Les antigènes et leurs propriétés. Propriétés de base et structure des antigènes
Lire aussi
1.1. CONCEPTS D'INFECTION ET DE MALADIE INFECTIEUSE
Montant de l'infection réactions biologiques, avec lequel le macro-organisme répond à l'introduction d'un agent microbien (infectieux) qui provoque une violation de la constance environnement interne(homéostasie).
Des processus similaires provoqués par des protozoaires sont appelés invasions.
Le processus complexe d'interaction entre les micro-organismes et leurs produits, d'une part, les cellules, tissus et organes humains et animaux, d'autre part, se caractérise par une très grande variété de ses manifestations. Les manifestations pathogéniques et cliniques de cette interaction entre micro-organismes et macro-organismes sont désignées par le terme maladie infectieuse (maladie).
Autrement dit, les notions de "maladie infectieuse" et d'"infection" ne sont absolument pas équivalentes, la maladie n'étant qu'une des manifestations de l'infection. Bien que même dans la littérature spécialisée à l'heure actuelle, le terme "infection" soit largement utilisé pour désigner les maladies infectieuses pertinentes. Par exemple, dans les expressions "infections intestinales", "infections transmises par voie aérienne". maladies infectieuses continuent de causer d'énormes dégâts à diverses espèces.
Ces dernières années, 38 nouvelles infections de maladies dites émergentes ont été enregistrées, dont le VIH, les fièvres hémorragiques, la maladie du légionnaire, hépatite virale, maladies à prions ; de plus, dans 40 % des cas, il s'agit de formes nosologiques qui étaient auparavant considérées comme non infectieuses.
Les caractéristiques des maladies infectieuses sont les suivantes :
- leur facteur étiologique est un agent microbien ;
- elles se transmettent du malade au bien-portant ;
- laisser un certain degré d'immunité;
- caractérisé par un flux cyclique ;
- ont un certain nombre de syndromes communs.
1.2. ÉTAPES CLINIQUES DES MALADIES INFECTIEUSES
Conformément à ces caractéristiques, toute maladie infectieuse a certains stades cliniques (périodes) de son évolution, exprimés à un degré ou à un autre :
- période d'incubation la période à partir du moment où un agent infectieux pénètre dans le corps humain jusqu'à ce que les premiers précurseurs de la maladie apparaissent. L'agent causal au cours de cette période n'est généralement pas isolé dans environnement, et le patient ne représente pas un danger épidémiologique pour les autres ;
- période prodromique manifestation des premiers symptômes non spécifiques de la maladie, caractéristique de l'intoxication générale du macro-organisme par les produits de l'activité vitale des micro-organismes et de l'action éventuelle des endotoxines bactériennes libérées lors de la mort de l'agent pathogène; ils ne sont pas non plus rejetés dans l'environnement;
La période de pointe de la maladie est la manifestation de symptômes spécifiques de la maladie. S'il existe un complexe symptomatique caractéristique au cours de cette période de développement de la maladie, les cliniciens appellent une telle manifestation de la maladie une infection manifeste et, dans les cas où la maladie survient sans symptômes prononcés pendant cette période, une infection asymptomatique. Cette période de développement d'une maladie infectieuse s'accompagne généralement de la libération de l'agent pathogène du corps, à la suite de quoi le patient représente un danger épidémiologique pour les autres; ces états sont caractérisés par une période de résultats. Pendant cette période, il est possible :
- rechute de la maladie retour des manifestations cliniques de la maladie sans réinfection due à des agents pathogènes restant dans l'organisme ;
- surinfection infection du macro-organisme par le même agent pathogène jusqu'à guérison. Si cela se produit après la guérison, on parlera de réinfection, car elle survient à la suite d'une nouvelle infection par le même agent pathogène (comme c'est souvent le cas avec la grippe, la dysenterie, la gonorrhée) ;
- bactérioporteur, ou plutôt porteur de microbes, porteur de l'agent causal de toute maladie infectieuse sans manifestations cliniques;
- récupération complète (reconvalescence) pendant cette période, les agents pathogènes sont également excrétés du corps humain en grande quantité, et les voies d'excrétion dépendent de la localisation du processus infectieux. Par exemple, avec une infection respiratoire du nasopharynx et de la cavité buccale avec de la salive et du mucus; avec des infections intestinales avec des matières fécales et de l'urine, avec des maladies pyoinflammatoires avec du pus;
- issue fatale. Dans le même temps, il convient de rappeler que les cadavres de patients infectieux sont soumis à une désinfection obligatoire, car ils représentent un certain risque épidémiologique en raison de leur forte teneur en agent microbien.
Dans la doctrine de l'infection, il y a aussi le concept de persistance (infection) : les micro-organismes pénètrent dans le corps d'un animal et peuvent y exister sans se manifester pendant un temps suffisamment long.
Cela se produit très souvent avec l'agent causal de la tuberculose.
La différence entre le portage bactérien et la persistance :
- lorsqu'il est transporté, l'animal libère l'agent pathogène dans l'environnement et est dangereux pour les autres ;
- pendant la persistance, les animaux infectés ne libèrent pas le micro-organisme dans l'environnement, par conséquent, ils ne sont pas épidémiologiquement dangereux pour les autres.
En plus des termes ci-dessus, il existe également le concept de "processus infectieux" - c'est la réponse du corps à la pénétration et à la circulation d'un agent microbien en son sein.
Dès la définition du concept d'"infection", les facteurs nécessaires à son apparition et à son développement deviennent évidents :
microorganisme pathogène;
Hôte susceptible;
environnement externe dans lequel ils interagissent.
1.3. PROPRIÉTÉS DES ANTIGÈNES
La réponse immunitaire est une réaction coopérative complexe à plusieurs composants système immunitaire organisme, induite par un antigène et visant son élimination. Le phénomène de réponse immunitaire sous-tend l'immunité.
La réponse immunitaire dépend de :
- propriétés antigéniques, composition, poids moléculaire, dose, fréquence d'exposition, durée de contact) ;
- conditions corporelles (réactivité immunologique);
- Conditions environnementales.
Antigènes
Initialement, le terme antigène (de l'anglais. Antibodi generator) était utilisé pour désigner toute molécule qui induit la formation d'anticorps spécifiques par les lymphocytes B. Cependant, le terme a désormais un sens plus large, désignant toute molécule pouvant être spécifiquement reconnue par des éléments du système immunitaire acquis, c'est-à-dire Cellules B ou cellules T, ou les deux.
L'antigène est l'initiateur et force motrice toutes les réponses immunitaires acquises. Le système immunitaire est né pour reconnaître et détruire les agents étrangers, ainsi que pour éliminer la source de leur formation - bactéries, cellules infectées par des virus, etc. Lorsque l'antigène est éliminé, la réponse immunitaire s'arrête.
Les antigènes sont des substances d'origines diverses qui portent des signes d'étrangeté génétique et provoquent le développement de réactions immunitaires (humorales, cellulaires, tolérance immunitaire, induction de la mémoire immunitaire).
Les propriétés d'un antigène sont déterminées par un ensemble de caractéristiques : immunogénicité, antigénicité, spécificité.
L'immunogénicité est la capacité d'un antigène à induire une réponse immunitaire dans l'organisme.
Antigénicité la capacité d'un antigène à interagir uniquement avec des anticorps homologues et des lymphocytes d'un certain clone.
Spécificité caractéristiques structurelles qui distinguent un antigène d'un autre.
La capacité de provoquer le développement d'une réponse immunitaire et de déterminer sa spécificité est possédée par un fragment d'une molécule d'antigène - un déterminant antigénique (épitope), qui réagit sélectivement avec les récepteurs et les anticorps reconnaissant l'antigène. Une molécule d'antigène peut avoir plusieurs épitopes, c'est-à-dire être polyvalente. Plus la molécule d'antigène est complexe et plus elle contient d'épitopes, plus elle est susceptible de développer une réponse immunitaire.
Les immunogènes ou antigènes complets sont des substances qui provoquent une réponse immunitaire complète et possèdent des propriétés : immunogénicité, antigénicité et spécificité. Les immunogènes sont des protéines biopolymères, leurs complexes avec des glucides (glycoprotéines), ainsi que des polysaccharides complexes, des lipopolysaccharides de haut poids moléculaire. Plus les organismes sont éloignés des humains en termes d'évolution, plus leurs protéines sont immunogènes.
Les haptènes sont des antigènes incomplets, des substances relativement simples capables de participer à des interactions immunologiques, mais incapables d'induire indépendamment une réponse immunitaire. Les haptènes sont antigéniques et spécifiques, mais non immunogènes.
Les haptènes, après s'être attachés à de grandes molécules généralement protéiques (porteurs), peuvent acquérir les propriétés d'un antigène complet.
Les tolérogènes sont des antigènes capables de supprimer les réactions immunitaires avec le développement d'une incapacité spécifique à y répondre.
Les antigènes sont des substances chimiques, libres ou présentes dans les cellules, qui peuvent induire une réponse immunitaire dans l'organisme.
Un antigène complet est composé de deux parties :
- support (partie stabilisatrice) 97 à 99 % de la molécule d'antigène ; ce sont, en règle générale, des macromolécules, des particules corpusculaires inertes ;
- les oligosaccharides ou oligopeptides du groupe déterminant (épitope), généralement situés à la surface de la molécule (epi); il peut y avoir plusieurs épitopes sur un support, à cet égard, le concept de densité d'épitopes est introduit ; le groupe déterminant détermine la spécificité de l'antigène.
Propriétés des antigènes :
- capable d'induire une réponse immunitaire ;
- capables d'interactions spécifiques avec diverses molécules et cellules (érythrocytes, etc.).
Si les deux sont mis en œuvre propriétés spécifiées, alors un tel antigène est dit complet, si seule la deuxième propriété est réalisée, alors un tel antigène est dit défectueux ou haptène.
L'haptène peut être fixé sur des adjuvants supports spéciaux. Mécanisme d'action des adjuvants :
- créer un dépôt d'antigènes;
- agrandir la molécule;
- activer le tissu lymphoïde.
Classification des antigènes :
- par l'étrangeté
- les xénoantigènes (hétéro) n'appartiennent pas aux individus de cette espèce ;
- les alloantigènes (homo) appartiennent aux individus de cette espèce ;
- les auto-antigènes possèdent des antigènes, par exemple les cellules «barrières» les spermatozoïdes, les cellules cérébrales; cellules vées à activité immunitaire;
- type de réponse immunitaire déclenchée
- immunogènes;
- allergènes;
- tolérogènes;
- les antigènes de transplantation ;
- en relation avec la glande thymus (thymus)
- T dépendant ;
- T indépendant.
- par localisation dans le micro-organisme
- Antigènes O lipopolysaccharides (LPS) paroi cellulaire, thermostable, hautement actif, diversifié dans différents micro-organismes et même dans le même ;
- protéine flagellaire de l'antigène H, thermolabile, assez active, également diversifiée;
- Antigènes K glycoprotéines capsulaires, l'immunogénicité dépend de la nature chimique ;
- les antigènes fimbriaux;
- les antigènes protoplasmiques ;
- exoallergènes;
- par spécificité pour le micro-organisme porteur
- espèce chez tous les individus de l'espèce;
- variante typique, pour vars ;
- groupe commun aux micro-organismes différents types et l'accouchement;
- mis en scène apparaissent à certains stades de développement;
- spécifique à la souche.
ANTIGÈNES DE MICROORGANISMES
La plupart des agents pathogènes maladies infectieuses humains, leurs structures et leurs toxines sont des antigènes à part entière qui provoquent le développement de réponses immunitaires.
ANTIGÈNES BACTÉRIENS
Les antigènes sont isolés selon leur localisation dans la cellule bactérienne :
Antigène capsulaire K Ag
Antigène flagellaire H Ag
Antigène somatique O Ag
Les OAg de la plupart des bactéries sont représentés par un complexe lipopolysaccharidepolypeptide thermostable; chez les bactéries gram-négatives, OAr est une endotoxine.
HAg est représenté par la protéine thermolabile flagelline.
Les KAg de la plupart des bactéries ont une nature polysaccharidique. Par sensibilité à la température, les KAg sont divisés en A, B et Lantigens. Les plus stables thermiquement sont l'AAr, qui peut résister à l'ébullition pendant plus de 2 heures. Les ВАг résistent au chauffage à une température de 60°С pendant une heure, et les LАг sont détruits lorsqu'ils sont chauffés à 60°С.
Pour identifier les micro-organismes isolés en laboratoire, une différenciation intraspécifique ou intragénérique des micro-organismes est utilisée, basée sur les différences de structure antigénique. Dans le même temps, la structure antigénique des bactéries est représentée symboliquement sous la forme d'une formule antigénique. Par exemple, la formule antigénique de l'un des sérovars d'E. coli qui cause la coli-entérite chez les jeunes jeunes animaux est désignée par O55:K5:H21 (un sérovar appartenant au sérogroupe O55).
Riz. 1. Antigènes de bactéries : Oantigène (3 parois cellulaires) ; Nantigen (7 flagelles); Kantigen (2 gélules).
ANTIGÈNES VIRUS
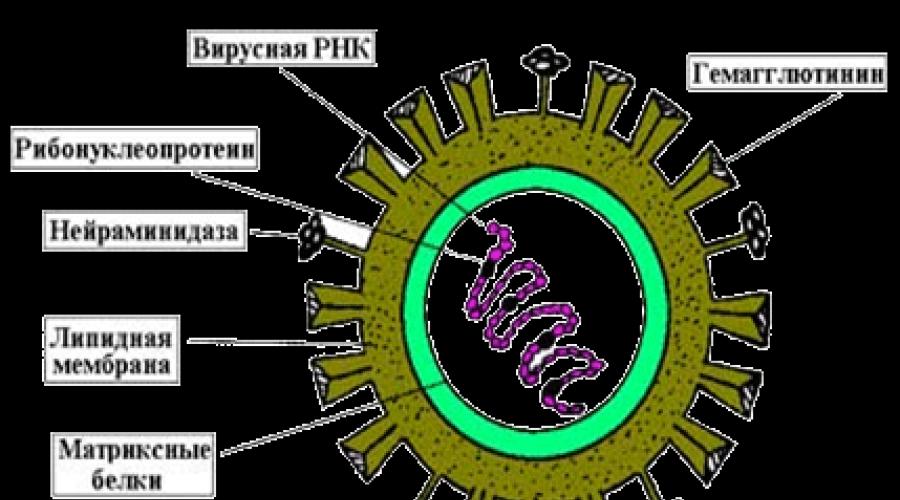
Chaque virion de tout virus contient différents antigènes. Certains d'entre eux sont spécifiques au virus. La composition d'autres antigènes comprend des composants de la cellule hôte (lipides, glucides), qui sont inclus dans son enveloppe externe. Les antigènes des virions simples sont associés à leurs nucléocapsides. Selon leur composition chimique, ils appartiennent aux ribonucléoprotéines ou aux désoxyribonucléoprotéines, qui sont des composés solubles et sont donc désignés sous le nom de Santigens (solutio solution). Dans les virions à organisation complexe, certains composants antigéniques sont associés aux nucléocapsides, d'autres aux glycoprotéines de l'enveloppe externe. De nombreux virions simples et complexes contiennent une hémagglutinine Vantigens de surface spéciale et l'enzyme neuraminidase.

Riz. 2. Antigènes des virus grippaux (surface (Vantigens) et core (Santigens)).

Riz. 3. Antigènes des virus de l'hépatite B (surface (Vantigens) et noyau (Santigens)).
ANTIGÈNES DU CORPS
Tous les tissus et cellules du corps ont des propriétés antigéniques. Certains antigènes sont spécifiques à tous les mammifères, d'autres sont spécifiques à l'espèce humaine, d'autres sont destinés à certains groupes, ils sont appelés isoantigènes (par exemple, les antigènes des groupes sanguins). Les antigènes qui sont uniques à un organisme donné comprennent les antigènes de compatibilité tissulaire.
isoantigènes
Les isoantigènes ou antigènes de groupe sont des antigènes par lesquels des individus individuels ou des groupes d'individus de la même espèce diffèrent les uns des autres.
Plusieurs dizaines d'isoantigènes ont été découverts dans les érythrocytes, les leucocytes, les plaquettes, ainsi que dans le plasma sanguin des personnes.
Les isoantigènes, génétiquement apparentés, sont combinés en groupes appelés: le système ABO, Rhésus, etc. La division des personnes en groupes selon le système ABO est basée sur la présence ou l'absence d'antigènes sur les érythrocytes, désignés A et B. Conformément à cela, toutes les personnes sont divisées en 4 groupes. Les antigènes du groupe I (O) sont absents, les érythrocytes du groupe II (A) contiennent l'antigène A, groupe III(B) les érythrocytes ont l'antigène B, les érythrocytes du groupe IV (AB) ont les deux antigènes. Puisqu'il existe des micro-organismes dans l'environnement qui ont les mêmes antigènes (on les appelle à réaction croisée), une personne a des anticorps contre ces antigènes, mais seulement contre ceux qu'elle n'a pas. Le corps tolère ses propres antigènes. Lorsque du sang ou des érythrocytes sont transfusés à un receveur dont le sang contient des anticorps dirigés contre l'antigène correspondant, une agglutination des érythrocytes incompatibles transfusés se produit dans les vaisseaux, ce qui peut provoquer un choc et la mort du receveur.
Chez certaines personnes, les globules rouges contiennent également un antigène spécial appelé antigène Rh. Par la présence ou l'absence du Rhantigène, les gens sont divisés en deux groupes rhésus (Rh) positif et rhésus (Rh) négatif. Lorsque le sang est transfusé à un receveur Rh négatif, si les érythrocytes du donneur contiennent l'antigène Rh, un ictère hémolytique peut se développer.

Riz. 4. Les récepteurs intégrés à la membrane érythrocytaire sont les antigènes de l'organisme (isoantigènes), notamment les antigènes A et B du système ABO et le facteur Rh.
Antigènes du complexe principal de compatibilité tissulaire (histo).
En plus des antigènes communs à toutes les personnes et des antigènes de groupe, chaque organisme possède un ensemble unique d'antigènes qui lui sont propres. Ces antigènes sont codés par un groupe de gènes situés sur le chromosome 6 chez l'homme et sont appelés antigènes du complexe majeur de compatibilité tissulaire et sont désignés antigènes MHC (complexe majeur d'histocompatibilité). Les antigènes humains MHC ont d'abord été découverts sur les leucocytes et portent donc un nom différent HLA (Human leucocyte antigens). Les antigènes MHC appartiennent aux glycoprotéines et sont contenus sur les membranes des cellules du corps, déterminant ses propriétés individuelles et induisant des réactions de transplantation, pour lesquelles ils ont reçu le troisième nom - antigènes de transplantation. De plus, les antigènes MHC jouent un rôle indispensable dans l'induction d'une réponse immunitaire à n'importe quel antigène.
Les protéines de classe I se trouvent à la surface de presque toutes les cellules du corps. Les antigènes de classe I assurent la présentation des antigènes aux lymphocytes CD8+ cytotoxiques, et la reconnaissance de cet antigène par les cellules présentatrices d'antigène d'un autre organisme lors de la transplantation conduit au développement de l'immunité de transplantation.
Les antigènes du CMH de classe II se trouvent principalement sur les cellules dendritiques présentatrices d'antigène, les macrophages et les lymphocytes B. Le rôle principal dans l'immunogénèse des antigènes de classe II est la participation à la présentation d'antigènes étrangers aux lymphocytes T auxiliaires.
Spécificité - c'est la capacité d'un antigène à interagir avec des anticorps strictement définis ou des récepteurs antigéniques des lymphocytes.
Dans ce cas, l'interaction ne se produit pas avec toute la surface de l'antigène, mais uniquement avec sa petite surface, appelée "déterminant antigénique" ou "épitope". Une molécule d'antigène peut avoir de plusieurs unités à plusieurs centaines d'épitopes de spécificité différente. Le nombre d'épitopes détermine la valence de l'antigène. Par exemple: l'albumine d'œuf (M 42 000) a 5 épitopes, c'est-à-dire 5-valent, la protéine thyroglobuline (M 680 000) est 40-valente.
Dans les molécules de protéines, un épitope (déterminant antigénique) est formé par une combinaison de résidus d'acides aminés. La taille des déterminants antigéniques des protéines peut comprendre de 5 à 7 à 20 résidus d'acides aminés. Les épitopes qui sont reconnus par les récepteurs antigéniques des lymphocytes B et T ont leurs propres caractéristiques.
Les épitopes de cellules B de type conformationnel (formés par des résidus d'acides aminés de différentes parties de la molécule de protéine, mais contigus dans la configuration spatiale du globule protéique) sont situés sur la surface externe de l'antigène, formant des boucles et des saillies. Typiquement, le nombre d'acides aminés ou de sucres dans un épitope est compris entre 6 et 8. Les récepteurs de reconnaissance d'antigène sur les cellules B reconnaissent la conformation native de l'épitope et non la séquence linéaire des résidus d'acides aminés.
Les épitopes des lymphocytes T sont une séquence linéaire de résidus d'acides aminés qui font partie de l'antigène et comprennent un plus grand nombre de résidus d'acides aminés que les épitopes des lymphocytes B. Leur reconnaissance ne nécessite pas de sauvegarder la configuration spatiale.
Immunogénicité - la capacité de l'antigène à provoquer une défense immunitaire du macro-organisme. Le degré d'immunogénicité est déterminé par les facteurs suivants :- étrangeté . Pour qu'une substance agisse comme un immunogène, elle doit être reconnue comme "non-soi". Plus l'antigène est étranger, c'est-à-dire moins il ressemble aux structures de l'organisme, plus la réponse immunitaire qu'il provoque est forte. Par exemple, la synthèse d'anticorps dirigés contre l'albumine sérique bovine est plus facile à induire chez un lapin que chez une chèvre. Les lapins appartiennent à l'ordre des lagomorphes et sont plus éloignés dans le développement phylogénétique de la chèvre et du taureau, qui appartiennent aux artiodactyles.
- Nature de l'antigène . Les immunogènes les plus puissants sont les protéines. Les polysaccharides purs, les acides nucléiques et les lipides ont de faibles propriétés immunogènes. Dans le même temps, les lipopolysaccharides, les glycoprotéines, les lipoprotéines sont capables d'activer suffisamment le système immunitaire.
- Masse moléculaire . Ceteris paribus, un grand poids moléculaire de l'antigène fournit une plus grande immunogénicité. Les antigènes sont considérés comme de bons immunogènes si leur poids moléculaire est supérieur à 10 kD. Plus le poids moléculaire est élevé, plus il y a de sites de liaison (épitopes), ce qui entraîne une augmentation de l'intensité de la réponse immunitaire.
- Solubilité. Les antigènes corpusculaires associés aux cellules (érythrocytes, bactéries) sont généralement plus immunogènes. Les antigènes solubles (albumine sérique) peuvent également être hautement immunogènes, mais ils sont éliminés plus rapidement. Pour augmenter la durée de leur séjour dans l'organisme, nécessaire au développement d'une réponse immunitaire efficace, des adjuvants (substances déposantes) sont utilisés. Les adjuvants sont des substances utilisées pour renforcer la réponse immunitaire, par exemple la paraffine liquide, la lanoline, l'hydroxyde et le phosphate d'aluminium, l'alun de potassium, le chlorure de calcium, etc.
- La structure chimique de l'antigène . L'augmentation du nombre d'acides aminés aromatiques dans les polypeptides synthétiques augmente leur immunogénicité. A poids moléculaire égal (environ 70 000), l'albumine est un antigène plus puissant que l'hémoglobine. Dans le même temps, la protéine de collagène, dont le poids moléculaire est 5 fois supérieur à celui de l'albumine, et est de 330 000, a une immunogénicité significativement plus faible par rapport à l'albumine, ce qui est sans aucun doute lié aux caractéristiques structurelles de ces protéines.
Antigènes du complexe majeur d'histocompatibilité.
Les antigènes sont des substances génétiquement étrangères qui, lorsqu'elles pénètrent dans le milieu interne de l'organisme ou se forment dans l'organisme, provoquent une réaction immunologique spécifique, se manifestant par la synthèse d'anticorps, l'apparition de lymphocytes sensibilisés ou l'émergence d'une tolérance à ce substance, hypersensibilité de type immédiat et retardé, mémoire immunologique.
Propriétés des antigènes: spécificité (antigénicité), immunogénicité.
antigénicité est la capacité d'un antigène à induire une réponse immunitaire dans l'organisme.
Immunogénicité est la capacité d'un antigène à former une immunité.
Spécificité - c'est la capacité d'un antigène à interagir sélectivement uniquement avec des anticorps complémentaires ou des récepteurs reconnaissant Ag des lymphocytes T d'un certain clone.
La spécificité des antigènes est déterminée par les caractéristiques structurelles de la macromolécule - la présence et la nature des épitopes.
Un épitope (déterminant antigénique) est une partie d'une molécule d'antigène qui interagit avec un site actif d'un anticorps ou d'un récepteur de cellule T. Un épitope est constitué de résidus d'acides aminés. Le nombre d'épitopes détermine la valence de l'antigène.
NatureAg. Les antigènes sont des biopolymères naturels ou synthétiques avec une structure assez rigide et un poids moléculaire élevé. Ce sont des protéines et leurs complexes avec des glucides (glycoprotéines), des lipides (lipoprotéines), des acides nucléiques (nucléoprotéines).
Les protéines en tant que biopolymères avec une étrangeté génétique prononcée ont les propriétés antigéniques les plus prononcées. Plus les animaux sont en relation phylogénétique, plus leurs protéines sont antigéniques les unes par rapport aux autres. Cette propriété des protéines est utilisée pour identifier la relation phylogénétique des animaux de différentes espèces, en médecine légale (détermination des espèces de taches de sang) et dans l'industrie alimentaire (pour détecter la falsification des produits carnés).
La sévérité des propriétés antigéniques est associée à:
Ø poids moléculaire ;
Ø solubilité (état colloïdal), par exemple, la kératine est une protéine de haut poids moléculaire, mais ne peut se présenter sous forme de solution colloïdale et n'est donc pas un antigène ;
Ø voie d'introduction dans le corps (les propriétés antigéniques de certains antigènes se manifestent mieux lorsqu'ils sont administrés par voie orale, d'autres - par voie intradermique, d'autres - par voie intramusculaire, etc.;
Ø le taux de leur métabolisme (destruction) dans l'organisme.
Valeur de poids moléculaire . Les polysaccharides n'ont des propriétés antigéniques qu'avec un poids moléculaire d'au moins 600 000. Les protéines ont une antigénicité avec un poids moléculaire supérieur à 5 000 - 10 000 (5 -10 kD). Les antigènes faibles sont des composés de haut poids moléculaire collagène, gélatine, protamines (ils ont un petit poids moléculaire). Mais il y a des exceptions à cette règle :
L'hormone pancréatique porcine - l'insuline d'un poids moléculaire de 3,8 kD a une antigénicité, le dextrane de substitut sanguin d'un poids moléculaire de 100 kD n'est pas un antigène;
Les acides nucléiques ont un poids moléculaire plus élevé mais moins antigénique que les protéines.
Lors de la dénaturation (coagulation) par haute température, les acides, les alcalis, les protéines perdent leurs propriétés antigéniques.
La valeur de la dose d'Ag. Plus la dose d'antigène est élevée, plus la réponse immunitaire est prononcée. Cependant, si la dose d'antigène est trop élevée, une tolérance immunologique peut se produire, c'est-à-dire absence de réponse du corps à l'irritation antigénique. Ce phénomène s'explique par la stimulation d'une sous-population de T-suppresseurs par l'antigène.
La valeur du taux de métabolisme de l'antigène dans le corps. Les polypeptides constitués d'acides aminés D sont lentement et incomplètement dégradés par les enzymes corporelles et ne sont pas Ag, contrairement aux polypeptides construits à partir d'acides aminés L, qui sont activement métabolisés dans le corps.
TYPES D'ANTIGÈNES :
1. Exogène, endogène ;
2. Complète et inférieure (haptènes, semi-haptènes) ;
3. Dépendant du thymus et indépendant du thymus ;
4. Superantigènes ;
5. Hétérogène ;
6. Autoantigènes ;
7. Tumeur ;
8. Bactérien (spécifique au groupe, spécifique à l'espèce, spécifique au type, O-, K-, H-antigènes et autres);
9. virale ;
10. Fongique ;
11. Protecteur ;
12. Isoantigènes ;
13. Antigènes du complexe majeur d'histocompatibilité.
antigènes exogènes- pénètrent dans l'organisme depuis l'environnement, subissent une endocytose et un clivage dans les cellules présentatrices d'Ag (macrophages, cellules dendritiques du thymus, cellules du processus folliculaire des ganglions lymphatiques et de la rate, cellules M des follicules lymphatiques du tube digestif, cellules de Langerhans du peau). Ensuite, l'Ag-déterminant (épitope), en complexe avec la molécule du CMH de classe II, est incorporé dans la membrane plasmique de la cellule présentatrice d'Ag et présenté aux lymphocytes T CD 4+ (T-helpers) ;
antigènes endogènes sont des produits des propres cellules du corps. Le plus souvent, il s'agit de protéines anormales de cellules tumorales et de protéines virales synthétisées par des cellules hôtes infectées par le virus. Leurs déterminants antigéniques (épitopes) sont présentés en association avec la molécule du CMH de classe I aux lymphocytes T CD 8+ (T-killers).
Ag complet– avoir la capacité d'induire la formation d'anticorps et d'interagir avec eux ;
Ag défectueux (haptènes)- des substances de faible poids moléculaire qui n'ont pas la capacité d'induire la formation d'anticorps et, mais interagissent avec des anticorps spécifiques prêts à l'emploi. Les haptènes acquièrent les propriétés d'antigènes à part entière lorsqu'ils sont liés à des substances macromoléculaires, telles que des protéines (schleppers). Les haptènes sont médicaments, par exemple, des antibiotiques capables de déclencher une réponse immunitaire lorsqu'ils sont liés à des protéines corporelles (albumine), ainsi que des protéines à la surface des cellules (érythrocytes, leucocytes). En conséquence, des anticorps se forment qui peuvent interagir avec l'haptène. Avec l'introduction répétée de l'haptène dans le corps, une réponse immunitaire secondaire se produit, souvent sous la forme d'une réaction allergique, telle que l'anaphylaxie ;
Semi-haptènes- substances inorganiques - iode, brome, chrome, nickel, groupe nitro, azote, etc. - en se liant aux protéines, par exemple, de la peau, ils peuvent provoquer une dermatite allergique de contact (THS), qui se développe lorsque la peau entre en contact de manière répétée avec des objets chromés, nickelés, que de l'iode est appliqué sur la peau, etc.
Antigènes thymo-dépendants - ce sont des antigènes qui nécessitent la participation des lymphocytes T pour induire une réponse immunitaire, ces antigènes sont majoritaires ;
Indépendant du thymus - des antigènes capables de stimuler la synthèse d'anticorps sans l'aide de lymphocytes T, par exemple, le LPS de parois cellulaires bactériennes, des polymères synthétiques de haut poids moléculaire.
Superantigènes(entérotoxines bactériennes (staphylocoques, choléra), certains virus (rotavirus), etc. - un groupe particulier d'antigènes qui, à des doses beaucoup plus faibles que d'autres antigènes, provoquent l'activation polyclonale et la prolifération d'un grand nombre de lymphocytes T (plus de 20 %, tandis que les antigènes ordinaires stimulent 0,01 % des lymphocytes T.) Cela produit beaucoup d'IL-2 et d'autres cytokines qui provoquent une inflammation et des lésions tissulaires.
Ag hétérogène- ce sont des antigènes à réaction croisée, des antigènes communs dans diverses sortes microbes, animaux et humains. Ce phénomène est appelé mimétisme antigénique. Par exemple, streptocoques hémolytiques du groupe A contiennent des antigènes à réaction croisée (en particulier, la protéine M) qui sont communs avec les antigènes de l'endocarde et des glomérules des reins humains. Ces antigènes bactériens provoquent la formation d'anticorps qui interagissent avec les cellules humaines, conduisant au développement de rhumatismes et de glomérulonéphrites post-streptococciques. L'agent causal de la syphilis il existe des antigènes phospholipidiques similaires aux phospholipides cardiaques humains et animaux, de sorte que l'antigène cardiolipine cardiaque bovin est utilisé pour détecter les anticorps anti-treponema pallidum dans le sérodiagnostic de la syphilis (réaction de Wassermann). Antigène de Forsman - détecté dans les érythrocytes de moutons, chats, chiens, reins de cobayes, salmonelles.
Autoantigènes sont des antigènes endogènes qui provoquent la production d'auto-anticorps. Distinguer:
- primaire naturel(tissu normal du cristallin de l'œil, tissu nerveux, etc.), qui est associé à une violation de l'autotolérance,
Secondaire acquis - produits de lésions tissulaires par des microbes, des virus, des brûlures, des radiations, du froid, qui proviennent de leurs propres tissus à la suite de modifications des tissus lors de brûlures, de gelures, sous l'action de radiations radioactives.
Tumeur (oncoantigènes, antigènes T (tumeur - tumeur)- à la suite de la transformation maligne des cellules normales en cellules tumorales, elles commencent à exprimer (manifester) des antigènes anormaux spécifiques absents des cellules normales. La détection des antigènes tumoraux par des méthodes immunologiques permettra un diagnostic précoce des maladies oncologiques.
Antigènes bactériens :
- spécifique au groupe - des antigènes communs à différentes espèces d'un même genre ou d'une même famille,
- spécifique à l'espèce - les antigènes caractéristiques des membres d'une même espèce,
- spécifique au type - déterminer les variants sérologiques (sérovars, sérotypes) au sein d'une même espèce,
- Antigènes H (flagellés) - la protéine flagelline, qui fait partie des flagelles bactériens, est thermolabile ;
- O-antigènes (somatiques) - est un LPS de gr-bactéries, thermostable. Les épitopes de l'antigène somatique sont représentés par des hexoses (galactorse, rhamnose, etc.) et des sucres aminés (N-acétylglucosamine, N-acétylgalactosamine). Chez les bactéries Gr+, l'antigène somatique est représenté par les acides glycéryltéichoïque et ribitoltéichoïque.
- Antigènes K (antigènes capsulaires) - sont situés dans la capsule et sont associés à la couche superficielle de lipopolysaccharide de la paroi cellulaire. Ils contiennent des polysaccharides acides, qui comprennent les acides galacturonique, glucuronique et iduronique. Les antigènes capsulaires sont utilisés pour préparer des vaccins contre les méningocoques, les pneumocoques, Klebsiella. Cependant, l'introduction de fortes doses d'antigènes polysaccharidiques peut entraîner une tolérance. Chez Escherichia coli, l'antigène K est divisé en fractions A (thermostable), B, L (thermolabile). Une variation de l'antigène K est l'antigène Vi de surface (chez Salmonella), qui détermine la virulence du microbe et la persistance de l'agent pathogène dans les bactéries porteuses.
- Les antigènes des bactéries sont aussi leurs toxines, ribosomes, enzymes.
Viral- a) supercapside (protéine et glycoprotéine, par exemple, hémagglutinine et neuraminidase du virus de la grippe), b) capside (protéine), c) noyau (nucléoprotéine).
fongique– les champignons ressemblant à des levures Candida albicans contiennent un polysaccharide de la paroi cellulaire – mannane, protéines cytoplasmiques et nucléaires. Parmi eux, 80 antigènes ont été identifiés. Ces antigènes provoquent des réactions et une sensibilisation immédiates (anticorps des classes Ig m, Ig G, Ig A, Ig E) et retardées (cellules T) sans manifestations cliniques. Les antigènes fongiques ont des effets immunostimulateurs et immunosuppresseurs.
Protecteur- ce sont des déterminants antigéniques (épitopes) de micro-organismes qui provoquent la réponse immunitaire la plus forte, qui confère une immunité au pathogène correspondant en cas d'infection répétée. Pour la première fois, ils ont été trouvés dans l'exsudat du tissu atteint d'anthrax. Les peptides les plus immunogènes et protecteurs des virus sont utilisés pour créer des vaccins synthétiques.
isoantigènes- les antigènes par lesquels les individus de la même espèce diffèrent les uns des autres (par exemple, les antigènes érythrocytaires - le système de groupe sanguin ABO, le facteur Rh, les antigènes leucocytaires - le principal complexe d'histocompatibilité).
Antigènes du complexe majeur d'histocompatibilité– glycoprotéines membranes cellulaires qui joue rôle important dans la réponse immunitaire, les réactions de rejet de greffe déterminent la prédisposition à certaines maladies. Le spectre des molécules du complexe majeur d'histocompatibilité est unique pour chaque organisme et détermine son individualité biologique, ce qui permet de distinguer les "propres" (histocompatibles) des "étrangers" (incompatibles). Le complexe majeur d'histocompatibilité est appelé CMH (complexe majeur d'histocompatibilité). Les antigènes du CMH chez différentes espèces animales sont désignés différemment: chez la souris - le système H2, chez le chien - DLA, chez le lapin - RLA, chez le porc - SLA. Chez l'homme, les antigènes du complexe majeur d'histocompatibilité sont appelés HLA (Human leucocyte antigenes), car à des fins cliniques et expérimentales, les antigènes leucocytaires sont définis comme des antigènes du complexe majeur d'histocompatibilité. Les antigènes leucocytaires humains sont codés par des gènes situés sur le 6e chromosome. Selon la structure chimique et le but fonctionnel, HLA est divisé en deux classes.
Antigènes l classe MHC présent à la surface de toutes les cellules nucléées. Ils régulent l'interaction entre les lymphocytes T tueurs et les cellules cibles. Principal rôle biologique antigènes de classe l, c'est qu'ils sont des marqueurs des "leurs propres". Les cellules porteuses d'antigènes de classe l ne sont pas attaquées par leurs propres tueurs T car, dans l'embryogenèse, les tueurs T autoréactifs qui reconnaissent les antigènes de classe l sur leurs propres cellules sont détruits. Les antigènes de classe l interagissent avec la molécule CD 8 sur la membrane T-killer.
Antigènes du CMH de classe II sont situés principalement sur la membrane des cellules immunocompétentes (macrophages, monocytes, lymphocytes B et T activés).Les antigènes de classe II interagissent avec la molécule CD 4 de la membrane T-helper, ce qui provoque la libération de lymphokines qui stimulent la prolifération et maturation des T-killers et des plasmocytes.
La détermination des antigènes HLA est nécessaire dans les situations suivantes :
Þ Lors du typage des tissus afin de sélectionner un donneur pour le receveur ;
Þ Établir un lien entre la présence de certains antigènes du CMH et la prédisposition à une maladie particulière. La corrélation la plus prononcée a été trouvée entre la présence de HLA-B27 et la maladie de Bechterew (spondylarthrite ankylosante) : 95 % des patients ont cet antigène.
Þ Lors de l'évaluation statut immunitaire(détection de porteurs d'antigènes HLA-DR a) lymphocytes T activés et b) cellules mononucléaires impliquées dans la reconnaissance des antigènes.
Distinguer les antigènes complets et incomplets, ou haptènes. Ces dernières sont des substances relativement simples qui peuvent participer à des interactions immunologiques, mais ne sont pas capables d'activer l'APC et d'induire indépendamment une réponse immunitaire. Ce n'est qu'après s'être attaché à de grosses molécules, généralement protéiques (porteurs), que l'haptène peut acquérir les propriétés d'un antigène complet.
Les biopolymères ont des propriétés antigéniques - les protéines, leurs complexes avec des glucides (glycoprotéines), des lipides (lipoprotéines), des acides nucléiques (nucléoprotéines), ainsi que des polysaccharides complexes, des lipopolysaccharides. Pour la manifestation des propriétés antigéniques, la taille de la molécule est importante. Les molécules d'un poids moléculaire supérieur à 10 000 sont généralement antigéniques et, avec un poids moléculaire inférieur, elles ont plus souvent les propriétés des haptènes. Les polysaccharides sont antigéniques à un poids moléculaire supérieur à 100 000. Les polypeptides constitués d'acides L-aminés sont antigéniques, et ceux constitués d'acides D-aminés n'ont pas cette propriété. Les protéines perdent leurs propriétés antigéniques lors de la dénaturation. Par exemple, les protéines coagulées par ébullition, traitées avec des solutions fortes d'acides ou d'alcalis, cessent d'être des antigènes. Manifestations de l'action antigénique associées à la destruction catabolique des antigènes dans le corps. Ainsi, les polypeptides D sont lentement et incomplètement détruits par les enzymes du corps et ne présentent pas de propriétés antigéniques.
Presque tous les substrats naturels ayant des propriétés antigéniques sont des complexes de plusieurs antigènes. Il sera montré ci-dessous qu'une cellule microbienne a une variété d'antigènes caractéristiques de ses structures individuelles. Même des molécules individuelles peuvent avoir plusieurs antigènes.
Les propriétés principales de l'antigène sont : la spécificité, l'étrangeté, l'immunogénicité ou la tolérogénicité.
Spécificité. La spécificité antigénique est un phénomène biologique unique qui sous-tend les interactions immunologiques dans le corps, ainsi que les méthodes de laboratoire pour la détermination de divers antigènes, le sérodiagnostic, les méthodes de prévention et de traitement spécifiques des maladies infectieuses.
Une structure qui a une spécificité antigénique individuelle est appelée déterminant antigénique ou épitope. Ce dernier nom reflète le fait que seules les structures situées à la surface de la molécule ont une activité antigénique, tandis que les structures profondes ne montrent une antigénicité que lorsque la conformation change ou que la molécule est détruite. La diversité des épitopes protéiques est obtenue grâce à une mosaïque de résidus d'acides aminés situés sur la surface globulaire de la molécule protéique.
Les épitopes qui déterminent l'antigénicité d'une molécule de protéine sont constitués de 625 acides aminés et sont situés dans Différents composants molécules, séparant les structures non antigéniques. Dans ce cas, les épitopes d'une même molécule ne doivent pas nécessairement avoir la même composition et la même spécificité. Le nombre d'épitopes identiques par molécule détermine le nombre de molécules d'anticorps qui peuvent la rejoindre, c'est-à-dire la valence d'un substrat antigénique donné. La valence des antigènes augmente avec leur poids moléculaire. Ainsi, la valence de l'ovalbumine d'un poids moléculaire de 45 000 est de 5 et la valence de l'hémocyanine d'un mol. pesant 6,5 millions - 231. Un épitope séparé d'une molécule ne peut avoir qu'une seule valence et avoir les propriétés d'un haptène, et la molécule entière pour cet épitopaptène joue le rôle de porteur.
Étant donné que les épitopes qui déterminent les propriétés antigéniques de la molécule sont situés dans certaines zones et que les propriétés toxiques des toxines microbiennes déterminent d'autres zones, il est possible de préparer des molécules d'anatoxine dépourvues de propriétés toxiques, mais conservant les propriétés antigéniques. Les anatoxines servent de base aux préparations vaccinales pour créer une immunité antitoxique.
L'étrangeté. Un antigène provoque une réponse immunitaire positive (formation d'anticorps et de lymphocytes actifs) uniquement dans les cas où il est étranger, c'est-à-dire a des structures qui sont absentes dans cet organisme. Le corps tolère ses propres antigènes. Ce n'est qu'avec des changements qui donnent à l'antigène des signes d'étrangeté qu'il acquiert la capacité d'induire une réponse immunitaire positive.
La structure des antigènes reflète la proximité évolutive des organismes qui les possèdent. Il existe des antigènes communs caractéristiques des représentants de différentes familles, genres, espèces. Il existe des antigènes variants qui sont différents pour les individus d'une même espèce. La détermination de la composition antigénique est utilisée pour la classification différents groupesêtres vivants et d'identifier les relations évolutives entre eux.
Au cours de l'évolution, les micro-organismes qui infectent les humains et les animaux acquièrent des antigènes similaires à ceux de l'hôte, ce que l'on appelle le mimétisme antigénique. Cela contribue au fait qu'il n'y a pas de réaction immunologique à de tels antigènes pendant longtemps et que les micro-organismes ont une chance supplémentaire de survivre dans l'organisme hôte, car ils ne sont pas reconnus comme étrangers. Les antigènes étrangers qui ont des structures similaires aux antigènes hôtes sont appelés antigènes à réaction croisée (ARC). Cependant, étant donné que les PRA sont associés à d'autres antigènes hautement immunogènes pour l'organisme, une réponse immunitaire à ceux-ci peut se produire. Dans ce cas, les anticorps humoraux et cellulaires résultants entrent en contact avec les antigènes de l'hôte et peuvent provoquer un processus immunopathologique. On sait que certaines souches de streptocoques hémolytiques peuvent avoir une PRA avec des antigènes de l'endocarde, des glomérules rénaux et du tissu nerveux humain, ce qui contribue au développement de rhumatismes, de glomérulonéphrite et de chorée. En conséquence, le virus de la rougeole a un PRA avec la protéine basique de la myéline, et la réponse immunitaire contribue à la démyélinisation des fibres nerveuses et au développement de la sclérose en plaques.
Les antigènes du système nerveux, des yeux, des organes reproducteurs sont séparés de l'environnement interne par des barrières physiologiques. Leurs antigènes n'induisent pas une tolérance totale et ne provoquent pas corps sain réaction auto-immune, car ils ne pénètrent pas dans les organes de l'immunogénèse. Ces antigènes sont appelés antigènes barrières. En cas de dommages aux barrières dus à une blessure ou à une maladie, les antigènes de la barrière pénètrent dans la circulation générale et peuvent provoquer un processus immunopathologique.
Les antigènes propres à l'organisme peuvent subir des modifications sous l'action de facteurs chimiques ou physiques externes ou entrer en contact avec des substances étrangères de nature haptique. En conséquence, des antigènes sont formés, dont la partie haptène est une structure étrangère, et le porteur est son propre antigène. Ces antigènes modifiés provoquent souvent des réactions allergiques.
Immunogénicité et tolérogénicité- propriétés alternatives de chaque substrat antigénique. Pour induire une réponse immunitaire et une tolérance, il est nécessaire d'influencer l'antigène sur un lymphocyte qui possède des récepteurs pour cet antigène - une cellule réactive à l'antigène (ARC). Les différences sont que lors de l'induction d'une réponse immunitaire positive, les ARC reçoivent des stimuli de cytokines, qui assurent leur prolifération et la formation d'un clone de cellules effectrices. Lors de l'induction de la tolérance immunologique, l'ARK n'est plus stimulée et meurt ou perd ses récepteurs pour l'antigène.
Antigène(du grec. anti- contre et génos- créer) est un biopolymère de nature organique, génétiquement étranger à un macro-organisme, qui, lorsqu'il pénètre dans celui-ci, est reconnu par son système immunitaire et provoque des réactions immunitaires visant à l'éliminer.
Propriétés des antigènes :
antigénicité - la capacité potentielle d'une molécule antigénique à activer des composants du système immunitaire et à interagir spécifiquement avec des facteurs immunitaires (anticorps, clone de lymphocytes effecteurs) ;
immunogénicité - la capacité potentielle d'un antigène à provoquer une réaction protectrice spécifique (réponse immunitaire) vis-à-vis de lui-même dans le macroorganisme.
Le degré d'immunogénicité dépend des caractéristiques moléculaires de l'antigène (nature, composition chimique, poids moléculaire, structure), clairance de l'antigène dans l'organisme, réactivité des macro-organismes.
spécificité - la capacité d'un antigène à induire une réponse immunitaire contre un épitope strictement défini.
La capacité de provoquer le développement d'une réponse immunitaire et de déterminer sa spécificité est possédée par un fragment de la molécule d'Ag ¾ du déterminant antigénique (épitope), qui réagit sélectivement avec les récepteurs reconnaissant l'Ag et l'Ab.
Classement d'origine :
exogène (provenant de l'extérieur du corps);
endogène (provenant de l'intérieur du corps) antigènes.
– autogène Les ¾ sont des antigènes endogènes, des molécules structurellement inchangées de son propre corps, synthétisées dans des conditions physiologiques. Normalement, les auto-antigènes ne provoquent pas de réaction du système immunitaire en raison de la tolérance immunologique formée (immunité) ou de leur inaccessibilité au contact avec des facteurs immunitaires - ce sont les antigènes dits barrières (cerveau, cristallin, follicules glande thyroïde, tubules séminifères des testicules). Lorsque la tolérance est rompue ou que l'intégrité des barrières biologiques est violée (la cause la plus fréquente est un traumatisme), les composants du système immunitaire commencent à réagir spécifiquement aux auto-antigènes en produisant des facteurs d'immunité spécifiques (auto-anticorps, un clone de lymphocytes autoréactifs).
– néoantigènes (tumeur)¾ sont des antigènes endogènes qui apparaissent dans le corps à la suite de mutations. Après modification, les molécules acquièrent les caractéristiques d'étrangeté.
Classement par nature : les biopolymères de nature protéique (protéines) et non protéique (polysaccharides, lipides, lipopolysaccharides, acides nucléiques, etc.).
Classification par structure moléculaire :
globulaire (la molécule a forme sphérique);
fibrillaire (forme de fil).
Classification selon le degré d'immunogénicité :
antigènes complets ¾ ont une antigénicité et une immunogénicité prononcées - le système immunitaire d'un organisme sensible réagit à leur introduction par le développement de facteurs immunitaires. En règle générale, ces substances ont un poids moléculaire suffisamment important (plus de 10 kDa), une grande taille de molécule (particule) sous la forme d'un globule et interagissent bien avec les facteurs d'immunité;
antigènes défectueux , ou haptènes ¾ ne sont pas capables lorsqu'ils sont introduits dans conditions normales induisent une réponse immunitaire dans le corps, car ils ont une immunogénicité extrêmement faible. Cependant, ils n'ont pas perdu leur propriété d'antigénicité, ce qui leur permet d'interagir spécifiquement avec des facteurs immunitaires prêts à l'emploi (anticorps, lymphocytes). Le plus souvent, les haptènes sont des composés de bas poids moléculaire (poids moléculaire inférieur à 10 kDa). Lorsqu'un haptène est associé à une molécule protéique, par exemple des albumines sériques sanguines, le conjugué résultant possède toutes les propriétés d'un antigène à part entière et, lorsqu'il est introduit dans l'organisme, provoque la production d'anticorps ou d'un clone de lymphocytes spécifiques à la partie haptène du complexe. Dans ce cas, la spécificité dans la composition de la molécule conjuguée est déterminée par la partie haptène et l'immunogénicité est déterminée par la protéine porteuse. La molécule de protéine porteuse est appelée schlepper (de lui. schlepper- remorqueur ).
Classification selon le degré d'étrangeté :
antigènes xénogéniques (hétérologue) - commun pour les organismes à différents stades de développement évolutif, par exemple, appartenant à différents genres et espèces. Un exemple serait l'antigène polysaccharidique de Forsman présent dans les érythrocytes des reins de chats, de chiens, de moutons et de cobayes ;
antigènes allogéniques (groupe) - commun aux organismes génétiquement non apparentés, mais appartenant à la même espèce. Sur la base des alloantigènes, la population générale d'organismes peut être divisée en groupes distincts. Un exemple de tels antigènes chez l'homme sont les antigènes sanguins (systèmes ABO, HLA, etc.). Les tissus allogéniques sont immunologiquement incompatibles lors de la transplantation - ils sont rejetés ou lysés par le receveur. Les microbes peuvent être subdivisés en sérogroupes basés sur des antigènes de groupe, qui ont grande importance pour diagnostic microbiologique(par exemple, la classification de Salmonella Kaufman-White);
antigènes isogéniques (individuel) - commun uniquement pour les organismes génétiquement identiques, par exemple pour les jumeaux identiques, les lignées consanguines d'animaux. Les isogreffes ont une compatibilité immunologique presque complète et ne sont pas rejetées par le receveur lors de la transplantation. Un exemple de tels antigènes dans la population humaine sont les antigènes d'histocompatibilité et, dans les bactéries, les antigènes de type qui ne donnent pas de clivage supplémentaire.
Classification selon le sens d'activation et la fourniture de la réponse immunitaire :
immunogènes lorsqu'ils sont ingérés, ils sont capables d'induire une réaction protectrice productive du système immunitaire, qui se termine par la production de facteurs immunitaires (anticorps, clones de lymphocytes réactifs aux antigènes). Dans la pratique clinique, les immunogènes sont utilisés pour l'immunodiagnostic, l'immunothérapie et l'immunoprophylaxie de nombreuses pathologies ;
tolérogène est l'exact opposé d'un immunogène. Lorsqu'il interagit avec le système d'immunité acquise, il provoque l'inclusion de mécanismes alternatifs conduisant à la formation d'une tolérance immunologique ou d'une non-réponse aux épitopes de ce tolérogène. Le tolérogène, en règle générale, se caractérise par un monomérisme, un faible poids moléculaire, une densité élevée d'épitopes et une dispersion élevée (non-agrégation) des solutions colloïdales. Les tolérogènes sont utilisés pour prévenir et traiter les conflits immunologiques et les allergies en induisant une non-réponse artificielle aux antigènes individuels ;
allergène ¾ l'effet qu'il produit, contrairement à l'immunogène, forme une réaction pathologique de l'organisme sous forme d'hypersensibilité de type immédiat ou retardé. Un allergène ne diffère pas dans ses propriétés d'un immunogène. En pratique clinique, les allergènes sont utilisés pour diagnostiquer les maladies infectieuses et allergiques.
Antigènes du corps humain. Du point de vue de la médecine clinique, le plus important est la détermination des antigènes spécifiques de groupe (antigènes du groupe sanguin), des antigènes spécifiques individuellement (antigènes d'histocompatibilité), spécifiques d'organes et de tissus (antigènes embryonnaires du cancer).
Antigènes d'histocompatibilité se trouvent sur les membranes cytoplasmiques de presque toutes les cellules du macro-organisme. La plupart d'entre eux sont liés au système complexe majeur d'histocompatibilité, ou WPC(de l'anglais. complexe principal d'hystocompatibilité). Chez l'homme, le CMH est désigné comme HLA(de l'anglais. antigène leucocytaire humain) puisqu'il est associé aux leucocytes. Les antigènes d'histocompatibilité jouent un rôle clé dans la reconnaissance spécifique de "l'ami ou l'ennemi" et l'induction de la réponse immunitaire acquise. Ils déterminent la compatibilité des organes et des tissus lors de la transplantation au sein d'une même espèce, la restriction génétique (restriction) de la réponse immunitaire et d'autres effets. Par nature chimique, les antigènes d'histocompatibilité sont des glycoprotéines solidement associées à la membrane cytoplasmique des cellules. Leurs fragments individuels ont une homologie structurelle avec les molécules d'immunoglobuline. Il existe deux classes principales de molécules du CMH. Il est classiquement admis que le CMH de classe I induit une réponse immunitaire à prédominance cellulaire, et le CMH de classe II ¾ humoral.
Antigènes bactériens :
flagelles, ou Antigènes H , localisé dans l'appareil locomoteur des bactéries - flagelles. Ce sont des épitopes de la flagelline, une protéine contractile. Lorsqu'elle est chauffée, la flagelline se dénature et l'antigène H perd sa spécificité. Le phénol n'agit pas sur cet antigène ;
somatique, ou O antigène , associée à la paroi cellulaire bactérienne. Il est à base de lipopolysaccharide (LPS). L'antigène O présente des propriétés thermostables - il n'est pas détruit par l'ébullition. Cependant, l'antigène somatique est soumis à l'action des aldéhydes (par exemple, le formol) et des alcools, qui perturbent sa structure. Si un animal est immunisé avec des bactéries vivantes qui ont des flagelles, alors des anticorps seront produits qui sont dirigés simultanément contre les antigènes O et H. L'introduction de la culture bouillie dans l'animal stimule la biosynthèse des anticorps dirigés contre l'antigène somatique. Une culture de bactéries traitées au phénol provoquera la formation d'anticorps dirigés contre les antigènes flagellaires ;
capsule, ou Antigènes K , situé à la surface de la paroi cellulaire. On les trouve dans les bactéries qui forment une capsule. En règle générale, les antigènes K sont constitués de polysaccharides acides (acides uroniques). Parallèlement, dans le bacille du charbon, cet antigène est construit à partir de chaînes polypeptidiques. Par sensibilité à la chaleur, on distingue trois types d'antigènes K: A, B et L. La stabilité thermique la plus élevée est caractéristique du type A, elle ne se dénature pas même avec une ébullition prolongée. Le type B résiste à un chauffage court (environ 1 heure) jusqu'à 60 °C. Le type L se dégrade rapidement à cette température. Par conséquent, une élimination partielle de l'antigène K est possible par ébullition prolongée de la culture bactérienne. À la surface de l'agent causal de la fièvre typhoïde et d'autres entérobactéries hautement virulentes, on peut trouver une variante spéciale de l'antigène capsulaire. Il a obtenu le nom antigène de virulence, ou Antigène Vi.
Les bactéries bactériennes ont également des propriétés antigéniques. toxines protéiques, enzymes et certaines autres protéines qui sont sécrétées par des bactéries dans l'environnement (par exemple la tuberculine). Le tétanos, la diphtérie et les toxines botuliques font partie des antigènes puissants à part entière, ils sont donc utilisés pour obtenir des anatoxines pour la vaccination humaine.
Antigènes viraux :
nucléaire(vache);
capside(coquille);
supercapside(surface) .
La composition antigénique du virion dépend de la structure de la particule virale. La spécificité antigénique des virus simplement organisés est associée aux ribo- et désoxyribonucléoprotéines. Ces substances sont très solubles dans l'eau et sont donc appelées antigènes S (lat. la solution- la solution). Dans les virus complexes, certains des antigènes sont associés à la nucléocapside et l'autre est localisé dans l'enveloppe externe (supercapside). Les antigènes de nombreux virus sont très variables. Cela est dû au processus mutationnel constant que subit l'appareil génétique de la particule virale. Un exemple est le virus de la grippe, les virus de l'immunodéficience humaine.