un nucléotide, de sorte que le dernier nucléotide d'un gène est le premier du suivant. Contrairement aux vertébrés, chez les plantes, les champignons et les protozoaires, l'ADNmt contient jusqu'à 80 % de séquences non codantes. Dans différentes espèces, l'ordre des gènes dans les génomes des mitochondries est différent.
Une concentration élevée d'espèces réactives de l'oxygène dans les mitochondries et un système de réparation faible augmentent la fréquence des mutations de l'ADNmt par rapport au nucléaire d'un ordre de grandeur. Les radicaux oxygène provoquent des substitutions spécifiques de C®T (désamination de la cytosine) et de G®T (dommages oxydatifs à la guanine), à la suite de quoi, peut-être, l'ADNmt est riche en paires AT. De plus, tous les ADNmt ont une propriété intéressante - ils ne sont pas méthylés, contrairement à l'ADN nucléaire et procaryote. On sait que la méthylation (modification chimique temporaire de la séquence nucléotidique sans violer la fonction codante de l'ADN) est l'un des mécanismes d'inactivation programmée des gènes.
La taille et la structure des molécules d'ADN dans les organites
Structure | Poids, millions. daltons | Remarques |
||
|
oh sèche | Animaux | Bague | Dans chaque espèce, toutes les molécules ont la même taille |
|
ra supérieur gémissant | Bague | Varie | Toutes les espèces étudiées possèdent des ADN circulaires de tailles différentes, dans lesquels le contenu total de l'information génétique correspond à une masse de 300 à 1000 millions de daltons, selon les espèces |
|
Champignons: Protozoaires | Bague Bague Bague Linéaire | |||
|
Chlore Paiement poêle | Algue | Bague Bague | ||
plus haut végétaux | Bague | Dans chaque espèce individuelle, des molécules d'une seule ont été trouvées. |
Quantité relative d'ADN organite dans certaines cellules et tissus
organisme | tissu ou type de cellule | Nombre de mol-l ADN/organelle- | Nombre d'organes- nell dans cage | La proportion d'ADN d'organite dans le total ADN cellulaire, % |
|
|
oh sèche | |||||
Cellules de ligne L | |||||
Œuf | |||||
|
Chlore Paiement poêle | Cellules diploïdes végétatives | ||||
Maïs |
Fonctionnement du génome mitochondrial
Quelle est la particularité des mécanismes de réplication et de transcription de l'ADN dans les mitochondries de mammifères ?
Les chaînes complémentaires" href="/text/category/komplementarij/" rel="bookmark">complémentaires de l'ADNmt diffèrent considérablement en termes de densité spécifique, car elles contiennent une quantité inégale de nucléotides puriques "lourds" et de nucléotides pyrimidiques "légers". C'est ainsi que ils sont appelés - chaîne H (lourde - lourde) et L (légère - légère) Au début de la réplication de la molécule d'ADNmt, la boucle dite D (de l'anglais. Boucle de déplacement - boucle de déplacement) est formée Cette structure, visible au microscope électronique, est constituée de sections double brin et simple brin (partie éloignée de la chaîne H). La section double brin est formée par une partie de la chaîne L et une fragment d'ADN nouvellement synthétisé complémentaire avec une longueur de 450-650 (selon le type d'organisme) nucléotides, ayant 5 "- extrémité de l'amorce ribonucléotidique, qui correspond au point de départ de la synthèse de la chaîne H (oriH). La synthèse de la chaîne L ne commence que lorsque la chaîne H fille atteint le point ori L. Cela est dû au fait que la région d'initiation de la réplication de la chaîne L n'est accessible aux enzymes de synthèse de l'ADN que dans un simple brin état, et donc, uniquement dans une double hélice non torsadée dans la synthèse de la chaîne H. Ainsi, les brins filles d'ADNmt sont synthétisés de manière continue et asynchrone (Fig. 3).
Figue 3. Schéma de réplication de l'ADNmt chez les mammifères. Tout d'abord, la boucle D est formée, puis la chaîne H fille est synthétisée, puis la synthèse de la chaîne L fille commence.
La fin du gène de l'ARNr 16S (Fig. 4). Il y a 10 fois plus de transcriptions courtes que de longues. À la suite de la maturation (traitement), de l'ARNr 12S et de l'ARNr 16S sont formés à partir d'eux, qui sont impliqués dans la formation de ribosomes mitochondriaux, ainsi que d'ARNt de phénylalanine et de valine. Les ARNt restants sont excisés des transcrits longs et des ARNm traduits sont formés, aux extrémités 3' desquels sont attachées des séquences polyadényliques.Les extrémités 5' de ces ARNm ne sont pas coiffées, ce qui est inhabituel chez les eucaryotes. L'épissage (fusion) ne se produit pas, car aucun des gènes mitochondriaux des mammifères ne contient d'introns.
Figue 4. Transcription de l'ADNmt humain contenant 37 gènes. Tous les transcrits commencent à être synthétisés dans la région ori H. Les ARN ribosomiques sont excisés des transcrits à chaîne H longue et courte. L'ARNt et l'ARNm sont formés à la suite du traitement à partir des transcrits des deux brins d'ADN. Les gènes d'ARNt sont représentés en vert clair.
Voulez-vous savoir quelles autres surprises le génome mitochondrial peut apporter ? Excellent! Continuer à lire!..
Les régions non codantes leader et 3 ", comme la plupart des ARNm nucléaires. Un certain nombre de gènes contiennent également des introns. Par exemple, le gène box codant pour la cytochrome b oxydase a deux introns. À partir du transcrit d'ARN primaire, de manière autocatalytique (sans la participation d'aucun ou protéines) une copie de la majeure partie du premier intron est découpée. L'ARN restant sert de matrice pour la formation de l'enzyme maturase impliquée dans l'épissage. Une partie de sa séquence d'acides aminés est codée dans les copies restantes des introns. Maturase les coupe, détruisant son propre ARNm, des copies des exons sont fusionnées et l'ARNm de la cytochrome oxydase b est formé (Fig. 5). La découverte de ce phénomène nous a obligés à reconsidérer l'idée des introns comme "rien ne codant des séquences" .
Figue 5. Traitement (maturation) de l'ARNm de la cytochrome oxydase b dans les mitochondries de levure. Au premier stade de l'épissage, l'ARNm est formé, selon lequel la maturase est synthétisée, ce qui est nécessaire pour le deuxième stade de l'épissage.
Lors de l'étude de l'expression des gènes mitochondriaux Trypanosoma brucei ont trouvé un écart surprenant par rapport à l'un des axiomes de base de la biologie moléculaire, qui stipule que la séquence de nucléotides dans l'ARNm correspond exactement à celle des régions codantes de l'ADN. Il s'est avéré que l'ARNm de l'une des sous-unités de la cytochrome c oxydase est modifié, c'est-à-dire qu'après la transcription, sa structure primaire change - quatre uraciles sont insérés. En conséquence, un nouvel ARNm est formé, qui sert de matrice pour la synthèse d'une sous-unité supplémentaire de l'enzyme, dont la séquence d'acides aminés n'a rien à voir avec la séquence des virus, des champignons, Le chercheur anglais Burrell a comparé le structure d'un des gènes mitochondriaux du veau avec la séquence en acides aminés dans la sous-unité de la cytochrome oxydase codée par ce gène, elle est « idéale », c'est-à-dire qu'elle obéit à la règle suivante : « si deux codons ont deux nucléotides identiques, et le les troisièmes nucléotides appartiennent à la même classe (purine - A, G, ou pyrimidine - U, C), alors ils codent pour le même acide aminé. » Dans le code universel, il y a deux exceptions à cette règle : le triplet AUA code pour l'isoleucine, et le codon AUG code pour la méthionine, tandis que dans le code mitochondrial idéal, ces deux triplets sont l'odyute méthionine; le triplet UGG code uniquement le tryptophane, tandis que le triplet UGA code un codon stop. Dans le code universel, les deux déviations concernent les moments fondamentaux de la synthèse protéique : le codon AUG est initiateur, et le codon stop UGA stoppe la synthèse du polypeptide. Le code idéal n'est pas inhérent à toutes les mitochondries décrites, mais aucune d'entre elles n'a de code universel. On peut dire que les mitochondries parlent des langues différentes, mais jamais la langue du noyau.
Différences entre le code génétique "universel" et les deux codes mitochondriaux
codon | Mitochondrial code mammifère | Mitochondrial code levure | “ Universel” |
Comme déjà mentionné, le génome mitochondrial des vertébrés contient 22 gènes d'ARNt. Comment un tel ensemble incomplet sert-il les 60 codons pour les acides aminés (le code idéal de 64 triplets a quatre codons d'arrêt, tandis que le code universel en a trois) ? Le fait est que lors de la synthèse des protéines dans les mitochondries, les interactions codon-anticodon sont simplifiées - deux des trois nucléotides anticodon sont utilisés pour la reconnaissance. Ainsi, un ARNt reconnaît les quatre représentants de la famille des codons, qui ne diffèrent que par le troisième nucléotide. Par exemple, un ARNt de leucine avec l'anticodon GAU se dresse sur le ribosome en face des codons CUU, CUU, CUA et CUG, assurant l'inclusion sans équivoque de la leucine dans la chaîne polypeptidique. Les deux autres codons de leucine UUA et UUG sont reconnus par les ARNt avec l'anticodon AAU. Au total, huit molécules d'ARNt différentes reconnaissent huit familles de quatre codons chacune, et 14 ARNt reconnaissent différentes paires de codons, chacun codant pour un acide aminé.
Il est important que les enzymes aminoacyl-ARNt synthétase responsables de la fixation des acides aminés aux ARNt mitochondriaux correspondants soient codées dans le noyau cellulaire et synthétisées sur les ribosomes du réticulum endoplasmique. Ainsi, chez les vertébrés, tous les composants protéiques de la synthèse mitochondriale des polypeptides sont cryptés dans le noyau. Dans le même temps, la synthèse des protéines dans les mitochondries n'est pas supprimée par le cycloheximide, qui bloque le travail des ribosomes eucaryotes, mais est sensible aux antibiotiques érythromycine et chloramphénicol, qui inhibent la synthèse des protéines chez les bactéries. Ce fait est l'un des arguments en faveur de l'origine des mitochondries à partir de bactéries aérobies lors de la formation symbiotique des cellules eucaryotes.
Importance d'avoir votre propre système génétique pour les mitochondries
Pourquoi les mitochondries ont-elles besoin de leur propre système génétique, alors que d'autres organites, tels que les peroxysomes et les lysosomes, n'en ont pas besoin ? Cette question n'est pas du tout anodine, car le maintien d'un système génétique séparé est coûteux pour une cellule, compte tenu du nombre de gènes supplémentaires requis dans le génome nucléaire. Les protéines ribosomales, les aminoacyl-ARNt synthétases, les ADN et ARN polymérases, les enzymes de traitement et de modification de l'ARN, etc. doivent être codées ici. La plupart des protéines étudiées des mitochondries diffèrent par la séquence d'acides aminés de leurs homologues d'autres parties de la cellule, et là Il y a des raisons de croire qu'il y a très peu de protéines dans ces organites qui pourraient être trouvées ailleurs. Cela signifie que rien que pour maintenir le système génétique des mitochondries, le génome nucléaire doit avoir plusieurs dizaines de gènes supplémentaires. Les raisons de ce "gaspillage" ne sont pas claires, et l'espoir que l'indice sera trouvé dans la séquence nucléotidique de l'ADN mitochondrial ne s'est pas concrétisé. Il est difficile d'imaginer pourquoi les protéines formées dans les mitochondries doivent nécessairement y être synthétisées, et non dans le cytosol.
Habituellement, l'existence d'un système génétique dans les organelles énergétiques s'explique par le fait que certaines des protéines synthétisées à l'intérieur de l'organelle sont trop hydrophobes pour traverser la membrane mitochondriale depuis l'extérieur. Cependant, l'étude du complexe ATP synthétase a montré qu'une telle explication est invraisemblable. Bien que les sous-unités protéiques individuelles de l'ATP synthétase soient hautement conservées au cours de l'évolution, les sites de leur synthèse changent. Dans les chloroplastes, plusieurs protéines plutôt hydrophiles, dont quatre des cinq sous-unités de la partie F1-ATPase du complexe, se forment sur les ribosomes au sein de l'organite. Au contraire, le champignon Neurospora et dans les cellules animales, un composant très hydrophobe (sous-unité 9) de la partie membranaire de l'ATPase est synthétisé sur les ribosomes du cytoplasme et seulement après cela, il passe dans l'organite. La localisation différente des gènes codant pour des sous-unités de protéines fonctionnellement équivalentes dans différents organismes est difficile à expliquer en utilisant une hypothèse postulant certains avantages évolutifs des systèmes génétiques modernes de mitochondries et de chloroplastes.
Compte tenu de tout ce qui précède, il ne reste plus qu'à supposer que le système génétique des mitochondries représente une impasse évolutive. Dans le cadre de l'hypothèse endosymbiotique, cela signifie que le processus de transfert des gènes endosymbiotes dans le génome nucléaire de l'hôte s'est arrêté avant qu'il ne soit complètement achevé.
Héritage cytoplasmique
Les conséquences du transfert de gènes cytoplasmiques pour certains animaux, y compris les humains, sont plus graves que pour la levure. Deux cellules de levure haploïdes fusionnées ont la même taille et contribuent la même quantité d'ADN mitochondrial au zygote résultant. Ainsi, chez la levure, le génome mitochondrial est hérité des deux parents, qui contribuent à parts égales au pool génétique de la progéniture (bien qu'après plusieurs générations séparé les descendants contiendront souvent des mitochondries d'un seul des types parentaux). En revanche, chez les animaux supérieurs, l'ovule apporte plus de cytoplasme au zygote que le sperme, et chez certains animaux, le sperme peut ne pas apporter du tout de cytoplasme. Par conséquent, on peut penser que chez les animaux supérieurs, le génome mitochondrial sera transmis à partir d'un seul parent (à savoir, maternel lignes); en effet, cela a été confirmé expérimentalement. Il s'est avéré, par exemple, que lorsque des rats de deux lignées de laboratoire sont croisés avec de l'ADN mitochondrial légèrement différent dans la séquence nucléotidique (types A et B), on obtient une progéniture contenant
ADN mitochondrial du seul type maternel.
L'hérédité cytoplasmique, contrairement au nucléaire, n'obéit pas aux lois de Mendel. Cela est dû au fait que chez les animaux et les plantes supérieurs, les gamètes de sexes différents contiennent des quantités disparates de mitochondries. Ainsi, dans l'œuf de souris, il y a 90 000 mitochondries et dans le sperme - seulement quatre. De toute évidence, dans un œuf fécondé, les mitochondries proviennent principalement ou uniquement de la femelle, c'est-à-dire que l'héritage de tous les gènes mitochondriaux est maternel. L'analyse génétique de l'hérédité cytoplasmique est difficile en raison des interactions nucléaire-cytoplasmique. Dans le cas de la stérilité mâle cytoplasmique, le génome mitochondrial mutant interagit avec certains gènes nucléaires dont les allèles récessifs sont nécessaires au développement du trait. Les allèles dominants de ces gènes, tant à l'état homozygote qu'hétérozygote, restaurent la fertilité des plantes, quel que soit l'état du génome mitochondrial.
Je voudrais m'attarder sur le mécanisme de transmission maternelle des gènes en rapprochant étude de cas. Afin de comprendre enfin et irrévocablement le mécanisme de l'hérédité non mendélienne (cytoplasmique) des gènes mitochondriaux, considérons ce qui arrive à ces gènes lorsque deux cellules haploïdes fusionnent pour former un zygote diploïde. Dans le cas où une cellule de levure porte une mutation déterminant la résistance de la synthèse des protéines mitochondriales au chloramphénicol et l'autre, une cellule de type sauvage, est sensible à cet antibiotique : les gènes mutants peuvent être facilement identifiés en cultivant la levure sur un milieu contenant le glycérol, qui ne peut être utilisé que par des cellules dont les mitochondries sont intactes ; ainsi, en présence de chloramphénicol, seules les cellules portant le gène mutant peuvent se développer sur un tel milieu. Notre zygote diploïde aura initialement des mitochondries mutantes et de type sauvage. À la suite de la mitose, une cellule fille diploïde bourgeonnera à partir du zygote, qui ne contiendra qu'un petit nombre de mitochondries. Après plusieurs cycles mitotiques, une des nouvelles cellules finira par recevoir toutes les mitochondries, qu'elles soient mutantes ou de type sauvage. Par conséquent, tous les descendants d'une telle cellule auront des mitochondries génétiquement identiques. Un tel processus aléatoire, qui aboutit à la formation d'une progéniture diploïde contenant un seul type de mitochondries, est appelé mitotiquee segrégatione. Lorsqu'une cellule diploïde avec un seul type de mitochondries subit une méiose, les quatre cellules filles haploïdes reçoivent les mêmes gènes mitochondriaux. Ce type d'héritage est appelé nemendeun lion parcourir ou cytoplasmique contrairement à l'héritage mendélien des gènes nucléaires. Le transfert de gènes selon le type cytoplasmique signifie que les gènes étudiés sont localisés dans les mitochondries.
L'étude des génomes mitochondriaux, leur évolution, procédant selon les lois spécifiques de la génétique des populations, la relation entre les systèmes génétiques nucléaire et mitochondrial, est nécessaire pour comprendre l'organisation hiérarchique complexe de la cellule eucaryote et de l'organisme dans son ensemble.
Certaines maladies héréditaires et le vieillissement humain sont associés à certaines mutations de l'ADN mitochondrial ou de gènes nucléaires qui contrôlent le fonctionnement des mitochondries. Les données s'accumulent sur l'implication des défauts de l'ADNmt dans la cancérogenèse. Par conséquent, les mitochondries peuvent être une cible pour la chimiothérapie anticancéreuse. Il existe des faits sur l'interaction étroite des génomes nucléaires et mitochondriaux dans le développement d'un certain nombre de pathologies humaines. De multiples délétions d'ADNmt ont été trouvées chez des patients présentant une faiblesse musculaire sévère, une ataxie, une surdité, un retard mental, hérités de manière autosomique dominante. Le dimorphisme sexuel a été établi dans les manifestations cliniques de la maladie coronarienne, ce qui est très probablement dû à l'effet maternel - l'hérédité cytoplasmique. Le développement de la thérapie génique laisse espérer la correction des défauts des génomes mitochondriaux dans un avenir prévisible.
Comme vous le savez, pour vérifier le fonctionnement de l'un des composants d'un système à plusieurs composants, il devient nécessaire d'éliminer ce composant avec une analyse ultérieure des changements survenus. Puisque le sujet de cet essai est d'indiquer le rôle du génome maternel pour le développement de la progéniture, il serait logique de connaître les conséquences des perturbations dans la composition du génome mitochondrial causées par divers facteurs. Le processus de mutation s'est avéré être un outil pour étudier le rôle ci-dessus, et les conséquences de son action qui nous intéressent sont ce qu'on appelle. maladies mitochondriales.
Les maladies mitochondriales sont un exemple d'hérédité cytoplasmique chez l'homme, ou plutôt « d'hérédité organelle ». Cette précision doit être apportée, car maintenant l'existence, au moins dans certains organismes, de déterminants héréditaires cytoplasmiques non associés aux organites cellulaires - cytogènes (-Vechtomov, 1996) a été prouvée.
Les maladies mitochondriales sont un groupe hétérogène de maladies causées par des défauts génétiques, structurels et biochimiques des mitochondries et une altération de la respiration des tissus. Une analyse généalogique, clinique, biochimique, morphologique et génétique complète est importante pour poser le diagnostic d'une maladie mitochondriale. Le principal signe biochimique pathologie mitochondriale est le développement de l'acidose lactique, l'acidémie hyperlactate est généralement détectée en association avec l'acidémie hyperpyruvate. Le nombre de variantes différentes a atteint 120 formes. Il y a une augmentation stable de la concentration des acides lactique et pyruvique dans le liquide céphalo-rachidien.
Les maladies mitochondriales (MD) sont un problème important pour la médecine moderne. Selon les modes de transmission héréditaire, chez les MB, il existe des maladies héréditaires monogéniques selon le type mendélien, dans lesquelles, du fait de la mutation de gènes nucléaires, soit la structure et le fonctionnement des protéines mitochondriales sont perturbés, soit l'expression de modifications de l'ADN mitochondrial, ainsi que des maladies causées par des mutations des gènes mitochondriaux, qui sont principalement transmises à la progéniture par la lignée maternelle.
Données études morphologiques indiquant une pathologie grossière des mitochondries: prolifération anormale des mitochondries, polymorphisme des mitochondries avec une violation de la forme et de la taille, désorganisation des crêtes, accumulations de mitochondries anormales sous le sarcolemme, inclusions paracristallines dans les mitochondries, présence de vacuoles interfibrillaires
Formes de maladies mitochondriales
1 . Maladies mitochondriales causées par des mutations de l'ADN mitochondrial
1.1 Maladies causées par des délétions de l'ADN mitochondrial
1.1.1.Syndrome de Kearns-Sayre
La maladie se manifeste à l'âge de 4 à 18 ans, ophtalmoplégie externe progressive, rétinite pigmentaire, ataxie, tremblement intentionnel, bloc cardiaque auriculo-ventriculaire, augmentation des taux de protéines dans le liquide céphalo-rachidien supérieur à 1 g/l, fibres rouges "déchiquetées" dans le squelette biopsies musculaires
1.1.2 Syndrome de Pearson
Au début de la maladie dès la naissance ou dans les premiers mois de la vie, il est parfois possible de développer des encéphalomyopathies, ataxie, démence, ophtalmoplégie externe progressive, anémie hypoplasique, altération de la fonction pancréatique exocrine, évolution progressive
2 .Maladies causées par des mutations ponctuelles dans l'ADN mitochondrial
Hérédité de type maternel, diminution aiguë ou subaiguë de l'acuité visuelle d'un ou des deux yeux, association avec des troubles neurologiques et ostéoarticulaires, microangiopathie rétinienne, évolution progressive avec possibilité de rémission ou de restauration de l'acuité visuelle, début de la maladie à l'âge de 20 ans -30 ans
2.2. Syndrome NAPR (neuropathie, ataxie, rétinite pigmentaire)
Type d'hérédité maternelle, une combinaison de neuropathie, ataxie et rétinite pigmentaire, retard psychomoteur, démence, présence de fibres rouges "déchirées" dans les biopsies de tissus musculaires
2.3 Syndrome MERRF (myoclonie-épilepsie, fibres rouges « déchirées »)
Type d'hérédité maternelle, apparition de la maladie à l'âge de 3 à 65 ans, épilepsie myoclonique, ataxie, démence associée à une surdité neurosensorielle, atrophie des nerfs optiques et troubles de la sensibilité profonde, acidose lactique, l'examen EEG révèle une épilepsie généralisée du bain complexes, fibres rouges "en lambeaux" dans les échantillons de biopsie musculaire squelettique, évolution progressive
2.4 Syndrome MELAS (encéphalomyopathie mitochondriale, acidose lactique, épisodes de type AVC)
Type d'hérédité maternelle, apparition de la maladie avant l'âge de 40 ans, intolérance à l'exercice, céphalées de type migraine avec nausées et vomissements, épisodes ressemblant à des accidents vasculaires cérébraux, convulsions, acidose lactique, fibres rouges "déchiquetées" dans les biopsies musculaires, évolution progressive.
3 .Pathologie associée à des défauts de communication intergénomique
3.1 Syndromes de délétions multiples de l'ADN mitochondrial
Blépharoptose, ophtalmoplégie externe, faiblesse musculaire, surdité neurosensorielle, atrophie du nerf optique, évolution progressive, fibres rouges "déchirées" dans les biopsies des muscles squelettiques, diminution de l'activité des enzymes de la chaîne respiratoire.
3.2 Syndrome de délétion de l'ADN mitochondrial
Mode de transmission autosomique récessif
Formes cliniques :
3.2.1.mortel infantile
a) insuffisance hépatique sévère b) hépatopathie c) hypotension musculaire
Débuts en période néonatale
3.2.2.myopathie congénitale
Faiblesse musculaire sévère, hypotension généralisée, cardiomyopathie et convulsions, lésions rénales, glycosurie, aminoacidopathie, phosphaturie
3.2.3.Myopathie infantile
survient au cours des 2 premières années de vie, faiblesse musculaire progressive, atrophie des groupes musculaires proximaux et perte des réflexes tendineux, évolution rapidement progressive, décès au cours des 3 premières années de vie.
4 .Maladies mitochondriales causées par des mutations de l'ADN nucléaire
4.1 Maladies associées à des défauts de la chaîne respiratoire
4.1.1 Déficit en complexe 1 (NADH:CoQ réductase)
Apparition avant l'âge de 15 ans, syndrome de myopathie, retard psychomoteur, troubles cardiovasculaires, crises épileptiques résistantes au traitement, troubles neurologiques multiples, évolution progressive
4.1.2 Déficit en complexe 2 (succinate-CoQ-réductase)
Caractérisé par le syndrome d'encéphalomyopathie, évolution progressive, convulsions, ptosis peut se développer
4.1.3 Déficit en complexe 3 (CoQ-cytochrome C-oxydoréductase)
Troubles multisystémiques, lésions de divers organes et systèmes, impliquant le système nerveux central et périphérique, le système endocrinien, les reins, évolution progressive
4.1.4 Déficit du complexe (cytochrome C-oxydase)
4.1.4.1 Acidose lactique congénitale infantile mortelle
Myopathie mitochondriale avec insuffisance rénale ou cardiomyopathie, début à l'âge néonatal, troubles respiratoires sévères, hypotension musculaire diffuse, évolution progressive, décès dans la première année de vie.
4.1.4.2.Faiblesse musculaire infantile bénigne
Atrophie, avec un traitement adéquat et opportun, il est possible de stabiliser rapidement le processus et de récupérer en 1 à 3 ans de vie
5 .Syndrome de Menkes (trichopolyodystrophie)
Un fort retard du développement psychomoteur, un retard de croissance, une croissance dystrophique et des modifications capillaires,
6 . Encéphalomyopathies mitochondriales
6.1.Syndrome de Leigh(encéphalomyélopathie névrotique subaiguë)
Se manifestant après 6 mois de vie, on note souvent une hypotension musculaire, une ataxie, un nystagmus, des symptômes pyramidaux, une ophtalmoplégie, une atrophie des nerfs optiques, une cardiomyopathie et une acidose métabolique légère.
6.2.Syndrome d'Alpers(polydystrophie sclérosante progressive)
Dégénérescence de la matière grise du cerveau associée à une cirrhose du foie, déficit en complexe 5 (ATP synthétase), retard psychomoteur, ataxie, démence, faiblesse musculaire, évolution progressive de la maladie, pronostic défavorable
6.3 Déficit en coenzyme-Q
Crises métaboliques, faiblesse et fatigue musculaires, ophtalmoplégie, surdité, diminution de la vision, épisodes de type accident vasculaire cérébral, ataxie, épilepsie myoclonique, lésions rénales : glycosurie, aminoacidopathie, phosphaturie, troubles endocriniens, évolution progressive, diminution de l'activité des enzymes de la chaîne respiratoire
7 .Maladies associées à une altération du métabolisme des acides lactique et pyruvique
7.1. Déficit en pyruvate carboxylase Type de transmission autosomique récessive, apparition de la maladie dans la période néonatale, complexe de symptômes "enfant paresseux", convulsions résistantes au traitement, concentrations élevées de corps cétoniques dans le sang, hyperammoniémie, hyperlysinémie, diminution de l'activité de la pyruvate carboxylase dans les muscles squelettiques
7.2 Déficit en pyruvate déshydrogénase
Manifestation dans la période néonatale, dysmorphie craniofaciale, convulsions résistantes au traitement, troubles respiratoires et de succion, complexe de symptômes "enfant paresseux", dysgénésie cérébrale, acidose prononcée avec une teneur élevée en lactate et en pyruvate
7.3 Diminution de l'activité de la pyruvate déshydrogénase
Présentation dans la première année de vie, microcéphalie, retard psychomoteur, ataxie, dystonie musculaire, choréoathétose, acidose lactique à pyruvate élevé
7.4 Déficit en dihydrolipoyl transacétylase
Transmission autosomique récessive, apparition de la maladie au cours de la période néonatale, microcéphalie, retard psychomoteur, hypotension musculaire avec augmentation ultérieure du tonus musculaire, atrophie des disques optiques, acidose lactique, diminution de l'activité de la dihydrolipoyl transacétylase
7.5 Déficit en dihydrolipoyl déshydrogénase
Hérédité de type autosomique récessif, apparition de la maladie au cours de la première année de vie, complexe symptomatique "enfant paresseux", crises dysmétaboliques avec vomissements et diarrhée, retard psychomoteur, atrophie des disques optiques, acidose lactique, augmentation du taux d'alanine dans le sang sérum, α-cétoglutarate, acides α-céto à chaîne ramifiée, diminution de l'activité di-hydrolipoyl déshydrogénase
8 .Maladies dues à des défauts de bêta-oxydation des acides gras
8.1 Déficit en acétyl-CoA déshydrogénase à longue chaîne
Héritage de type autosomique récessif, apparition de la maladie dans les premiers mois de la vie, crises métaboliques avec vomissements et diarrhée, complexe symptomatique "enfant paresseux", hypoglycémie, acidurie dicarboxylique, diminution de l'activité de l'acétyl-CoA déshydrogénase des acides gras à longue chaîne
8.2 Déficit en acétyl-CoA déshydrogénase à chaîne moyenne
Transmission de type autosomique récessif, apparition de la maladie dans la période néonatale ou les premiers mois de la vie, crises métaboliques avec vomissements et diarrhée,
faiblesse musculaire et hypotension, le syndrome se développe souvent mort subite, hypoglycémie, acidurie dicarboxylique, diminution de l'activité de l'acétyl-CoA déshydrogénase des acides gras à chaîne moyenne
8.3. Déficit en acide gras à chaîne courte en acétyl-CoA déshydrogénase
Transmission de type autosomique récessif, âge d'apparition de la maladie différent, diminution de la tolérance à activité physique, crises métaboliques avec vomissements et diarrhée, faiblesse musculaire et hypotonie, augmentation de l'excrétion urinaire d'acide méthylsuccinique, acétyl-CoA-déshydrogénase des acides gras à chaîne carbonée courte
8.4 Déficit multiple en acétyl-CoA déshydrogénases des acides gras
forme néonatale: dysmorphie craniofaciale, dysgénésie cérébrale, hypoglycémie et acidose sévères, évolution maligne, diminution de l'activité de toutes les déshydrogénases des acides gras acétyl-CoA,
Forme infantile : complexe de symptômes « enfant paresseux », cardiomyopathie, crises métaboliques, hypoglycémie et acidose
8.5 Diminution de l'activité de toutes les acétyl-CoA déshydrogénases d'acides gras
Forme des débuts tardifs : les épisodes périodiques de faiblesse musculaire, les crises métaboliques, l'hypoglycémie et l'acidose sont moins prononcés, l'intelligence est préservée,
9 .Fermentopathies du cycle de Krebs
9.1 Déficit en fumarase
Mode de transmission autosomique récessif, apparition de la maladie au cours de la période néonatale ou néonatale, microcéphalie, faiblesse musculaire généralisée et hypotension, épisodes de léthargie, encéphalopathie à évolution rapide, mauvais pronostic
9.2 Déficit en succinate déshydrogénase
Une maladie rare caractérisée par une encéphalomyopathie progressive
9.3 Déficit en alpha-cétoglutarate déshydrogénase
Type de transmission autosomique récessive, apparition néonatale de la maladie, microcéphalie, complexe de symptômes « bébé paresseux », épisodes de léthargie, acidose lactique, évolution rapidement progressive, diminution de la teneur en enzymes du cycle de Krebs dans les tissus
9.4. Syndromes de carence en carnitine et en enzymes de son métabolisme
Déficit en carnitine palmitoyltransférase-1, transmission autosomique récessive, apparition précoce de la maladie, épisodes de coma hypoglycémique non cétonémique, hépatomégalie, hypertriglycéridémie et hyperammoniémie modérée, diminution de l'activité de la carnitine palmitoyltransférase-1 dans les fibroblastes et les cellules hépatiques
9.5 Déficit en carnitine acylcarnitine translocase
Apparition précoce de la maladie, troubles cardiovasculaires et respiratoires, complexe de symptômes "enfant paresseux", épisodes de léthargie et de coma, augmentation des concentrations d'esters de carnitine et d'une longue chaîne carbonée dans le contexte d'une diminution de la carnitine libre dans le sérum sanguin, diminution de l'activité de carnitine-acylcarnitine translocase
9.6. Déficit en carnitine palmitoyltransférase-2
Transmission autosomique récessive, faiblesse musculaire, myalgie, myoglobinurie, diminution de l'activité de la carnitine palmitoyltransférase-2 dans les muscles squelettiques
Type de transmission autosomique récessive, complexe de symptômes myopathiques, épisodes de léthargie et de léthargie, cardiomyopathie, épisodes d'hypoglycémie, diminution du taux de carnitine dans le sérum sanguin et augmentation de son excrétion dans l'urine.
Après analyse d'une liste aussi « terrible » de pathologies associées à certaines modifications du fonctionnement du génome mitochondrial (et pas seulement), certaines questions se posent. Quels sont les produits des gènes mitochondriaux et à quels processus cellulaires super-méga-vitaux participent-ils ?
Il s'est avéré que certaines des pathologies ci-dessus peuvent survenir avec une synthèse altérée de 7 sous-unités du complexe NADH déshydrogénase, 2 sous-unités d'ATP synthétase, 3 sous-unités de cytochrome c oxydase et 1 sous-unité d'ubiquinol-cytochrome-c-réductase (cytochrome b) , qui sont les produits géniques des mitochondries. Sur cette base, nous pouvons conclure que ces protéines jouent un rôle clé dans les processus de respiration cellulaire, d'oxydation des acides gras et de synthèse d'ATP, de transfert d'électrons dans le système de transport d'électrons de la membrane mt interne, du fonctionnement du système antioxydant, etc.
A en juger par les dernières données sur les mécanismes de l'apoptose, de nombreux scientifiques sont arrivés à la conclusion qu'il existe un centre de contrôle de l'apoptose précisément ...
Le rôle des protéines mitochondriales a également été démontré dans l'utilisation d'antibiotiques qui bloquent la synthèse de la MT. Si des cellules humaines en culture tissulaire sont traitées avec un antibiotique, tel que la tétracycline ou le chloramphénicol, après une ou deux divisions, leur croissance s'arrêtera. Cela est dû à l'inhibition de la synthèse des protéines mitochondriales, conduisant à l'apparition de mitochondries défectueuses et, par conséquent, à une formation insuffisante d'ATP. Pourquoi, alors, les antibiotiques peuvent-ils être utilisés dans le traitement des infections bactériennes ? Il y a plusieurs réponses à cette question:
1. Certains antibiotiques (comme l'érythromycine) ne traversent pas la membrane interne des mitochondries des mammifères.
2. La plupart des cellules de notre corps ne se divisent pas ou se divisent très lentement, de sorte que le remplacement des mitochondries existantes par de nouvelles est tout aussi lent (dans de nombreux tissus, la moitié des mitochondries sont remplacées en environ cinq jours ou plus). Ainsi, le nombre de mitochondries normales ne diminuera à un niveau critique que si le blocage de la synthèse des protéines mitochondriales est maintenu pendant plusieurs jours.
3. Certaines conditions à l'intérieur des tissus empêchent la pénétration de certains médicaments dans les mitochondries des cellules les plus sensibles. Par exemple, une concentration élevée de Ca2+ dans la moelle osseuse conduit à la formation d'un complexe Ca2+-tétracycline, qui ne peut pas pénétrer les précurseurs des cellules sanguines qui se divisent rapidement (et donc les plus vulnérables).
Ces facteurs permettent d'utiliser certains médicaments qui inhibent la synthèse des protéines mitochondriales comme antibiotiques dans le traitement des animaux supérieurs. Seuls deux de ces médicaments ont effet secondaire: un traitement à long terme avec de fortes doses de chloramphénicol peut entraîner une altération de la fonction hématopoïétique de la moelle osseuse (suppression de la formation de globules rouges et de globules blancs), et l'utilisation à long terme de tétracycline peut endommager l'épithélium intestinal. Mais dans les deux cas, il n'est pas encore tout à fait clair si ces Effets secondaires blocage de la biogenèse mitochondriale ou d'autres raisons.
Conclusion
Les caractéristiques structurelles et fonctionnelles du génome mt sont les suivantes. Tout d'abord, il a été établi que l'ADNmt est transmis de la mère à tous ses
descendants et de ses filles à toutes les générations suivantes, mais les fils ne transmettent pas leur ADN (héritage maternel). caractère maternel
La transmission de l'ADNmt est probablement due à deux circonstances : soit la proportion d'ADNmt paternel est si faible (pas
plus d'une molécule d'ADN pour 25 000 ADNmt maternel) qu'ils ne peuvent pas être détectés par les méthodes existantes, ou après la fécondation, la réplication des mitochondries paternelles est bloquée. Deuxièmement, l'absence de variabilité combinatoire - l'ADNmt n'appartient qu'à l'un des parents, par conséquent, il n'y a pas d'événements de recombinaison caractéristiques de l'ADN nucléaire dans la méiose, et la séquence de nucléotides change de génération en génération uniquement en raison de mutations. Troisièmement, l'ADNmt n'a pas d'introns.
(forte probabilité qu'une mutation aléatoire touche la région codante de l'ADN), des histones protectrices et un système efficace de réparation de l'ADN - tout cela détermine un taux de mutation 10 fois plus élevé que dans l'ADN nucléaire. Quatrièmement, l'ADNmt normal et mutant peut coexister simultanément à l'intérieur d'une cellule - le phénomène d'hétéroplasmie (la présence uniquement d'ADNmt normal ou mutant est appelée homoplasmie). Enfin, les deux brins sont transcrits et traduits en ADNmt, et le code génétique de l'ADNmt diffère du code universel par un certain nombre de caractéristiques (UGA code pour le tryptophane, AUA code pour la méthionine, AGA et AGG sont stop-
codons).
Ces propriétés et les fonctions susmentionnées du génome mt ont fait de l'étude de la variabilité de la séquence nucléotidique de l'ADNmt un outil inestimable pour les médecins, les médecins légistes, les biologistes de l'évolution,
représentants de la science historique dans la résolution de leurs tâches spécifiques.
Depuis 1988, date à laquelle on a découvert que les mutations du gène de l'ADNmt sous-tendent les myopathies mitochondriales (J. Y. Holt et al., 1988) et la neuropathie optique héréditaire de Leber (D. C. Wallace, 1988), une identification plus systématique des mutations dans le génome mt humain a conduit à la formation de le concept de maladies mitochondriales (MD). À l'heure actuelle, des mutations pathologiques de l'ADNmt ont été découvertes dans tous les types de gènes mitochondriaux.
Bibliographie
1. Skulachev, mitochondries et oxygène, Soros. éduquer. magazine
2. Fondamentaux de la biochimie : En trois tomes, M. : Mir, .
3. Nicholes D. G. Bioenergetics, An Introd. au Chimiosme. Th., Acad. Presse, 1982.
4. Stryer L. Biochimie, 2e éd. San Francisco, Freeman, 1981.
5. Skoulatchev membranes biologiques. M., 1989.
6., réticulum de Chentsov : Structure et certaines fonctions // Résultats de la science. Problèmes généraux de biologie. 1989
7. Cytologie de Chentsov. M.: Maison d'édition de l'Université d'État de Moscou, 1995
8. , La sphère de compétence du génome mitochondrial // Vestn. RAMN, 2001. ‹ 10. S. 31-43.
9. Holt I. J, Harding A. E., Morgan-Hughes I. A. Suppression de l'ADN mitochondrial musculaire chez les patients atteints de myopathies mitochondriales. Nature, 1988, 331:717-719.
10. et etc. Le génome humain et les gènes de prédisposition. SPb., 2000
11. , Génome mitochondrial. Novossibirsk, 1990.
12. // Soro. éduquer. magazine 1999. N° 10. pp.11-17.
13. Le rôle de la symbiose dans l'évolution cellulaire. M., 1983.
14. // Soro. éduquer. magazine 1998. N° 8. S.2-7.
15. // Soro. éduquer. magazine 2000. N° 1. S.32-36.
Université nationale de Kyiv. Taras Chevtchenko
Département de biologie
abstrait
sur le sujet:
« Le rôle du génome maternel dans le développement de la progéniture »
Aveclàenta IVcours
Département de biochimie
Frolova Artem
Kyiv 2004
Planifier:
Introduction................................................. ..............................une
Théorie symbiotique de l'origine des mitochondries ...... 2
Le rôle du noyau cellulaire dans la biogenèse mitochondriale .................................................. .....................5
Systèmes de transport des mitochondries .................................................. .................. ......7
La taille et la forme des génomes mitochondriaux .......................10
Fonctionnement du génome mitochondrial..............14
Signification de la présence de son propre système génétique pour les mitochondries ......................................... ...................... ................................ .......19
Hérédité cytoplasmique................................................20
Pourquoi les mitochondries ont-elles leur propre ADN ? Mais pourquoi les symbiotes n'auraient-ils pas leur ADN en eux, produisant tout ce dont ils ont besoin localement ? Pourquoi alors transférer une partie de l'ADN mitochondrial vers le noyau cellulaire, créant ainsi le besoin de transporter les produits géniques vers les mitochondries ? Pourquoi les mitochondries ne sont-elles transmises que d'un seul parent ? Comment les mitochondries obtenues de la mère s'articulent-elles avec le génome cellulaire, constitué de l'ADN de la mère et du père ? Plus les gens en apprennent sur les mitochondries, plus les questions se posent.

Cependant, cela ne s'applique pas seulement aux mitochondries : dans n'importe quel domaine de n'importe quelle science, l'expansion de la sphère de la connaissance ne conduit qu'à une augmentation de sa surface, au contact de l'inconnu, provoquant de plus en plus de nouvelles questions, dont les réponses seront étendre cette même sphère avec le même résultat prévisible.
Ainsi, l'ADN des mitochondries modernes est distribué de manière très étrange : une petite partie des gènes est contenue directement dans les mitochondries du chromosome en anneau (plus précisément, dans plusieurs copies du même chromosome dans chaque mitochondrie), et la plupart des schémas de production parties constitutives Les mitochondries sont stockées dans le noyau de la cellule. Par conséquent, la copie de ces gènes se produit simultanément avec la copie du génome de l'organisme entier, et les produits qu'ils produisent vont très loin du cytoplasme de la cellule aux mitochondries. Cependant, cela est pratique à bien des égards : les mitochondries n'ont pas besoin de copier tous ces gènes lorsqu'elles se reproduisent, les lisent et construisent des protéines et d'autres composants, en se concentrant sur leur fonction principale de production d'énergie. Pourquoi, alors, y a-t-il encore un petit ADN dans les mitochondries, pour le maintien duquel tous ces mécanismes sont nécessaires, s'être débarrassé de quelles mitochondries pourrait jeter encore plus de ressources sur le but principal de leur existence?

Dans un premier temps, on supposait que l'ADN restant dans les mitochondries était un atavisme, héritage de la pro-mitochondrie absorbée par le méthanogène, qui possède un génome bactérien complet. Au début de leur symbiose, malgré l'existence dans le noyau de ces gènes mitochondriaux ( gènes m), qui étaient nécessaires pour maintenir un environnement confortable pour les pro-mitochondries à l'intérieur du méthanogène (ceci est décrit en détail dans la section sur les mitochondries), les mêmes gènes étaient stockés dans chacune des mitochondries. La pro-mitochondrie au début de sa vie en tant que symbiote ressemblait beaucoup à la bactérie moderne dans le diagramme à gauche de ce paragraphe.
Et très lentement, faute de demande, ces gènes ont disparu du chromosome mitochondrial à la suite de diverses mutations. Mais le noyau cellulaire a accumulé de plus en plus de gènes m qui sont entrés dans le cytoplasme à partir des symbiotes mitochondriaux détruits et intégrés dans le génome de la chimère eucaryote. Dès que le gène m nouvellement intégré a commencé à être lu, les mécanismes cellulaires ont produit les produits nécessaires aux mitochondries, libérant les symbiotes de leur création indépendante. Cela signifie que l'analogue mitochondrial du gène qui est passé dans le noyau n'a plus été maintenu en état de marche par la sélection naturelle et a été effacé par des mutations au même titre que tous les précédents. Par conséquent, il serait logique de supposer que bientôt les gènes qui restent encore dans les mitochondries passeront dans le noyau, ce qui conduira à de grands avantages énergétiques pour les eucaryotes : après tout, les mécanismes encombrants de copie, de lecture et de réparation de l'ADN peuvent être supprimés de chaque mitochondrie, et donc tout ce dont vous avez besoin pour fabriquer des protéines.

Arrivés à cette conclusion, les scientifiques ont calculé le temps qu'il faudrait à tous les gènes pour migrer des mitochondries vers le noyau par dérive naturelle. Et il s'est avéré que cette période est révolue depuis longtemps. Au moment de l'apparition de la cellule eucaryote, les mitochondries avaient un génome bactérien commun de plusieurs milliers de gènes (les scientifiques déterminent ce qu'était ce génome en étudiant les gènes m transférés au noyau dans différents organismes), et maintenant les mitochondries de toutes les espèces eucaryotes ont perdu de 95 à 99,9 % de leurs gènes. Personne n'a plus d'une centaine de gènes dans les mitochondries, mais personne non plus n'a de mitochondries sans gènes. Si le hasard jouait un rôle clé dans ce processus, alors au moins plusieurs espèces auraient déjà franchi le chemin du transfert de gènes vers le noyau jusqu'au bout. Mais cela ne s'est pas produit, et les mitochondries de différentes espèces étudiées à l'heure actuelle, perdant leurs gènes indépendamment les unes des autres, en ont conservé le même ensemble, ce qui indique directement la nécessité de la présence de ces gènes particuliers dans les mitochondries.

De plus, d'autres organites cellulaires producteurs d'énergie, les chloroplastes, ont également leur propre ADN, et de la même manière, des chloroplastes d'espèces différentes ont évolué en parallèle et indépendamment, chacun restant avec le même ensemble de gènes.
Cela signifie que tous ces inconvénients importants liés au maintien de son propre génome dans chaque mitochondrie cellulaire (et en moyenne, il y en a plusieurs centaines dans une cellule!) Et un appareil encombrant pour le copier-corriger-traduire (le principal, mais pas tous! Vous pouvez voir ses parties sur la photo de gauche) sont quelque peu dépassées.
Et pour le moment, il existe une théorie cohérente de ce «quelque chose»: la capacité de produire certains détails des mitochondries directement à l'intérieur est nécessaire pour réguler le rythme de la respiration et ajuster les processus se produisant dans les mitochondries aux besoins en constante évolution de la organisme entier.

Imaginez que l'une des centaines de mitochondries d'une cellule manque soudainement d'éléments de la chaîne respiratoire (voir les détails à ce sujet dans), ou qu'elle ne contient pas assez d'ATP synthase. Il s'avère être soit surchargé de nourriture et d'oxygène et ne peut pas les traiter assez rapidement, soit son espace intermembranaire regorge de protons qui n'ont nulle part où aller - un désastre complet en général. Bien sûr, tous ces écarts par rapport à la situation de vie idéale déclenchent de multiples signaux visant à équilibrer le roulis d'un navire qui coule.
Ces signaux déclenchent la production des parties exactes qui manquent actuellement aux mitochondries, activant la lecture des gènes qui fabriquent les protéines. Dès que la mitochondrie aura suffisamment de composants de la chaîne respiratoire ou d'ATPases, le «rouleau s'égalisera», les signaux sur la nécessité de construire de nouvelles pièces cesseront d'arriver et les gènes seront à nouveau désactivés. C'est l'un des mécanismes nécessaires de l'autorégulation cellulaire, étonnamment élégant dans sa simplicité, la moindre violation de celui-ci conduit à une maladie grave voire à la non-viabilité de l'organisme.

Essayons de déterminer logiquement où doivent se situer les gènes nécessaires pour répondre à ce signal de détresse. Imaginez la situation où ces gènes se trouvent dans le noyau d'une cellule contenant quelques centaines de mitochondries. Dans l'une des mitochondries, par exemple, il y avait une carence NADH déshydrogénases: la première enzyme de la chaîne respiratoire, dont le rôle est de dépouiller deux électrons de la molécule de NADH, de les transmettre à l'enzyme suivante et de pomper 2 à 4 protons à travers la membrane.
En fait, de telles carences en enzyme se produisent assez souvent, car elles échouent périodiquement, la quantité de nourriture consommée change constamment, les besoins de la cellule en ATP sautent également après les sauts ou les vautres de l'organisme qui contient cette cellule. Par conséquent, la situation est très typique. Et maintenant, la mitochondrie émet un signal: "Nous devons construire plus de NADH-déshydrogénase!", Qui dépasse ses limites, traverse le cytoplasme jusqu'au noyau, pénètre dans le noyau et commence à lire les gènes nécessaires. Selon les normes cellulaires, le temps de transit de ce signal est très important, et vous devez également tirer l'ARN messager construit du noyau dans le cytoplasme, créer des protéines en l'utilisant et les envoyer aux mitochondries ...
Et ici se pose un problème bien plus important que la perte de temps supplémentaire : lors de la création de protéines mitochondriales spécialisées, elles sont marquées du signal « délivrer aux mitochondries », mais laquelle ? Inconnue. Par conséquent, les protéines dont ils n'ont pas besoin commencent à pénétrer dans chacune des quelques centaines de mitochondries. La cellule dépense des ressources pour leur production et leur délivrance, les mitochondries sont remplies de chaînes respiratoires supplémentaires (ce qui entraîne une inefficacité des processus respiratoires), et la seule mitochondrie qui a besoin de ces protéines ne les reçoit pas en quantité suffisante, car elle obtient, au mieux, un centième de ce qui est produit. Alors elle continue d'envoyer des appels de détresse et le chaos continue. Même à partir de cette description lyrique et superficielle de ce qui se passe, il est clair qu'une telle cellule n'est pas viable. Et qu'il existe des gènes qui doivent être lus et traduits directement dans les mitochondries afin de réguler les processus qui s'y déroulent, et non pas sur le plan de noyau lancé par lots pour la production d'ongles .. c'est-à-dire des protéines de la chaîne respiratoire pour tous mitochondries à la fois.

Après avoir vérifié ce qui est exactement produit par les organismes restant dans les mitochondries (et donc déplaçant les gènes m dans le noyau indépendamment les uns des autres), ils ont découvert que ce sont précisément les éléments nécessaires à la construction des chaînes respiratoires et de l'ATPase, ainsi que des ribosomes (qui c'est-à-dire le dispositif de transmission de la partie principale).
Vous pouvez en savoir plus à ce sujet (et plus) de Lane dans « Énergie, sexe, suicide : les mitochondries et le sens de la vie ». Eh bien, vous pouvez simplement comparer le schéma de l'ADN mitochondrial, où les produits codés sont décodés (à droite de ce paragraphe), avec le schéma de la chaîne respiratoire (ci-dessus), de sorte qu'il devienne clair ce qui est exactement produit dans les mitochondries. Bien sûr, toutes les protéines intégrées à cette chaîne ne sont pas produites localement, certaines d'entre elles sont construites dans le cytoplasme de la cellule. Mais les principales "ancres" sur lesquelles s'accrochent le reste des détails sont créées à l'intérieur des mitochondries. Cela vous permet de produire exactement autant d'enzymes que vous en avez besoin, et exactement là où elles sont nécessaires.
Comment les mitochondries sont liées au sexe et comment différents génomes s'entendent dans une cellule, j'écrirai dans l'un des prochains chapitres de cette ligne.
Article principal : ADN mitochondrial
L'ADN mitochondrial situé dans la matrice est une molécule circulaire fermée à double brin, dans les cellules humaines ayant une taille de 16569 paires de nucléotides, soit environ 10 5 fois plus petit que l'ADN localisé dans le noyau. En général, l'ADN mitochondrial code pour 2 ARNr, 22 ARNt et 13 sous-unités d'enzymes de la chaîne respiratoire, ce qui ne représente pas plus de la moitié des protéines qui s'y trouvent. En particulier, sous le contrôle du génome mitochondrial, sept sous-unités d'ATP synthétase, trois sous-unités de cytochrome oxydase et une sous-unité de cytochrome d'ubiquinol sont codées. Avec-réductases. Dans ce cas, toutes les protéines, à l'exception d'un, deux ARN ribosomiques et six ARN de transport sont transcrites à partir de la chaîne d'ADN plus lourde (externe), et 14 autres ARNt et une protéine sont transcrites à partir de la chaîne plus légère (interne).
Dans ce contexte, le génome mitochondrial végétal est beaucoup plus grand et peut atteindre 370 000 paires de nucléotides, soit environ 20 fois plus grand que le génome mitochondrial humain décrit ci-dessus. Le nombre de gènes y est également environ 7 fois supérieur, ce qui s'accompagne de l'apparition dans les mitochondries végétales de voies de transport d'électrons supplémentaires non associées à la synthèse d'ATP.
L'ADN mitochondrial se réplique en interphase, ce qui est en partie synchronisé avec la réplication de l'ADN dans le noyau. Au cours du cycle cellulaire, les mitochondries se divisent en deux par constriction, dont la formation commence par un sillon annulaire sur la membrane mitochondriale interne. Une étude détaillée de la séquence nucléotidique du génome mitochondrial a permis d'établir que les déviations du code génétique universel ne sont pas rares dans les mitochondries des animaux et des champignons. Ainsi, dans les mitochondries humaines, le codon TAT au lieu d'isoleucine dans le code standard code pour l'acide aminé méthionine, les codons TCT et TCC, codant généralement pour l'arginine, sont des codons stop, et le codon ACT, qui est le codon stop dans le code standard. , code pour l'acide aminé méthionine. Quant aux mitochondries végétales, elles semblent utiliser un code génétique universel. Une autre caractéristique des mitochondries est la caractéristique de la reconnaissance des codons d'ARNt, qui consiste dans le fait qu'une telle molécule est capable de reconnaître non pas un, mais trois ou quatre codons à la fois. Caractéristique spécifiée réduit l'importance du troisième nucléotide dans le codon et conduit au fait que les mitochondries nécessitent une plus petite variété de types d'ARNt. Dans ce cas, seuls 22 ARNt différents suffisent.
Possédant son propre appareil génétique, la mitochondrie possède également son propre système de synthèse des protéines, dont une caractéristique dans les cellules des animaux et des champignons sont de très petits ribosomes caractérisés par un coefficient de sédimentation de 55S, qui est encore inférieur à celui des 70S-ribosomes de type procaryote. Dans le même temps, deux gros ARN ribosomiques sont également plus petits que chez les procaryotes, et le petit ARNr est totalement absent. Dans les mitochondries végétales, au contraire, les ribosomes ressemblent davantage aux procaryotes en taille et en structure.
Protéines mitochondriales[modifier | modifier la source]
Le nombre de protéines traduites à partir d'ARNm mitochondrial, qui forment des sous-unités de grands complexes enzymatiques, est limité. Une partie importante des protéines est codée dans le noyau et synthétisée sur les ribosomes 80S cytoplasmiques. En particulier, certaines protéines se forment de cette manière - transporteurs d'électrons, translocases mitochondriales, composants du transport des protéines dans les mitochondries, ainsi que des facteurs nécessaires à la transcription, à la traduction et à la réplication de l'ADN mitochondrial. En même temps, ces protéines ont des peptides signal spéciaux à leur extrémité N-terminale, dont la taille varie de 12 à 80 résidus d'acides aminés. Ces sites forment des boucles amphiphiles, assurent un contact spécifique des protéines avec les domaines de liaison des récepteurs de reconnaissance mitochondriale localisés sur la membrane externe. Vers la membrane mitochondriale externe, ces protéines sont transportées à l'état partiellement déplié en association avec des protéines chaperonnes (en particulier, avec hsp70). Après transfert à travers les membranes externe et interne, aux points de leurs contacts, les protéines entrant dans les mitochondries se lient à nouveau à des chaperons, mais d'origine mitochondriale propre, qui captent la protéine traversant les membranes, favorisent sa rétraction dans les mitochondries, et contrôlent également le processus de repliement correct de la chaîne polypeptidique. La plupart des chaperons ont une activité ATPase, à la suite de quoi le transport des protéines dans les mitochondries et la formation de leurs formes fonctionnellement actives sont des processus dépendants de l'énergie.