Tensiunea peretelui celular la plante. perete celular
Citeste si
În zorii dezvoltării vieții pe Pământ, toate formele celulare erau reprezentate de bacterii. Au aspirat materia organică dizolvată în oceanul primordial prin suprafața corpului.
De-a lungul timpului, unele bacterii s-au adaptat pentru a produce substanțe organice din cele anorganice. Pentru a face acest lucru, au folosit energia luminii solare. Primul sistem ecologicîn care aceste organisme au fost producători. Ca urmare, oxigenul eliberat de aceste organisme a apărut în atmosfera Pământului. Cu el, puteți obține mult mai multă energie din același aliment și puteți utiliza energia suplimentară pentru a complica structura corpului: împărțirea corpului în părți.
Unul dintre realizări importante viata - separarea nucleului si citoplasmei. Nucleul conține informații ereditare. O membrană specială în jurul miezului a făcut posibilă protejarea împotriva daunelor accidentale. După cum este necesar, citoplasma primește comenzi de la nucleu care direcționează activitatea vitală și dezvoltarea celulei.
Organismele în care nucleul este separat de citoplasmă au format super-regnul nuclearului (acestea includ plante, ciuperci, animale).
Astfel, celula - baza organizării plantelor și animalelor - a apărut și s-a dezvoltat în cursul evoluției biologice.
Chiar și cu ochiul liber, și chiar mai bine la lupă, puteți vedea că pulpa unui pepene copt este formată din boabe foarte mici, sau boabe. Acestea sunt celule – cele mai mici „cărămizi” care alcătuiesc corpurile tuturor organismelor vii, inclusiv ale plantelor.
Viața unei plante se desfășoară prin activitatea combinată a celulelor sale, creând un singur întreg. Odată cu multicelularitatea părților plantelor, are loc o diferențiere fiziologică a funcțiilor acestora, specializarea diferitelor celule în funcție de localizarea lor în corpul plantei.
O celulă vegetală diferă de o celulă animală prin faptul că are o înveliș dens care acoperă conținutul interior din toate părțile. Celula nu este plată (așa cum este de obicei descrisă), cel mai probabil arată ca o fiolă foarte mică umplută cu conținut viros.
Structura și funcțiile unei celule vegetale
Considerați o celulă ca o unitate structurală și funcțională a unui organism. În exterior, celula este acoperită cu un perete celular dens, în care există secțiuni mai subțiri - pori. Sub ea se află o peliculă foarte subțire - o membrană care acoperă conținutul celulei - citoplasma. În citoplasmă există cavități - vacuole umplute cu seva celulară. La sau aproape de centrul celulei perete celular este situat un corp dens - un nucleu cu un nucleol. Nucleul este separat de citoplasmă prin învelișul nuclear. Corpurile mici, plastidele, sunt distribuite în întreaga citoplasmă.
Structura celula plantei
Structura și funcțiile organelelor celulelor vegetale
| Organoid | Desen | Descriere | Funcţie | Particularități |
Peretele celular sau membrana plasmatica | Incolor, transparent și foarte durabil | Trece în celulă și eliberează substanțe din celulă. | Membrana celulară este semi-permeabilă |
|
Citoplasma | Substanță groasă, vâscoasă | Conține toate celelalte părți ale celulei. | Este în continuă mișcare |
|
Nucleu (parte importantă a celulei) | rotund sau oval | Asigură transferul proprietăților ereditare către celulele fiice în timpul diviziunii | Partea centrală a celulei |
|
Forma sferică sau neregulată | Ia parte la sinteza proteinelor | |||
 | Rezervor separat de citoplasmă printr-o membrană. Conține seva celulară | Se acumulează substanțe nutritive de rezervă și deșeuri care nu sunt necesare celulei. | Pe măsură ce celula crește, vacuolele mici se contopesc într-o singură vacuola mare (centrală). |
|
plastide | Cloroplaste | Utilizați energia luminoasă a soarelui și creați organic din anorganic | Forma discurilor separate de citoplasmă printr-o membrană dublă |
|
Cromoplastele | Formată ca urmare a acumulării de carotenoizi | Galben, portocaliu sau maro |
||
 | Leucoplaste | Plastide incolore | ||
plic nuclear | Constă din două membrane (exterioară și interioară) cu pori | Separă nucleul de citoplasmă | Permite schimbul între nucleu și citoplasmă |
Partea vie a celulei este un sistem limitat de membrană, ordonat, structurat de biopolimeri și structuri membranare interne implicate în totalitatea proceselor metabolice și energetice care mențin și reproduc întregul sistem în ansamblu.
O caracteristică importantă este că nu există membrane deschise cu capete libere în celulă. Membranele celulare limitează întotdeauna cavitățile sau zonele, închizându-le din toate părțile.

Diagrama generalizată modernă a unei celule vegetale
plasmalema(membrană celulară exterioară) - o peliculă ultramicroscopică de 7,5 nm grosime., Constând din proteine, fosfolipide și apă. Aceasta este o peliculă foarte elastică care este bine umezită de apă și restabilește rapid integritatea după deteriorare. Are o structură universală, adică tipică pentru toate membranele biologice. Celulele vegetale din afara membranei celulare au un perete celular puternic care creează un suport extern și menține forma celulei. Este alcătuit din fibre (celuloză), o polizaharidă insolubilă în apă.
Plasmodesmate a unei celule vegetale, sunt tubuli submicroscopici care pătrund prin membrane și căptușiți cu o membrană plasmatică, care trece astfel de la o celulă la alta fără întrerupere. Cu ajutorul lor, are loc circulația intercelulară a soluțiilor care conțin nutrienți organici. Ele transmit, de asemenea, biopotențiale și alte informații.
Poromy numite găuri în membrana secundară, unde celulele sunt separate doar de membrana primară și placa de mijloc. Zonele membranei primare și ale plăcii de mijloc care separă porii adiacenți ai celulelor adiacente se numesc membrana porilor sau filmul de închidere a porului. Pelicula de închidere a porului este străpunsă de tubuli plasmodesmenali, dar de obicei nu se formează un orificiu traversant în pori. Porii facilitează transportul apei și al substanțelor dizolvate de la celulă la celulă. În pereții celulelor învecinate, de regulă, unul împotriva celuilalt, se formează pori.
Perete celular are o învelișă bine definită, relativ groasă, de natură polizaharidă. Peretele celular al plantei este un produs al citoplasmei. Aparatul Golgi și reticulul endoplasmatic participă activ la formarea acestuia.

Structura membranei celulare
Baza citoplasmei este matricea sa, sau hialoplasma, un sistem coloidal complex incolor, transparent optic, capabil de tranziții reversibile de la sol la gel. Rolul cel mai important al hialoplasmei este de a uni toate structurile celulare în sistem unic si asigurarea interactiunii intre ele in procesele de metabolism celular.
Hialoplasma(sau matricea citoplasmatică) este mediu intern celule. Compus din apă și diverși biopolimeri (proteine, acizi nucleici, polizaharide, lipide), dintre care cea mai mare parte sunt proteine cu diverse specificități chimice și funcționale. Hialoplasma conține, de asemenea, aminoacizi, monozaharuri, nucleotide și alte substanțe cu greutate moleculară mică.
Biopolimerii formează un mediu coloidal cu apă, care, în funcție de condiții, poate fi dens (sub formă de gel) sau mai lichid (sub formă de sol), atât în întreaga citoplasmă, cât și în secțiunile sale individuale. În hialoplasmă, diverse organele și incluziuni sunt localizate și interacționează între ele și cu mediul hialoplasmei. Mai mult, locația lor este cel mai adesea specifică anumitor tipuri de celule. Prin membrana bilipidiană, hialoplasma interacționează cu mediul extracelular. Prin urmare, hialoplasma este un mediu dinamic și joacă rol importantîn funcționarea organelelor individuale și în activitatea vitală a celulelor în ansamblu.
Formațiuni citoplasmatice – organite
Organelele (organelele) sunt componentele structurale ale citoplasmei. Au o anumită formă și dimensiune, sunt structuri citoplasmatice obligatorii ale celulei. În absența sau deteriorarea lor, celula își pierde de obicei capacitatea de a continua să existe. Multe dintre organele sunt capabile de divizare și auto-reproducere. Sunt atât de mici încât pot fi văzute doar cu un microscop electronic.
Miez
Nucleul este cel mai vizibil și, de obicei, cel mai mare organel al celulei. A fost studiat pentru prima dată în detaliu de Robert Brown în 1831. Nucleul asigură cele mai importante funcții metabolice și genetice ale celulei. Are formă destul de variabilă: poate fi sferică, ovală, lobată, lenticulară.
Nucleul joacă un rol important în viața celulei. O celulă din care a fost îndepărtat nucleul nu mai secretă o înveliș, nu mai crește și nu mai sintetizează substanțe. Produsele degradării și distrugerii se intensifică în ea, în urma cărora moare rapid. Formarea unui nou nucleu din citoplasmă nu are loc. Nucleele noi se formează numai prin fisiunea sau zdrobirea celui vechi.
Conținutul intern al nucleului este cariolimfa (sucul nuclear), care umple spațiul dintre structurile nucleului. Conține unul sau mai mulți nucleoli, precum și un număr semnificativ de molecule de ADN conectate la anumite proteine - histone.

Structura nucleului
nucleol
Nucleolul, ca și citoplasma, conține în principal ARN și proteine specifice. Funcția sa cea mai importantă este aceea că are loc în ea formarea ribozomilor, care realizează sinteza proteinelor în celulă.
aparate Golgi
Aparatul Golgi este un organoid care are o distribuție universală în toate tipurile de celule eucariote. Este un sistem cu mai multe niveluri de saci cu membrană plate, care se îngroașă de-a lungul periferiei și formează procese veziculare. Cel mai adesea este situat în apropierea nucleului.

aparate Golgi
Aparatul Golgi include în mod necesar un sistem de vezicule mici (vezicule), care sunt împletite din cisterne (discuri) îngroșate și sunt situate de-a lungul periferiei acestei structuri. Aceste vezicule joacă rolul de intracelular sistem de transport granulele specifice sectorului pot servi ca sursă de lizozomi celulari.
Funcțiile aparatului Golgi constau și în acumularea, separarea și eliberarea produselor de sinteză intracelulară, a produselor de descompunere și a substanțelor toxice în afara celulei cu ajutorul bulelor. Produsele activității sintetice a celulei, precum și diferite substanțe care intră în celulă din mediu prin canalele reticulului endoplasmatic, sunt transportate în aparatul Golgi, se acumulează în acest organoid și apoi intră în citoplasmă sub formă de picături sau boabe și sunt fie utilizate de celula însăși, fie excretate. În celulele vegetale, aparatul Golgi conține enzime pentru sinteza polizaharidelor și materialul polizaharidic în sine, care este folosit pentru a construi peretele celular. Se crede că este implicat în formarea vacuolelor. Aparatul Golgi a fost numit după omul de știință italian Camillo Golgi, care l-a descoperit pentru prima dată în 1897.
Lizozomi
Lizozomii sunt vezicule mici, limitate de o membrană, a căror funcție principală este realizarea digestiei intracelulare. Utilizarea aparatului lizozomal are loc în timpul germinării semințelor plantei (hidroliza de rezervă nutrienți).
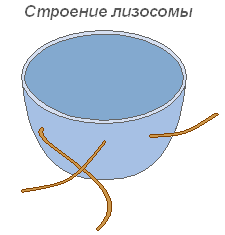
Structura lizozomului
microtubuli
Microtubulii sunt structuri supramoleculare membranare formate din globule proteice dispuse in siruri spiralate sau drepte. Microtubulii îndeplinesc o funcție predominant mecanică (motorie), oferind mobilitate și contractilitate organelelor celulare. Situate în citoplasmă, ele dau celulei o anumită formă și asigură stabilitatea aranjamentului spațial al organelelor. Microtubulii facilitează mișcarea organitelor către locații determinate de nevoi fiziologice celule. Un număr semnificativ dintre aceste structuri sunt situate în plasmalemă, în apropierea membranei celulare, unde sunt implicate în formarea și orientarea microfibrilelor de celuloză ale membranelor celulare vegetale.

Structura microtubulilor
Vacuole
Vacuola este cea mai importantă componentă a celulelor vegetale. Este un fel de cavitate (rezervor) în masa citoplasmei, umplută cu o soluție apoasă saruri minerale, aminoacizi, acizi organici, pigmenți, carbohidrați și separat de citoplasmă printr-o membrană vacuolară - tonoplastul.
Citoplasma umple întreaga cavitate internă numai în cele mai tinere celule vegetale. Odată cu creșterea celulei, aranjarea spațială a masei inițial continue a citoplasmei se modifică semnificativ: în ea apar mici vacuole umplute cu seva celulară, iar întreaga masă devine spongioasă. Odată cu creșterea ulterioară a celulelor, vacuolele individuale se îmbină, împingând straturile citoplasmatice la periferie, ca urmare a faptului că există de obicei o vacuola mare în celula formată, iar citoplasma cu toate organitele sunt situate în apropierea membranei.
Compușii organici și minerali solubili în apă ai vacuolelor determină proprietățile osmotice corespunzătoare ale celulelor vii. Această soluție de o anumită concentrație este un fel de pompă osmotică pentru pătrunderea controlată în celulă și eliberarea de apă, ioni și molecule de metaboliți din aceasta.
În combinație cu stratul de citoplasmă și membranele sale, care se caracterizează prin proprietăți de semipermeabilitate, vacuola formează un sistem osmotic eficient. Determinați osmotic sunt indicatori ai celulelor vii ale plantelor, cum ar fi potențialul osmotic, forța de aspirație și presiunea turgenței.

Structura vacuolei
plastide
Plastidele sunt cele mai mari (după nucleu) organele citoplasmatice, inerente doar celulelor vegetale. Nu se găsesc numai în ciuperci. Plastidele joacă un rol important în metabolism. Ele sunt separate de citoplasmă printr-o membrană dublă cu membrană, iar unele dintre tipurile lor au un sistem bine dezvoltat și ordonat de membrane interne. Toate plastidele sunt de aceeași origine.
Cloroplaste- cele mai comune și mai importante plastide funcțional ale organismelor fotoautotrofe care realizează procese fotosintetice care duc în cele din urmă la formarea materie organicăși eliberarea de oxigen liber. Cloroplastele plantelor superioare au un complex structura interna.

Structura cloroplastei
Dimensiunile cloroplastelor din diferite plante nu sunt aceleași, dar în medie diametrul lor este de 4-6 microni. Cloroplastele sunt capabile să se deplaseze sub influența mișcării citoplasmei. În plus, sub influența iluminării, se observă o mișcare activă a cloroplastelor de tip ameboid către sursa de lumină.
Clorofila este substanța principală a cloroplastelor. Datorită clorofilei plante verzi capabil să utilizeze energia luminoasă.
Leucoplaste(plastide incolore) sunt corpuri clar marcate ale citoplasmei. Dimensiunile lor sunt oarecum mai mici decât dimensiunile cloroplastelor. Mai uniforme și forma lor, apropiindu-se de sferic.

Structura leucoplastei
Se găsesc în celulele epidermei, tuberculi, rizomi. Când sunt iluminate, se transformă foarte repede în cloroplaste, cu o modificare corespunzătoare a structurii interne. Leucoplastele conțin enzime, cu ajutorul cărora amidonul este sintetizat din excesul de glucoză format în timpul fotosintezei, cea mai mare parte a cărora se depune în țesuturile sau organele de depozitare (tuberculi, rizomi, semințe) sub formă de boabe de amidon. La unele plante, grăsimile se depun în leucoplaste. Funcția de rezervă a leucoplastelor se manifestă ocazional prin formarea de proteine de depozitare sub formă de cristale sau incluziuni amorfe.
Cromoplasteleîn majoritatea cazurilor sunt derivați ai cloroplastelor, ocazional - leucoplaste.

Structura cromoplastei
Maturarea maceselor, ardeilor, rosiilor este insotita de transformarea cloro- sau leucoplastelor celulelor pulpei in carotenoizi. Acestea din urmă conțin preponderent pigmenți plastidici galbeni - carotenoizi, care, la maturare, se sintetizează intens în ei, formând picături lipidice colorate, globule solide sau cristale. Clorofila este distrusă.
Mitocondriile
Mitocondriile sunt organite care se găsesc în majoritatea celulelor vegetale. Au o formă variabilă de bețe, boabe, fire. Au fost descoperite în 1894 de R. Altman folosind un microscop cu lumină, iar structura internă a fost studiată ulterior folosind unul electronic.
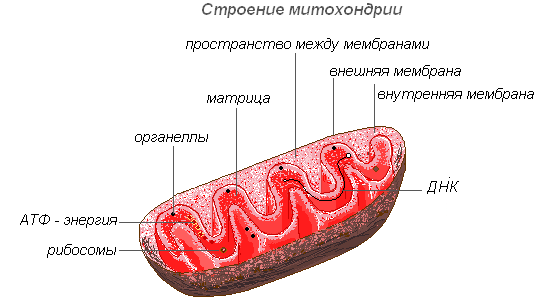
Structura mitocondriilor
Mitocondriile au o structură cu două membrane. Membrana exterioară este netedă, se formează cea interioară diverse forme excrescențe - tubuli din celulele plantelor. Spațiul din interiorul mitocondriilor este umplut cu conținut de semi-lichid (matrice), care include enzime, proteine, lipide, săruri de calciu și magneziu, vitamine, precum și ARN, ADN și ribozomi. Complexul enzimatic al mitocondriilor accelerează activitatea unui mecanism complex și interconectat al reacțiilor biochimice, în urma căruia se formează ATP. În aceste organite, celulele sunt furnizate cu energie - energia legăturilor chimice ale nutrienților este transformată în legături de mare energie ale ATP în procesul de respirație celulară. În mitocondrii are loc descompunerea enzimatică a carbohidraților, acizilor grași și aminoacizilor odată cu eliberarea de energie și conversia ulterioară a acesteia în energie ATP. Energia acumulată este cheltuită pe procese de creștere, pe noi sinteze etc. Mitocondriile se reproduc prin diviziune și trăiesc aproximativ 10 zile, după care sunt distruse.
Reticulul endoplasmatic
Reticulul endoplasmatic - o rețea de canale, tubuli, vezicule, cisterne situate în interiorul citoplasmei. Deschis în 1945 de omul de știință englez K. Porter, este un sistem de membrane cu structură ultramicroscopică.

Structura reticulului endoplasmatic
Întreaga rețea este integrată într-un singur întreg cu un extern membrana celulara carcasă nucleară. Distingeți ER neted și aspru, purtător de ribozomi. Pe membranele EPS netede există sisteme enzimatice implicate în metabolismul grăsimilor și carbohidraților. Acest tip de membrană predomină în celulele semințelor bogate în substanțe de rezervă (proteine, carbohidrați, uleiuri), ribozomii sunt atașați de membrana ER granulară, iar în timpul sintezei unei molecule proteice, lanțul polipeptidic cu ribozomi este scufundat în ER. canal. Funcțiile reticulului endoplasmatic sunt foarte diverse: transportul de substanțe atât în interiorul celulei, cât și între celulele învecinate; divizarea unei celule în secțiuni separate în care au loc simultan diferite procese fiziologice și reacții chimice.
Ribozomi
Ribozomii sunt organite celulare non-membranare. Fiecare ribozom este format din două particule de dimensiuni inegale și poate fi împărțit în două fragmente care continuă să păstreze capacitatea de a sintetiza proteine după combinarea într-un ribozom întreg.
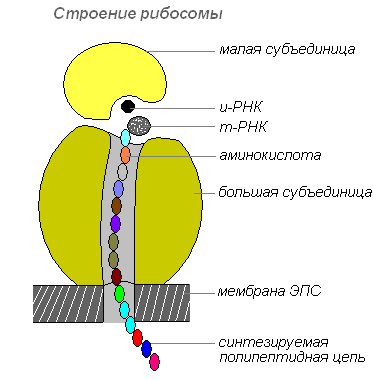
Structura ribozomului
Ribozomii sunt sintetizați în nucleu, apoi îl părăsesc, trecând în citoplasmă, unde sunt atașați de suprafața exterioară a membranelor reticulului endoplasmatic sau sunt localizați liber. În funcție de tipul de proteină sintetizată, ribozomii pot funcționa singuri sau se pot combina în complexe - poliribozomi.
Peretele celular este un derivat al protoplastei, adică. se formează în cursul activităţii sale vitale (fig. 61). Oferă celulei o anumită formă, protejează protoplastul și, rezistând presiunii intracelulare, previne ruperea celulei. Îndeplinesc funcțiile scheletului intern al unei plante, pereții celulari conferă organelor acesteia rezistența mecanică necesară.
Pereții celulari transmit bine lumina soarelui, apa și mineralele dizolvate în ea se deplasează cu ușurință de-a lungul lor. Între pereții celulelor adiacente se află placa de mijloc - stratul de pectină, care, fiind, de fapt, o substanță intercelulară, ține împreună pereții celulelor învecinate. În acele locuri în care pereții celulari ai celulelor vecine nu se închid, se formează celule pline cu apă. spatii intercelulare. Procesul de distrugere a substanței intercelulare, în urma căruia pereții celulelor învecinate sunt separați, se numește macerare. Se poate observa macerare naturală
Orez. 61.
A- schema structurii peretelui celular; B- schema de participare a aparatului Golgi la construirea peretelui celular; ÎN- structura detaliată a peretelui celular: 1 - placa de mijloc 2 - este timpul; 3 - perete secundar;
- 4 - perete primar; 5 - dictiozom; 6 - vezicule Golgi;
- 7- plasmalema; 8- perete celular; 9- macrofibrilă;
- 10 - microfibrila; 11 - micela; 12 - moleculă de celuloză;
- 13 - structura unui fragment dintr-o moleculă de celuloză
în fructele supracoapte de măr, frasin de munte, pepene galben etc. Macerația artificială se efectuează, de exemplu, la înmuierea tulpinilor de in pentru a separa fibrele de liban de ele; apare și în timpul tratamentului termic al fructelor.
Peretele celular conține polizaharide: pectine, hemiceluloză și celuloză. Moleculele de celuloză foarte lungi sunt ordonate paralel între ele (40-60), formându-se micelii. Micelele sunt colectate în ciorchini - microfibrile, care sunt unitatea structurală de bază a celulozei. Microfibrilele, la rândul lor, sunt combinate în macrofibrile- fibre foarte subtiri de lungime nedeterminata. Macrofibrilele de celuloză sunt scufundate într-un mediu foarte udat matrice, constând din pectine, hemiceluloze și alte substanțe. Rezistența peretelui celular este dată de microfibrile de celuloză elastică, care sunt apropiate de oțel ca rezistență la tracțiune. Forța și elasticitatea peretelui celular stau la baza capacității sale de a se întinde reversibil. Datorită pectinelor și hemicelulozei, peretele celular este foarte permeabil la apă - apa și substanțele dizolvate în ea se deplasează cu ușurință de-a lungul ei de la celulă la celulă.
Peretele celular se învecinează la exterior cu plasmalema, care este implicată activ în creșterea acesteia. Moleculele de pectine, hemiceluloză, celuloză și alte substanțe sunt sintetizate și acumulate în cisternele dictiozomale ale aparatului Golgi. Veziculele Golgi le livrează la periferia protoplastei - la plasmalemă. În punctul de contact dintre veziculă și plasmalemă, aceasta din urmă se dizolvă, iar conținutul veziculei, ajungând în afara plasmalemei, merge pentru a construi peretele celular. Membrana cu bule nu numai că restabilește integritatea plasmalemei, dar asigură și creșterea superficială a acesteia. Creșterea peretelui celular se realizează datorită activității enzimatice a plasmalemei.
Se numesc pereții celulelor în diviziune și creștere primar. Conțin multă apă (60-90%), pectinele și hemiceluloza predomină în substanța uscată - celuloza în ea nu este mai mare de 30%. Când o celulă se împarte în telofaza mitozei, celula mamă este împărțită în două celule fiice ca urmare a formării unui sept în planul său ecuatorial - placa de mijloc. Pe ambele părți ale plăcii de mijloc, fiecare dintre cele două celule fiice începe să-și creeze peretele celular primar. Creșterea laminei mediane și a pereților primari ai două celule fiice se desfășoară într-o direcție centrifugă - de la centrul celulei mamă la periferia acesteia. Lamina mediană este foarte subțire și este formată din pectină.
Noua celulă formată ca urmare a diviziunii începe să crească, în timp ce volumul ei poate crește de 100 sau de mai multe ori. Creșterea celulară are loc în principal prin entorse datorita absorbtiei de apa si cresterii volumului vacuolelor. Presiunea internă rezultată întinde peretele primar, în care pătrund ușor micelele de celuloză, pectinele și hemiceluloza. Metoda de creștere a peretelui celular prin implementare material de construcție între structurile existente se numește invaginatie.
În peretele celular primar, inițial există secțiuni mai subțiri în care fibrilele de celuloză sunt situate mai liber, - câmpurile porilor primari. Câmpurile de pori primare ale pereților a două celule învecinate coincid de obicei. Aici, tubulii reticulului endoplasmatic trec de la o celulă la alta - plasmodesmate. Căile pe care plasmodesmele le iau de la o celulă la alta sunt numite tubii plasmodesmenali. Prin acești tubuli, hialoplasmele celulelor învecinate sunt conectate între ele. Prin plasmodesmate se realizează transportul intercelular al substanțelor (hormoni, aminoacizi, ATP, zaharuri etc.). Protoplastele celulelor corpului unite într-un singur întreg cu ajutorul plasmodesmelor se numesc simplast. Transportul substanțelor de-a lungul plasmodesmelor se numește simplistic.(Combinația dintre pereții celulari, lamina medie și spațiile intercelulare se numește apoplast,îi urmărește apoplastic transport de substanțe.)
După terminarea creșterii celulare, peretele său poate rămâne primar subțire (în celulele țesuturilor educaționale) sau poate începe să crească în grosime (în celulele țesuturilor permanente). Creșterea peretelui celular în grosime se numește îngroșare secundară. Ca urmare, se depune pe suprafața interioară a peretelui primar perete secundar, care crește cu apozitii- impunerea micelilor de celuloză pe peretele deja existent. În acest caz, cele mai tinere straturi ale peretelui celular secundar sunt situate lângă plasmalemă. Peretele celular secundar îndeplinește în principal funcții mecanice de susținere. Conține în mod semnificativ mai putina apa decât în primară, iar celuloza predomină în substanța uscată (până la 50%). De exemplu, în pereții secundari ai firelor de păr de bumbac unicelulare și a fibrelor de liban de in, conținutul de celuloză poate ajunge la 95%.
Îngroșarea secundară a peretelui celular are loc neuniform. Zonele peretelui celular secundar din locațiile câmpurilor primare ale porilor rămân de obicei neîngroșate. Astfel de zone neîngroșate ale peretelui celular sunt numite porii. Porii din pereții a două celule adiacente, de regulă, coincid, formându-se de cateva ori. Canalul de pori format dintr-o pereche de pori este blocat pelicula de închidere a porilor - un sept format dintr-o lamină mediană și doi pereți primari ai celulelor adiacente. Pelicula de închidere a porului este străpunsă de numeroși tubuli plasmodesmenali prin care trec plasmodesmele.
Distinge porii simpluȘi franjuri(Fig. 62). Pentru porii simpli, diametrul secțiunii lor a canalului porilor este același pe toată lungimea, adică. este de formă cilindrică. Porii simpli sunt tipici celulelor parenchimatoase. Porii mărginiți sunt caracteristici pereților celulelor care conduc apa cu minerale dizolvate - traheide și segmente vasculare. În astfel de pori, secțiunea lor a canalului porilor are forma unei pâlnii, care, cu latura sa largă, este adiacentă filmului de închidere a porului.
În celulele țesuturilor conductoare de conifere, pelicula de închidere a porului este permeabilă la apă numai la margini, deoarece partea centrală îngroșată și lignificată, asemănătoare unui disc, este torus - nu trece apa. Torul acționează ca o supapă. Dacă presiunea apei în celulele vecine nu este aceeași, pelicula de închidere este deviată și torul blochează mișcarea apei prin canalul porilor.

Orez. 62.
A- simplu; B- marginita; ÎN- semi-marginit:
1 - folie de inchidere; 2 - canalul porilor; 3 - torus
În pereții celulelor conductoare de apă, pe lângă pori, perforatii- prin orificii (segmente de vase de sânge, celule de stocare a apei din mușchi de sphagnum).
Modificări ale peretelui celular. În funcție de funcțiile îndeplinite de celulă, peretele acesteia poate fi modificat datorită depunerii în ea a oricăror substanțe. Modificările sale uzuale sunt: lignificarea, plutarea, cutinizarea, mineralizarea și slimarea.
Lignificarea peretelui celular, sau lignificare, apare ca urmare a depunerii de lignină în spațiile intermicelare - o substanță de natură aromatică cu un complex structura chimica. Rezistența și duritatea peretelui cresc în același timp, dar elasticitatea acestuia scade. Pereții lemnos sunt capabili să treacă apa și aerul. Cu un perete celular lignificat, protoplastul celulei poate rămâne în viață, dar de obicei moare. La unele plante lemnoase, până la 30% din lignină se acumulează în lemn. Lignina se poate acumula și în pereții celulari ai lăstarilor de iarbă îmbătrâniți, ceea ce reduce semnificativ valoarea lor nutritivă și determină momentul recoltării fânului. În procesul de obținere a celulozei din lemn, care este necesară pentru producția de hârtie, se efectuează despădurirea artificială. Lignificarea naturală a peretelui celular este posibilă, dar rară.
dop, sau suberinizare, - depunerea în peretele celular a unei substanțe amorfe persistente asemănătoare grăsimii suberina (polimer hidrofob). Pereții celulelor corky sunt impermeabili la gaze și apă, ceea ce provoacă moartea protoplastei. Celulele cu pereți înfundați protejează în mod fiabil plantele de pierderea de apă, temperaturi extreme, bacterii patogene și ciuperci.
Cutinizarea - depunerea în pereții celulari a cutinei (o substanță similară cu compoziție chimică la suberina). Cutina este de obicei depusă în straturile superficiale ale pereților exteriori ai celulelor și pe suprafața acestora. Sub forma unui film - o cuticulă - acoperă, de exemplu, suprafața celulelor țesutului tegumentar - epiderma.
Mineralizare peretele celular se datorează depunerii de săruri de calciu și silice în acesta. Aceste substanțe dau peretelui duritate și fragilitate. Procesul de mineralizare este deosebit de bine exprimat în pereții celulari ai epidermei lăstarilor de cereale, rogoz și coada-calului. Din acest motiv, se recomandă cosirea lăstarilor de rogoz și ierburi înainte ca aceștia să înflorească - ulterior, datorită mineralizării puternice, se îngroșează, ceea ce scade calitatea fânului.
slime- transformarea celulozei și a pectinelor peretelui celular în polizaharide speciale - mucus și gingii, capabile să se umfle puternic la contactul cu apa. Mucilagiul peretelui este observat în celulele învelișului semințelor, de exemplu, la gutui, in, castraveți și pătlagină. Mucusul lipicios poate ajuta la dispersarea semintelor (psyllium); în timpul germinării semințelor, mucusul, absorbind și reținând apa, le protejează de uscare. În capacul rădăcinii, mucusul joacă rolul unui lubrifiant, facilitând trecerea rădăcinii între bulgări de sol. Mucusul și gingiile se pot forma în cantități semnificative atunci când pereții celulari sunt dizolvați din cauza deteriorării lor. La cireșe și prune, secreția de gumă este adesea observată atunci când ramurile și trunchiurile sunt rănite. Așa-numitul clei de cireșe este o gumă care se întărește sub formă de influxuri, care acoperă suprafața rănilor, găurile de îngheț, împiedicând pătrunderea infecției în ele. Mucusul de această natură se numește humozași considerat un fenomen patologic.
Deoarece pereții celulari secundari joacă rolul scheletului intern al plantei, dând puterea necesară organelor acesteia (ceea ce este deosebit de important pentru plante terestre), adesea sunt capabili să se îngroașă semnificativ - local sau complet - pentru a da o rezistență mai mare țesutului și, prin urmare, organului plantei. Îngroșarea peretelui celular are loc din cauza depunerii de celuloză.
Funcțiile celulelor sunt adesea îndeplinite exclusiv de pereții lor, deoarece protoplastele celulare mor. Acest lucru se aplică celulelor de plută,

Orez. 63.
traheide, segmente vasculare, fibre tisulare mecanice. Lemnul, care ocupă majoritatea trunchiurilor uriașe de copaci, constă, de exemplu, în principal din pereți celulari lignificati, ale căror protoplaste au murit de mult.
Pereții celulari joacă un rol important în viața noastră. Din acestea se obțin materii prime textile (păr din semințe de bumbac, fibre de in etc.) și materii prime pentru funii și funii (fibre de cânepă, sfoară, sisal etc.). Celuloza extrasă din pereții celulelor este folosită la fabricarea hârtiei (molid, lemn de aspen), acetat de mătase, viscoză, materiale plastice, celofan și multe altele. O țesătură constând din celule moarte cu pereți cu plută - pluta a fost folosită de mult timp ca material valoros de izolare termică etanșă la apă și la aer și este din ce în ce mai folosită în construcțiile moderne.
Peretele celular al plantelor este format cu participarea membranei plasmatice și este o formațiune multistrat extracelulară (extracelulară) care protejează suprafața celulei și servește ca schelet exterior al celulei plantei (Fig. 1). Peretele celular al plantelor este format din două componente: o matrice (bază) de plastic amorf, asemănător unui gel, cu un conținut ridicat de apă și un sistem fibrilar de susținere. Substanțele și sărurile polimerice suplimentare, adesea incluse în compoziția cochiliilor, le conferă rigiditate și le fac neumezibile.
Orez. 1. Schema structurii peretelui celular al plantelor: 0 - placa de mijloc; 1 -- înveliș primar (două straturi pe fiecare parte a lui 0); 2 -- straturi ale cochiliei secundare; 3 -- înveliș terțiar; PM - membrana plasmatica; B - vacuol; Eu sunt miezul
Din punct de vedere chimic, principalele componente ale membranelor plantelor sunt polizaharidele structurale. Compoziția matricei membranelor plantelor include grupe eterogene de polizaharide care se dizolvă în alcalii concentrate, hemiceluloze și substanțe pectinice. Hemicelulozele sunt lanțuri polimerice ramificate formate din diverse hexoze (glucoză, manoză, galactoză etc.), pentoze (xiloză, arabinoză) și acizi uronici (glucuronic și galacturonic). Aceste componente ale hemicelulozelor sunt combinate între ele în diferite rapoarte cantitative și formează diverse combinații. Lanțurile de molecule de hemiceluloză nu cristalizează și nu formează fibrile elementare. Datorită prezenței grupelor polare de acizi uronici, aceștia sunt foarte hidratați.
Substanțele pectice sunt un grup eterogen care include polimeri ramificați, foarte hidratați, care poartă sarcini negative datorită numeroaselor reziduuri de acid galacturonic. Datorită proprietăților componentelor sale, matricea este o masă de plastic moale întărită cu fibrile.
Componentele fibroase ale pereților celulelor vegetale constau de obicei din celuloză, un polimer liniar, neramificat al glucozei. Greutatea moleculară a celulozei variază de la 5.104 la 5.105, ceea ce corespunde la 300-3000 de resturi de glucoză. Astfel de molecule liniare de celuloză pot fi combinate în mănunchiuri sau fibre. În peretele celular, celuloza formează fibrile, care constau din microfibrile submicroscopice de până la 25 nm grosime, care la rândul lor constau din multe lanțuri paralele de molecule de celuloză.
Raporturile cantitative dintre celuloză și substanțele matrice (hemiceluloză) pot fi foarte diferite pentru diferite obiecte. Peste 60% din masa uscată a membranelor primare este matricea acestora și aproximativ 30% este substanța scheletică - celuloza. În membranele celulare brute, aproape toată apa este asociată cu hemiceluloze; prin urmare, masa substanței principale în starea umflată atinge 80% din masa umedă a întregii membrane, în timp ce conținutul de substanțe fibroase se reduce la doar 12%. La firele de păr din bumbac, componenta celuloză este de 90%; în lemn, celuloza reprezintă 50% din componentele peretelui celular.
Pe lângă celuloză, hemiceluloză și pectine, membranele celulare includ componente suplimentare conferindu-le proprietati deosebite. De exemplu, incrustația (includerea în interior) a cochiliilor cu lignină (un polimer al alcoolului coniferilic) duce la lignificarea pereților celulari și la creșterea rezistenței acestora (Fig. 2). Lignina amestecă substanțele plastice ale matricei în astfel de învelișuri și joacă rolul substanței principale cu rezistență ridicată. Matricea este adesea întărită cu substanțe minerale (SiO2, CaCO3 etc.).

Orez. 2. Incrustație peretelui celular: a - schelă fibrilară și matrice interfibrilară; b - s-a încrustat cu lignină și și-a pierdut capacitatea de a întinde coaja cu resturile matricei; c - încrustarea ulterioară cu fenoli și (sau) minerale, ceea ce duce la creșterea durității învelișului
Pe suprafața membranei celulare se pot acumula diverse substanțe adcrustante, cum ar fi cutina și suberina, ceea ce duce la suberizarea celulelor. În celulele epidermei, pe suprafața membranelor celulare se depune ceara, care formează un strat impermeabil care împiedică celulele să piardă apă.
Datorită structurii sale poroase și libere, peretele celular al plantei este în mare măsură permeabil la compuși cu greutate moleculară mică, cum ar fi apa, zaharurile și ionii. Dar macromoleculele nu pătrund bine prin învelișurile de celuloză: dimensiunea porilor din învelișuri, care permite difuzarea liberă a substanțelor, este de numai 3-5 nm.
Experimentele cu compuși marcați au arătat că în timpul creșterii membranei celulare, eliberarea substanțelor din care este construită are loc pe întreaga suprafață a celulei. Substanțele amorfe ale matricei, hemicelulozele și pectinele sunt sintetizate în vacuolele aparatului Golgi și sunt eliberate prin plasmalemă prin exocitoză. Fibrile de celuloză sunt sintetizate de enzime speciale încorporate în plasmalemă.
Membranele celulelor diferențiate, mature sunt de obicei multistratificate, fibrilele de celuloză din straturi sunt orientate diferit, iar numărul lor poate varia, de asemenea, semnificativ. Pereții celulari primari, secundari și terțiari sunt de obicei descriși (vezi Fig. 1). Pentru a înțelege structura și aspectul acestor membrane, este necesar să vă familiarizați cu modul în care se formează după diviziunea celulară.
În timpul diviziunii celulelor vegetale, după divergența cromozomilor în planul ecuatorial al celulelor, apare o acumulare de vezicule membranare mici, care în partea centrală a celulelor încep să se contopească între ele (Fig. 3). Acest proces de fuziune a vacuolelor mici are loc din centrul celulei spre periferie și continuă până când veziculele membranare se contopesc între ele și cu membrana plasmatică a suprafeței laterale a celulei. Aceasta formează o placă celulară, sau phragmoplast. În partea sa centrală există o substanță amorfă a matricei, care a umplut bulele de fuziune. S-a dovedit că aceste vacuole primare provin din membranele aparatului Golgi. Compoziția peretelui celular primar include și o cantitate mică de proteină (aproximativ 10%) bogată în hidroxiprolină și având multe lanțuri scurte de oligozaharide, ceea ce determină această proteină ca glicoproteină. De-a lungul periferiei plăcii celulare, atunci când este observată în lumină polarizată, se detectează o birefringență vizibilă, cauzată de faptul că în acest loc sunt localizate fibrile de celuloză orientate. Astfel, peretele celular primar în creștere este format deja din trei straturi: cel central - placa de mijloc, constând doar dintr-o matrice amorfă, și două periferice - membrana primară care conține hemiceluloză și fibrile de celuloză. Dacă placa de mijloc este un produs al activității celulei originale, atunci membrana primară se formează datorită eliberării de fibrile de hemiceluloză și celuloză de către două noi corpuri celulare. Și toată creșterea în continuare a grosimii peretelui celular (sau mai bine zis, intercelular) va avea loc datorită activității a două celule fiice, care secretă substanțe ale membranei celulare din părți opuse, îngroșându-se prin stratificarea din ce în ce mai multe straturi noi. Încă de la început, eliberarea substanțelor matricei se realizează datorită apropierii veziculelor aparatului Golgi de membrana plasmatică, fuziunii lor cu membrana și eliberării conținutului lor în afara citoplasmei. Aici, în afara celulei, pe membrana plasmatică a acesteia, are loc sinteza și polimerizarea fibrilelor de celuloză. Așa se formează treptat membrana celulară secundară. Este dificil să se determine și să poată distinge învelișul primar de cel secundar cu suficientă precizie, deoarece acestea sunt interconectate prin mai multe straturi intermediare.
Masa principală a peretelui celular care și-a încheiat formarea este membrana secundară. Își dă cușca forma finala. După ce celula este împărțită în două celule fiice, celulele noi cresc, volumul lor crește și forma lor se schimbă; celulele sunt adesea alungite. În același timp, are loc o creștere a grosimii membranei celulare și o restructurare a structurii sale interne.
În timpul formării peretelui celular primar, există încă puține fibrile de celuloză în compoziția sa și sunt situate mai mult sau mai puțin perpendicular pe viitoarea axă longitudinală a celulei. Mai târziu, în timpul perioadei de alungire (alungirea celulei datorită creșterii vacuolelor în citoplasmă), orientarea acestor fibrile direcționate transversal suferă modificări pasive: fibrilele încep să fie situate în unghi drept unele față de altele și în cele din urmă devin alungite. mai mult sau mai puţin paralel cu axa longitudinală a celulei. Procesul se desfășoară constant: în straturile vechi (mai aproape de centrul cochiliei), fibrilele suferă deplasări pasive, iar depunerea de noi fibrile în straturile interioare (cel mai apropiat de membrana celulară) continuă în conformitate cu originalul. plan de construcție a carcasei. Acest proces creează posibilitatea ca fibrilele să alunece unele față de altele, iar rearanjarea armăturii membranei celulare este posibilă datorită stării gelatinoase a componentelor matricei sale. Ulterior, atunci când hemiceluloza este înlocuită cu lignină în matrice, mobilitatea fibrilelor scade brusc, coaja devine densă și are loc lignificarea.
Orez. 3. Schema de creștere a membranei celulare de la depunerea acesteia în timpul diviziunii celulare (I) până la maturarea completă (V): 1 - membrană primară; 2 -- straturi ale cochiliei secundare; 3 -- înveliș terțiar; B - vacuole; SP - placa de mijloc; PM - membranele plasmatice a două celule învecinate
Adesea, sub membrana secundară, se găsește o membrană terțiară, care poate fi considerată ca o rămășiță uscată a stratului degenerat al citoplasmei propriu-zise.
Trebuie remarcat faptul că, în diviziunea celulară a plantelor, formarea membranei primare nu este în toate cazurile precedată de formarea unei plăci celulare. Astfel, la algele verzi Spirogyra apar noi septuri transversale prin formarea de proeminențe pe pereții laterali ai celulei originale, care, crescând treptat spre centrul celulei, închid și împart celula în două.
După cum sa menționat deja, dacă o celulă este lipsită de membrana sa într-un mediu apos hipotonic, atunci va avea loc liza, ruptura celulei. S-a dovedit că prin selectarea concentrațiilor adecvate de săruri și zaharuri, este posibilă egalizarea presiunii osmotice în exteriorul și în interiorul celulelor, lipsite de membranele acestora. În același timp, astfel de protoplaste dobândesc formă sferică(sferoplaste). Dacă există o cantitate suficientă de nutrienți și săruri în mediul în care se află protoplastele (printre acestea este nevoie de Ca2+), atunci celulele sunt restaurate din nou, regenerează membrana celulară. Mai mult, în prezența hormonilor (auxine) aceștia sunt capabili să se dividă și să creeze colonii celulare, care pot da naștere la creșterea întregii plante din care a fost prelevată celula.
Principala componentă fibroasă a peretelui celular al grupurilor mari de ciuperci (bazibiomicete, ascomicete, zigomicete) este chitina; este o polizaharidă în care zaharida principală este N-acetilglucozamina. Compoziția peretelui celular al ciupercilor, pe lângă chitină, poate include substanțe ale matricei, glicoproteine și diverse proteine sintetizat în citoplasmă şi eliberat de celulă în exterior.
O coajă nouă format în procesul de diviziune celulară în stadiul final al mitozei - telofaza. După divergența cromozomilor în planul ecuatorial al celulelor, apare o acumulare de vezicule membranare mici, care în partea centrală a celulelor încep să fuzioneze unele cu altele. Acest proces de fuziune a vacuolelor mici are loc din centrul celulei spre periferie și continuă până când veziculele membranare se contopesc între ele și cu membrana plasmatică a suprafeței laterale a celulei. Așa se formează placă celulară sau fragmoplast.
ÎN Partea centrală conţine o substanţă amorfă a matricei, care a umplut veziculele care se fuzionează. S-a dovedit că aceste vacuole primare provin din membranele aparatului Golgi. Compoziția peretelui celular primar include și o cantitate mică de proteină (aproximativ 10%) bogată în hidroxiprolină și având multe lanțuri scurte de oligozaharide, ceea ce determină această proteină ca glicoproteină.
După formarea laminei mediane, protoplastul celulelor învecinate depune membrana primară pe ea. Stratul de celuloză care se depune în timpul creșterii celulare se numește peretele celular primar.
Pe lângă celuloză, hemiceluloză și pectină, învelișurile primare mai conțin proteină structurală – glicoproteină. Cojile primare se pot lignifica și ele, deși, de regulă, lignina nu este caracteristică acestora. Cu toate acestea, partea cea mai caracteristică a învelișului primar este componentă pectină. Oferă plasticitate cochiliei, îi permite să se întindă, pe măsură ce organele se alungesc: rădăcină, tulpină, frunză. Substanțele pectinice se pot umfla puternic, astfel încât învelișurile primare conțin multă apă (60-90%). Ponderea hemicelulozelor și a substanțelor pectinice reprezintă 50-60% din greutatea uscată a învelișului primar, conținutul de celuloză nu depășește 30%, proteina structurală ia până la 10%.
Procesul în curs de eliberare a substanțelor matricei se realizează datorită apropierii veziculelor aparatului Golgi de membrana plasmatică, fuziunii lor cu membrana și eliberării conținutului lor în afara citoplasmei. Aici, în afara celulei, pe membrana plasmatică a acesteia, are loc sinteza și polimerizarea fibrilelor de celuloză. Așa se dezvoltă treptat perete celular secundar. Este dificil să se determine și să poată distinge învelișul primar de cel secundar cu suficientă precizie, deoarece acestea sunt interconectate de mai multe straturi intermediare.
În vrac peretele celular care și-a încheiat formarea este membrana secundară. Acesta oferă celulei forma sa finală. După ce celula este împărțită în două celule fiice, celulele noi cresc, volumul lor crește și forma lor se schimbă; celulele sunt adesea alungite. În același timp, are loc o creștere a grosimii membranei celulare și o restructurare a structurii sale interne.
În timpul perioadei de alungire, fibrilele încep să fie așezate în unghi drept unul față de celălalt și în cele din urmă devin alungite mai mult sau mai puțin paralel cu axa longitudinală a celulei. Procesul se desfășoară constant: în straturile vechi (mai aproape de centrul cochiliei), fibrilele suferă deplasări pasive, iar depunerea de noi fibrile în straturile interioare (cel mai apropiate de membranele celulare) continuă în conformitate cu originalul. planul de proiectare a carcasei. Acest proces creează posibilitatea ca fibrilele să alunece unele față de altele, iar rearanjarea armăturii membranei celulare este posibilă datorită stării gelatinoase a componentelor matricei sale. Ulterior, la înlocuirea ligninei cu hemiceluloza din matrice mobilitatea fibrilelor scade brusc, coaja devine densă, apare lignificarea. Conţinut diverse substanțe cam asa: foarte putina apa, celuloza 40-50%, lignina 25-30%, hemiceluloza 20-30% si practic fara pectina.
Învelișul secundar nu este întotdeauna distanțat uniform. În unele celule specializate conducătoare de apă, are forma inele sau benzi spiralate. Astfel de celule își păstrează capacitatea de a se întinde longitudinal chiar și după moarte.
Se găsește adesea sub membrana secundară înveliș terțiar, care poate fi considerată ca o rămășiță uscată a stratului degenerat al citoplasmei în sine.
Peretele celular (peretele celular)- o trăsătură caracteristică a unei celule vegetale care o deosebește de celula animală. Peretele celular conferă celulei o formă specifică. Cultivate pe medii nutritive speciale, celulele vegetale, în care peretele este îndepărtat prin metoda enzimatică, iau întotdeauna o formă sferică. Peretele celular conferă rezistență celulei și protejează protoplastul, echilibrează presiunea turgenței și astfel previne ruperea plasmalemei. Totalitatea pereților celulari formează un schelet intern care susține corpul plantei și îi conferă rezistență mecanică.
Peretele celular este incolor și transparent, transmite cu ușurință lumina soarelui. De obicei, pereții sunt saturati cu apă. Sistemul pereților celulari transportă apa și compușii cu greutate moleculară mică dizolvați în el (transportul de-a lungul apoplastului).
Peretele celular este compus în principal din polizaharide, care pot fi subdivizate în substanțe ale scheletuluiȘi substanțe ale matricei.
substanță scheletică peretele celular al plantei este celuloza (fibre), care este beta-1,4-D-glucan. Este cea mai abundentă materie organică din biosferă. Moleculele de celuloză sunt lanțuri neramificate foarte lungi, sunt dispuse paralel între ele în grupuri de câteva zeci și sunt ținute împreună prin numeroase legături de hidrogen. Ca urmare, microfibrile, care creează cadrul structural al peretelui și determină rezistența acestuia. Microfibrilele de celuloză sunt vizibile numai la microscopul electronic, diametrul lor este de 10-30 nm, lungimea ajunge la câțiva microni.
Celuloza este insolubilă și nu se umflă în apă. Este foarte inert din punct de vedere chimic, nu se dizolvă în solvenți organici, alcalii concentrați și acizi diluați. Microfibrilele de celuloză sunt elastice și foarte rezistente la rupere (asemănătoare cu oțelul). Aceste proprietăți determină utilizarea pe scară largă a celulozei și a produselor acesteia. Producția mondială de fibre de bumbac, care constă aproape în întregime din celuloză, este de 1,5 10 7 tone pe an. Din celuloză se obțin pulbere fără fum, acetat de mătase și viscoză, celofan și hârtie. Reacție calitativă pe celuloză se efectuează cu un reactiv clor-zinc-iod, peretele celular de celuloză se colorează în albastru-violet.
La ciuperci, substanța scheletică a peretelui celular este chitină- o polizaharidă construită din reziduuri de glucozamină. Chitina este chiar mai durabilă decât celuloza.
Microfibrilele sunt scufundate într-un amorf matrice, care este de obicei un gel de plastic saturat cu apă. Matricea este un amestec complex de polizaharide, ale căror molecule sunt compuse din reziduuri de mai multe zaharuri diferite și sunt mai scurte decât cele de celuloză și lanțuri ramificate. Polizaharidele matricei determină astfel de proprietăți ale peretelui celular, cum ar fi umflarea puternică, permeabilitatea ridicată la apă și compușii cu greutate moleculară mică dizolvați în acesta și proprietățile de schimb de cationi. Polizaharidele matriceale sunt împărțite în două grupe - substanțe pectiniceȘi hemicelulozelor.
substanțe pectinice se umfla sau se dizolva in apa. Ele sunt ușor distruse de alcalii și acizi. Cei mai simpli reprezentanți ai substanțelor pectinice sunt cei solubili în apă. acizi pectici- produși de polimerizare ai acidului alfa-D-galacturonic (până la 100 de unități) legați prin legături 1,4 în lanțuri liniare (alfa-1,4-D-galacturonan). Acizi pectici (pectine)- Aceștia sunt compuși polimerici cu greutate moleculară mai mare (100-200 unități) ai acidului alfa-D-galacturonic, în care grupările carboxil sunt parțial metilate. PectateȘi pectinează– săruri de calciu și magneziu ale acizilor pectic și pectinic. Acizii pectici, pectații și pectinații sunt solubili în apă în prezența zaharurilor și a acizilor organici cu formarea de geluri dense.
Pereții celulari vegetali conțin în principal protopectine- polimeri cu moleculară înaltă ai acidului poligalacturonic metoxilat cu arabinani și galactani; la plantele dicotiledonate, lanțurile galacturonanului conțin o cantitate mică de ramnoză. Protopectinele sunt insolubile în apă.
Hemiceluloze sunt lanțuri ramificate construite din reziduuri neutre de zahăr, glucoză, galactoză, manoză, xiloză sunt mai frecvente; grad de polimerizare 50-300. Hemicelulozele sunt mai stabile din punct de vedere chimic decât substanțele pectinice, sunt mai greu de hidrolizat și se umflă mai puțin în apă. Hemicelulozele pot fi depuse în pereții celulari ai semințelor ca substanțe de rezervă (palmier curmal, curmal). Substanțele pectice și hemicelulozele sunt conectate prin tranziții reciproce. Pe lângă polizaharide, în matricea pereților celulari este prezentă o proteină structurală specială. Este asociat cu reziduuri de zahăr arabinoză și, prin urmare, este o glicoproteină.
Polizaharidele matricei fac mai mult decât umple golurile dintre microfibrilele de celuloză. Lanțurile lor sunt aranjate ordonat și formează numeroase legături atât între ele, cât și cu microfibrile, ceea ce crește semnificativ rezistența peretelui celular.
Pereții celulelor plantelor suferă adesea modificări chimice. Lucrărire, sau lignificarea apare atunci când matricea este depusă lignină- un compus polimeric de natura fenolica, insolubil in apa. Peretele celular lignificat își pierde elasticitatea, duritatea și rezistența la compresiune cresc brusc, iar permeabilitatea la apă scade. Reactivii pentru lignină sunt: 1) floroglucinolȘi clorhidric concentrat sau acid sulfuric(pereții lignificati capătă o culoare roșu-vișiniu) și 2) sulfat anilină, sub influența căreia pereții lignificati devin galben lamaie. Lignificarea este caracteristică pereților celulari ai țesutului conductor al xilemului (lemnului) și țesutului mecanic al sclerenchimului.
Plută, sau suberinizare apare ca urmare a depunerii de polimeri hidrofobi în interiorul peretelui celular - suberinaȘi ceară. Suberina este un amestec de esteri polimerici ai acizilor grași. Monomerii de ceară sunt alcooli grași și esteri de ceară. Ceara este ușor îndepărtată de solvenți organici și se topește rapid și formează cristale. Suberina este un compus amorf care nu se topește și nu se dizolvă în solvenți organici. Suberina și ceara, formând straturi paralele alternative, căptușesc întreaga cavitate a celulei din interior sub forma unui film. Filmul de suberină este practic impermeabil la apă și gaze; prin urmare, după formarea sa, celula moare de obicei. Pluta este caracteristică pereților celulari ai țesutului tegumentar al plută. Reactivul pentru peretele celular suberizat este SudanIII, culoare portocaliu-rosu.
Cutinizarea expus la pereții exteriori ai celulelor țesutului tegumentar al epidermei. KutinȘi ceară depus în straturi alternative pe suprafața exterioară a peretelui celular sub formă de peliculă - cuticulelor. Cutina este un compus polimer asemănător grăsimii similare ca natură chimică și proprietăți cu suberinei. Cuticula protejează planta de evaporarea excesivă a apei de la suprafața plantei. Îl poți colora cu un reactiv SudanIIIîn roșu portocaliu.
Mineralizare peretele celular se datorează depunerii în matrice un numar mare minerale, cel mai adesea silice (oxid de siliciu), mai rar oxalat și carbonat de calciu. Minerale da peretelui duritate si fragilitate. Depunerea de silice este caracteristică celulelor epidermei de coada-calului, rogoz și ierburi. Rigiditatea tulpinilor și a frunzelor dobândite ca urmare a silicificării servește ca agent de protecție împotriva melcilor și, de asemenea, reduce semnificativ palatabilitatea și valoarea nutritivă a plantelor.
Unele celule specializate au slăbire perete celular. În acest caz, în locul unui perete secundar de celuloză, se depun polizaharide acide amorfe, foarte hidratate, sub formă slimeȘi gumă, apropiată ca natură chimică de substanțele pectinice. Mucusul se dizolvă bine în apă cu formarea de soluții mucoase. Gingiile sunt lipicioase și se întind în fire. Când sunt uscate, au o textură excitată. Odată cu depunerea de mucus, protoplastul este împins treptat în centrul celulei, volumul acestuia și volumul vacuolei scad treptat. În cele din urmă, cavitatea celulei se poate umple complet cu mucus, iar celula moare. În unele cazuri, mucusul poate trece prin peretele celular primar la suprafață. Aparatul Golgi are rolul principal în sinteza și secreția de mucus.
Mucusul secretat de celulele vegetale îndeplinește diverse funcții. Astfel, mucusul capacului rădăcinii servește ca lubrifiant, facilitând creșterea vârfului rădăcinii în sol. Glandele mucoase ale plantelor insectivore (roua) secretă un mucus de capcană de care se lipesc insectele. Mucusul secretat de celulele exterioare ale învelișului semințelor (in, gutui, pătlagină) fixează sămânța pe suprafața solului și protejează răsadul de uscare. Mucus colorat cu reactiv albastru de metil in albastru.
Izolarea gingiilor are loc de obicei atunci când plantele sunt rănite. De exemplu, scurgerile de gumă din zonele rănite ale trunchiurilor și ramurilor sunt adesea observate la cireșe și prune. Adezivul de cireșe este o gumă întărită. Guma îndeplinește o funcție de protecție, închizând rana de la suprafață. Guma se formează în principal în plantele lemnoase din familiile de leguminoase (salcâm, astragalus tragacanth) și rosaceae din subfamilia prunelor (cireș, prun, cais). Guma și mucusul sunt folosite în medicină.
Peretele celular este un produs rezidual al protoplastului. În aparatul Golgi se formează polizaharidele matriceale, glicoproteina de perete, lignina și mucusul. Sinteza celulozei, formarea și orientarea microfibrilelor sunt efectuate de plasmalemă. Un rol important în orientarea microfibrilelor revine microtubulilor, care sunt localizați paralel cu microfibrilele depuse în apropierea plasmalemei. Dacă microtubulii sunt distruși, se formează doar celule izodiametrice.
Formarea peretelui celular începe în timpul diviziunii celulare. În planul de diviziune se formează o placă celulară, un singur strat comun celor două celule fiice. Se compune din substanțe pectinice cu consistență semi-lichidă; celuloza este absentă. Într-o celulă adultă, placa celulară se păstrează, dar suferă modificări, motiv pentru care se numește mijloc, sau placă intercelulară (substanță intercelulară) (orez. 2.16). Placa de mijloc este de obicei foarte subțire și aproape imposibil de distins.
Imediat după formarea plăcii celulare, protoplastele celulelor fiice încep să-și așeze propriul perete celular. Se depune din interior atât pe suprafața plăcii celulare, cât și pe suprafața altor pereți celulari care au aparținut anterior celulei mamă. După diviziunea celulară, celula intră în faza de expansiune a creșterii, care se datorează absorbției osmotice intense a apei de către celulă, asociată cu formarea și creșterea vacuolei centrale. Presiunea turgenței începe să întindă peretele, dar nu se rupe datorită faptului că în el se depun în mod constant noi porțiuni de microfibrile și substanțe matrice. Depunerea de noi porțiuni de material are loc uniform pe întreaga suprafață a protoplastei, astfel încât grosimea peretelui celular nu scade.
Se numesc pereții celulelor în diviziune și creștere primar. Conțin multă (60-90%) apă. Substanța uscată este dominată de polizaharide de matrice (60-70%), conținutul de celuloză nu depășește 30%, iar lignina este absentă. Grosimea peretelui primar este foarte mică (0,1-0,5 microni).
Pentru multe celule, depunerea peretelui celular încetează în același timp cu încetarea creșterii celulare. Astfel de celule sunt înconjurate de un perete primar subțire până la sfârșitul vieții ( orez. 2.16).
Orez. 2.16. Celulă parenchimoasă cu perete primar.
În alte celule, depunerea peretelui continuă chiar și după ce celula atinge dimensiunea finală. În acest caz, grosimea peretelui crește, iar volumul ocupat de cavitatea celulară scade. Un astfel de proces se numește îngroșare secundară ziduri, iar zidul însuși se numește secundar(orez. 2.17).
Peretele secundar poate fi considerat ca unul suplimentar, îndeplinind în principal o funcție mecanică, de susținere. Este peretele secundar care este responsabil pentru proprietățile lemnului, fibrelor textile și hârtiei. Peretele secundar conține semnificativ mai puțină apă decât cel primar; este dominată de microfibrile de celuloză (40-50% în greutate substanță uscată), care sunt paralele între ele. Dintre polizaharidele matricei, hemicelulozele (20-30%) sunt caracteristice, există foarte puține substanțe pectinice. Pereții celulari secundari, de regulă, suferă lignificare. În pereții secundari nelignificati (fibre de liban de in, fire de păr de bumbac), conținutul de celuloză poate ajunge la 95%. Conținutul ridicat și orientarea strict ordonată a microfibrilelor determină un nivel ridicat proprietăți mecanice pereții secundari. Adesea, celulele care au un perete celular lignificat secundar mor după terminarea îngroșării secundare.
Lamina mediană lipește celulele adiacente. Dacă se dizolvă, pereții celulari pierd contactul unul cu celălalt și se separă. Acest proces se numește macerare. Macerarea naturală este destul de comună, în care substanțele pectinice ale plăcii medii sunt transformate într-o stare solubilă cu ajutorul enzimei pectinaze și apoi spălate cu apă (fructe supracoapte ale unei pere, pepene galben, piersici, banane). Se observă adesea macerarea parțială, în care placa mediană nu se dizolvă pe toată suprafața, ci doar în colțurile celulelor. Datorită presiunii turgenței, celulele învecinate din aceste locuri sunt rotunjite, rezultând formarea de spatii intercelulare(orez. 2.16). Spațiile intercelulare formează o singură rețea ramificată, care este umplută cu vapori de apă și gaze. Astfel, spațiile intercelulare îmbunătățesc schimbul de gaze celulare.
O trăsătură caracteristică a peretelui secundar este depunerea sa neuniformă pe partea superioară a peretelui primar, drept urmare zonele neîngroșate rămân în peretele secundar - porii. Dacă peretele secundar nu atinge o grosime mare, porii arată ca niște mici depresiuni. În celulele cu un perete secundar puternic, porii din secțiune arată ca niște canale radiale care se extind de la cavitatea celulară la peretele primar. În funcție de forma canalului porilor, se disting două tipuri de pori - simpluși despre mărginit(Fig. 2.17).

Orez. 2.17. Tipuri de pori: A - celule cu pereți secundari și numeroși pori simpli; B – pereche de pori simpli; C, pereche de pori mărginiți.
La pori simpli diametrul canalului porilor pe toată lungimea este același și are forma unui cilindru îngust. Porii simpli sunt caracteristici celulelor parenchimatoase, a fibrelor de libră și de lemn.
Porii din două celule adiacente, de regulă, apar unul vizavi de celălalt. Aceste porii comuni au forma unui singur canal, despărțit printr-o partiție subțire a plăcii mediane și a peretelui primar. Se numește o astfel de combinație de doi pori ai pereților adiacenți ai celulelor învecinate cupluri de poriși funcționează ca una. Secțiunea de perete care le desparte se numește film de închidere porii, sau membrana poroasa. În celulele vii, pelicula de închidere a porilor este pătrunsă cu numeroase plasmodesmate(orez. 2.18).
Plasmodesmate se găsește doar în celulele vegetale. Sunt fire de citoplasmă care traversează peretele celulelor adiacente. Numărul de plasmodesme dintr-o celulă este foarte mare - de la câteva sute la zeci de mii, de obicei plasmodesmele sunt colectate în grupuri. Diametrul canalului plasmodesmenal este de 30-60 nm. Pereții săi sunt căptușiți cu plasmalemă, continuu cu plasmalema celulelor adiacente. Un cilindru membranos trece prin centrul plasmodesmelor. tija centrală plasmodesmate, continuu cu elementele membranare ale reticulului endoplasmatic al ambelor celule. Intre tija centrala si plasmalema din canal se afla o hialoplasma, continua cu hialoplasma celulelor adiacente.

Orez. 2.18. Plasmodesmă la microscop electronic (schemă): 1 - pe o secţiune longitudinală; 2 - pe o secțiune transversală; pl- plasmalema; CA- nucleul central al plasmodesmelor; ER- un element al reticulului endoplasmatic.
Astfel, protoplastele celulare nu sunt complet izolate unele de altele, ci comunică prin canale plasmodesmate. Prin intermediul acestora are loc transportul intercelular al ionilor și al moleculelor mici, precum și se transmit stimuli hormonali. Prin plasmodesmate, protoplastele celulelor dintr-un organism vegetal formează un singur întreg, numit simptom, iar transportul substanțelor prin plasmodesme se numește simplistic Spre deosebire de apoplastic transportul de-a lungul pereților celulari și a spațiilor intercelulare.
La pori franjuri(orez. 2.17) canalul se îngustează brusc în procesul de depunere a peretelui celular; prin urmare, deschiderea interioară a porului, care se deschide în cavitatea celulară, este mult mai îngustă decât cea exterioară, care se sprijină pe peretele primar. Porii mărginiți sunt caracteristici celulelor de moarte timpurie ale elementelor conductoare de apă ale lemnului. Canalul lor de pori se extinde în formă de pâlnie spre filmul de închidere, iar peretele secundar atârnă sub formă de rolă peste partea expandată a canalului, formând o cameră de pori. Denumirea porului mărginit provine de la faptul că, văzută de la suprafață, gaura interioară arată ca un cerc mic sau o fantă îngustă, în timp ce orificiul exterior, parcă, îl mărginește pe cel interior sub forma unui cerc de un diametru mai mare sau o fantă mai largă.
Porii facilitează transportul apei și substanțelor dizolvate de la celulă la celulă, fără a compromite rezistența peretelui celular.