Zelluläre Mitose. Mitose
Lehrbuch für die Klassen 10-11
Abschnitt II. Fortpflanzung und Entwicklung von Organismen
Kapitel V. Fortpflanzung von Organismen
Jede Sekunde stirbt auf der Erde eine astronomische Zahl von Lebewesen an Altersschwäche, Krankheiten und Raubtieren, und nur dank der Fortpflanzung, dieser universellen Eigenschaft von Organismen, hört das Leben auf der Erde nicht auf.
Es mag den Anschein haben, dass die Fortpflanzungsprozesse bei Lebewesen sehr unterschiedlich sind, aber alle können auf zwei Formen reduziert werden: asexuell und sexuell. Einige Organismen haben verschiedene Formen Reproduktion. Beispielsweise können sich viele Pflanzen durch Stecklinge, Schichten, Knollen (asexuelle Vermehrung) und Samen (sexuelle Vermehrung) vermehren.
Bei der sexuellen Fortpflanzung entwickelt sich jeder Organismus aus einer Zelle, die aus der Verschmelzung zweier Geschlechtszellen – männlich und weiblich – entsteht.
Grundlage der Fortpflanzung und individuellen Entwicklung eines Organismus ist der Prozess der Zellteilung.
§ 20. Zellteilung. Mitose
Die Teilungsfähigkeit ist die wichtigste Eigenschaft von Zellen. Ohne Teilung ist eine Zunahme der Zahl einzelliger Lebewesen, die Entwicklung eines komplexen vielzelligen Organismus aus einer befruchteten Eizelle, die Erneuerung von Zellen, Geweben und sogar Organen, die im Laufe des Lebens des Organismus verloren gehen, nicht vorstellbar.
Die Zellteilung erfolgt stufenweise. In jeder Teilungsstufe finden bestimmte Prozesse statt. Sie führen zur Verdoppelung des genetischen Materials (DNA-Synthese) und seiner Verteilung auf Tochterzellen. Der Zeitraum des Zelllebens von einer Teilung zur nächsten wird Zellzyklus genannt.
Vorbereitung auf die Teilung. Eukaryontische Organismen, die aus Zellen mit Kernen bestehen, beginnen in einem bestimmten Stadium des Zellzyklus, in der Interphase, mit der Vorbereitung auf die Teilung.
In der Interphase findet in der Zelle der Prozess der Proteinbiosynthese statt und die Chromosomen verdoppeln sich. Entlang des ursprünglichen Chromosoms wird aus den in der Zelle vorhandenen chemischen Verbindungen eine exakte Kopie davon synthetisiert und das DNA-Molekül verdoppelt. Ein doppeltes Chromosom besteht aus zwei Hälften – Chromatiden. Jedes Chromatid enthält ein DNA-Molekül.
Die Interphase in pflanzlichen und tierischen Zellen dauert durchschnittlich 10–20 Stunden. Dann beginnt der Prozess der Zellteilung – Mitose.
Während der Mitose durchläuft die Zelle eine Reihe aufeinanderfolgender Phasen, in deren Folge jede Tochterzelle denselben Chromosomensatz erhält wie die Mutterzelle.
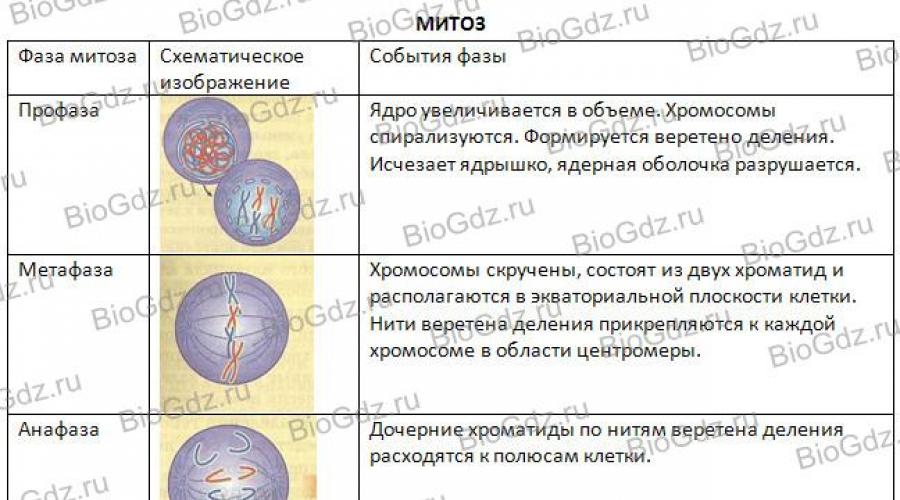
Phasen der Mitose. Es gibt vier Phasen der Mitose: Prophase, Metaphase, Anaphase und Telophase. Abbildung 29 zeigt schematisch den Verlauf der Mitose. In der Prophase sind Zentriolen deutlich sichtbar – Formationen, die sich im Zellzentrum befinden und eine Rolle bei der Divergenz der Tochterchromosomen von Tieren spielen. (Denken Sie daran, dass nur einige Pflanzen Zentriolen im Zellzentrum haben, die die Trennung der Chromosomen organisieren.) Wir werden die Mitose anhand des Beispiels betrachten Tierzelle, da das Vorhandensein eines Zentriols den Prozess der Chromosomentrennung sichtbarer macht. Die Zentriolen verdoppeln sich und wandern zu verschiedenen Polen der Zelle. Mikrotubuli gehen von den Zentriolen aus und bilden Filamente der Spindel, die die Divergenz der Chromosomen zu den Polen der sich teilenden Zelle regulieren.
Reis. 29. Schema der Mitose
Am Ende der Prophase löst sich die Kernmembran auf, der Nukleolus verschwindet allmählich, die Chromosomen spiralen und verkürzen und verdicken sich dadurch, und sie sind bereits unter dem Lichtmikroskop zu beobachten. Sie sind im nächsten Stadium der Mitose – der Metaphase – noch besser sichtbar.
In der Metaphase befinden sich die Chromosomen in der Äquatorialebene der Zelle. Es ist deutlich zu erkennen, dass jedes Chromosom, bestehend aus zwei Chromatiden, eine Verengung – ein Zentromer – aufweist. Chromosomen sind mit ihren Zentromeren an den Spindelfilamenten befestigt. Nach der Zentromerteilung wird jedes Chromatid zu einem unabhängigen Tochterchromosom.
Dann kommt die nächste Phase der Mitose – die Anaphase, in der die Tochterchromosomen (Chromatiden eines Chromosoms) zu verschiedenen Zellpolen auseinanderlaufen.
Die nächste Stufe der Zellteilung ist die Telophase. Sie beginnt, nachdem die Tochterchromosomen, bestehend aus einem Chromatid, die Pole der Zelle erreicht haben. In diesem Stadium despirieren die Chromosomen erneut und nehmen das gleiche Aussehen an wie vor Beginn der Zellteilung in der Interphase (lange dünne Fäden). Um sie herum entsteht eine Kernhülle, und im Kern bildet sich ein Nukleolus, in dem Ribosomen synthetisiert werden. Während des Prozesses der zytoplasmatischen Teilung werden alle Organellen (Mitochondrien, Golgi-Komplex, Ribosomen usw.) mehr oder weniger gleichmäßig auf die Tochterzellen verteilt.
Durch die Mitose werden also aus einer Zelle zwei, von denen jede eine für einen bestimmten Organismustyp charakteristische Anzahl und Form von Chromosomen und damit eine konstante Menge an DNA aufweist.
Der gesamte Mitoseprozess dauert durchschnittlich 1-2 Stunden. Die Dauer variiert geringfügig verschiedene Typen Zellen. Es kommt auch auf die Bedingungen an Außenumgebung(Temperatur, Lichtverhältnisse und andere Indikatoren).
Die biologische Bedeutung der Mitose besteht darin, dass sie die Konstanz der Chromosomenzahl in allen Körperzellen gewährleistet. Beim Prozess der Mitose wird die DNA der Chromosomen der Mutterzelle streng gleichmäßig auf die beiden daraus entstehenden Tochterzellen verteilt. Durch die Mitose erhalten alle Tochterzellen die gleichen genetischen Informationen.
- Welche Veränderungen in einer Zelle gehen der Zellteilung voraus?
- Wann entsteht die Spindel? Was ist seine Rolle?
- Beschreiben Sie die Phasen der Mitose und beschreiben Sie kurz, wie dieser Prozess abläuft.
- Was ist Chromatid? Wann wird es zum Chromosom?
- Was ist ein Zentromer? Welche Rolle spielt es bei der Mitose?
- Was ist biologische Bedeutung Mitose?
Erinnern Sie sich aus dem Studium der Botanik, Zoologie, Anatomie, Physiologie und menschlichen Hygiene daran, wie die Fortpflanzung in der organischen Welt erfolgt.
1. Geben Sie Definitionen von Konzepten an.
Interphase– die Phase der Vorbereitung auf die mitotische Teilung, in der die DNA-Duplikation stattfindet.
Mitose- Hierbei handelt es sich um eine Teilung, die zu einer streng identischen Verteilung exakt kopierter Chromosomen zwischen den Tochterzellen führt und so die Bildung genetisch identischer Zellen gewährleistet.
Lebenszyklus
- die Lebensdauer einer Zelle vom Zeitpunkt ihrer Entstehung im Teilungsprozess bis zum Tod oder dem Ende der nachfolgenden Teilung.
2. Wie unterscheidet sich das Wachstum einzelliger Organismen vom Wachstum mehrzelliger Organismen?
Das Wachstum eines einzelligen Organismus ist eine Zunahme der Größe und Komplikation der Struktur einer einzelnen Zelle, und das Wachstum eines mehrzelligen Organismus ist auch die aktive Teilung von Zellen – eine Zunahme ihrer Anzahl.
3. Warum gibt es im Lebenszyklus einer Zelle notwendigerweise eine Interphase?
In der Interphase erfolgt die Vorbereitung auf die Teilung und DNA-Vervielfältigung. Geschieht dies nicht, würde sich bei jeder Zellteilung die Zahl der Chromosomen halbieren, und schon bald gäbe es überhaupt keine Chromosomen mehr in der Zelle.
4. Vervollständigen Sie den Cluster „Phasen der Mitose“.
5. Füllen Sie die Tabelle unter Verwendung von Abbildung 52 in § 3.4 aus.

6. Erfinden Sie einen Syncwine für den Begriff „Mitose“.
Mitose
Vierphasig, gleichmäßig
Teilt, verteilt, zerkleinert
Versorgt Tochterzellen mit genetischem Material
Zellteilung.
7. Stellen Sie eine Entsprechung zwischen den Phasen des Mitosezyklus und den in ihnen stattfindenden Ereignissen her.
Phasen
1. Anaphase
2. Metaphase
3. Interphase
4. Telophase
5. Prophase
Veranstaltungen
A. Die Zelle wächst, Organellen werden gebildet, DNA verdoppelt sich.
B. Chromatiden divergieren und werden zu unabhängigen Chromosomen.
B. Die Spiralisierung der Chromosomen beginnt und die Kernmembran wird zerstört.
D. Chromosomen befinden sich in der Äquatorialebene der Zelle. Die Spindelfilamente sind an den Zentromeren befestigt.
D. Die Spindel verschwindet, Kernmembranen bilden sich, Chromosomen lösen sich.
8. Warum verläuft der Abschluss der Mitose – die Teilung des Zytoplasmas – in tierischen und pflanzlichen Zellen unterschiedlich?
Nicht in tierischen Zellen Zellenwand, bei ihnen Zellmembran ragt nach innen und die Zelle teilt sich durch Verengung.
In Pflanzenzellen bildet sich die Membran in der Äquatorebene im Inneren der Zelle und teilt die Zelle, indem sie sich zur Peripherie hin ausbreitet, in zwei Hälften.
9. Warum dauert die Interphase im Mitosezyklus viel länger als die Teilung selbst?
Während der Interphase bereitet sich die Zelle intensiv auf die Mitose vor, in ihr finden Syntheseprozesse statt, DNA wird dupliziert, die Zelle wächst, durchläuft Lebenszyklus, ohne die Abteilung selbst.
10. Wählen Sie die richtige Antwort.
Test 1.
Als Ergebnis der Mitose produziert eine diploide Zelle:
4) 2 diploide Zellen.
Test 2.
Die Teilung der Zentromere und die Divergenz der Chromatiden zu den Zellpolen erfolgt in:
3) Anaphase;
Test 3.
Der Lebenszyklus ist:
2) das Leben einer Zelle von der Teilung bis zum Ende der nächsten Teilung oder zum Tod;
Test 4.
Welcher Begriff ist falsch geschrieben?
4) Telophase.
11. Erklären Sie den Ursprung und allgemeine Bedeutung Wörter (Begriffe), basierend auf der Bedeutung der Wurzeln, aus denen sie bestehen.

12. Wählen Sie einen Begriff aus und erklären Sie, wie seine moderne Bedeutung mit der ursprünglichen Bedeutung seiner Wurzeln übereinstimmt.
Der gewählte Begriff ist Interphase.
Korrespondenz. Der Begriff entspricht und bezieht sich auf den Zeitraum zwischen den Phasen der Mitose, in dem die Vorbereitung auf die Teilung erfolgt.
13. Formulieren und notieren Sie die Hauptgedanken von § 3.4.
Der Lebenszyklus ist das Leben einer Zelle von der Teilung bis zum Ende der nächsten Teilung oder zum Tod. Zwischen den Teilungen bereitet sich die Zelle während der Interphase darauf vor. Zu diesem Zeitpunkt findet die Substanzsynthese statt, die DNA-Verdoppelung.
Die Zellteilung erfolgt durch Mitose. Es besteht aus 4 Phasen:
Prophase.
Metaphase.
Anaphase.
Telophase.
Der Zweck der Mitose: Dadurch werden aus einer Mutterzelle zwei Tochterzellen mit identischem Gensatz gebildet. Die Menge an genetischem Material und Chromosomen bleibt gleich und gewährleistet so die genetische Stabilität der Zellen.
Der wichtigste Bestandteil des Zellzyklus ist der mitotische (proliferative) Zyklus. Es handelt sich um einen Komplex miteinander verbundener und koordinierter Phänomene während der Zellteilung sowie davor und danach. Der Mitosezyklus besteht aus einer Reihe von Prozessen, die in einer Zelle von einer Zellteilung zur nächsten ablaufen und mit der Bildung von zwei Zellen der nächsten Generation enden. Darüber hinaus umfasst der Begriff des Lebenszyklus auch den Zeitraum, in dem die Zelle ihre Funktionen erfüllt, und Ruhephasen. Zu diesem Zeitpunkt ist das weitere Schicksal der Zelle ungewiss: Die Zelle kann beginnen, sich zu teilen (in die Mitose einzutreten) oder sich auf die Ausführung bestimmter Funktionen vorzubereiten.
Hauptstadien der Mitose
1.
Reduktion (Selbstverdopplung) der genetischen Information der Mutterzelle und deren gleichmäßige Verteilung zwischen den Tochterzellen. Damit gehen Veränderungen in der Struktur und Morphologie der Chromosomen einher, in denen mehr als 90 % der Informationen einer eukaryontischen Zelle konzentriert sind.
2.
Der Mitosezyklus besteht aus vier aufeinanderfolgenden Perioden: präsynthetischer (oder postmitotischer) G1, synthetischer S, postsynthetischer (oder prämitotischer) G2 und Mitose selbst. Sie bilden die autokatalytische Interphase (Vorbereitungsphase).
Phasen des Zellzyklus:
1) präsynthetisch (G1). Tritt unmittelbar nach der Zellteilung auf. Die DNA-Synthese hat noch nicht stattgefunden. Die Zelle wächst aktiv an Größe und speichert für die Teilung notwendige Substanzen: Proteine (Histone, Strukturproteine, Enzyme), RNA, ATP-Moleküle. Es kommt zur Teilung von Mitochondrien und Chloroplasten (d. h. Strukturen, die zur Selbstreproduktion fähig sind). Die organisatorischen Merkmale der Interphasenzelle werden nach der vorherigen Teilung wiederhergestellt;
2)
synthetisch (S). Genetisches Material wird durch DNA-Replikation vervielfältigt. Es geschieht auf halbkonservative Weise, wenn die Doppelhelix des DNA-Moleküls in zwei Ketten zerfällt und auf jeder von ihnen eine komplementäre Kette synthetisiert wird.
Als Ergebnis zwei identische Doppelhelix DNA, von denen jede aus einem neuen und besteht alte Kette DNA. Die Menge an Erbmaterial verdoppelt sich. Darüber hinaus wird die Synthese von RNA und Proteinen fortgesetzt. Außerdem wird ein kleiner Teil der mitochondrialen DNA repliziert (der größte Teil davon wird in der G2-Periode repliziert);
3) postsynthetisch (G2). DNA wird nicht mehr synthetisiert, aber die bei ihrer Synthese in der S-Periode entstandenen Defekte werden korrigiert (Reparatur). Energie und Nährstoffe, die Synthese von RNA und Proteinen (hauptsächlich Kernproteine) geht weiter.
S und G2 stehen in direktem Zusammenhang mit der Mitose und werden daher manchmal in eine separate Periode unterteilt – die Präprophase.
Danach erfolgt die eigentliche Mitose, die aus vier Phasen besteht. Der Teilungsprozess umfasst mehrere aufeinanderfolgende Phasen und ist ein Zyklus. Ihre Dauer variiert und liegt in den meisten Zellen zwischen 10 und 50 Stunden. In menschlichen Körperzellen beträgt die Dauer der Mitose selbst 1–1,5 Stunden, die G2-Interphase 2–3 Stunden und die S-Interphase 6–10 Stunden Std .
Die Dauer der einzelnen Stadien ist unterschiedlich und variiert je nach Gewebeart, physiologischer Zustand Körper, externe Faktoren. Die längsten Stadien sind mit den Prozessen der intrazellulären Synthese verbunden: Prophase und Telophase. Die schnellsten Phasen der Mitose, in denen die Bewegung der Chromosomen stattfindet: Metaphase und Anaphase. Der eigentliche Prozess der Chromosomendivergenz zu den Polen dauert in der Regel nicht länger als 10 Minuten.
Prophase
Zu den Hauptereignissen der Prophase gehören die Kondensation von Chromosomen im Zellkern und die Bildung einer Teilungsspindel im Zytoplasma der Zelle. Der Zerfall des Nukleolus in der Prophase ist ein charakteristisches, aber nicht zwingendes Merkmal für alle Zellen.
Als Beginn der Prophase wird üblicherweise der Zeitpunkt des Auftretens mikroskopisch sichtbarer Chromosomen aufgrund der Kondensation von intranukleärem Chromatin angesehen. Die Chromosomenverdichtung erfolgt aufgrund der mehrstufigen DNA-Helixierung. Diese Veränderungen gehen mit einer Erhöhung der Aktivität von Phosphorylasen einher, die Histone modifizieren, die direkt an der DNA-Zusammensetzung beteiligt sind. Infolgedessen nimmt die Transkriptionsaktivität des Chromatins stark ab, nukleoläre Gene werden inaktiviert und die meisten nukleolären Proteine dissoziieren. Kondensierende Schwesterchromatiden in der frühen Prophase bleiben mit Hilfe von Kohäsinproteinen über ihre gesamte Länge gepaart, aber zu Beginn der Prometaphase bleibt die Verbindung zwischen Chromatiden nur im Zentromerbereich bestehen. In der späten Prophase werden auf jedem Zentromer der Schwesterchromatiden reife Kinetochoren gebildet, die für die Bindung der Chromosomen an die Mikrotubuli der Spindel in der Prometaphase erforderlich sind.
Zusammen mit den Prozessen der intranukleären Kondensation von Chromosomen beginnt sich im Zytoplasma eine mitotische Spindel zu bilden – eine der Hauptstrukturen des Zellteilungsapparats, die für die Verteilung der Chromosomen zwischen Tochterzellen verantwortlich ist. Polkörperchen, Mikrotubuli und Chromosomenkinetochore sind in allen eukaryotischen Zellen an der Bildung der Teilungsspindel beteiligt.
Der Beginn der mitotischen Spindelbildung in der Prophase ist mit dramatischen Veränderungen der dynamischen Eigenschaften der Mikrotubuli verbunden. Die Halbwertszeit eines durchschnittlichen Mikrotubulus verringert sich etwa um das Zwanzigfache von 5 Minuten auf 15 Sekunden. Allerdings erhöht sich ihre Wachstumsrate im Vergleich zu den gleichen Interphase-Mikrotubuli etwa um das Zweifache. Polymerisierende Plusenden sind „dynamisch instabil“ und wechseln abrupt von gleichmäßigem Wachstum zu schneller Verkürzung, bei der häufig die gesamten Mikrotubuli depolymerisieren. Es ist bemerkenswert, dass für das ordnungsgemäße Funktionieren der mitotischen Spindel ein gewisses Gleichgewicht zwischen den Prozessen des Zusammenbaus und der Depolymerisation von Mikrotubuli erforderlich ist, da weder stabilisierte noch depolymerisierte Spindelmikrotubuli in der Lage sind, Chromosomen zu bewegen.
Zusammen mit den beobachteten Veränderungen der dynamischen Eigenschaften der Mikrotubuli, aus denen die Spindelfilamente bestehen, werden in der Prophase Teilungspole gebildet. In der S-Phase replizierte Zentrosomen divergieren aufgrund der Wechselwirkung der zueinander wachsenden Pol-Mikrotubuli in entgegengesetzte Richtungen. Mit ihren Minus-Enden tauchen die Mikrotubuli in die amorphe Substanz der Zentrosomen ein und Polymerisationsprozesse finden von den Plus-Enden aus, die der Äquatorialebene der Zelle zugewandt sind, statt. In diesem Fall wird der wahrscheinliche Mechanismus der Poldivergenz wie folgt erklärt: Dynein-ähnliche Proteine sind nach innen ausgerichtet parallele Richtung Polymerisierende Plusenden polarer Mikrotubuli und Kinesin-ähnliche Proteine wiederum schieben sie in Richtung der Teilungspole.
Parallel zur Kondensation der Chromosomen und der Bildung der mitotischen Spindel kommt es während der Prophase zu einer Fragmentierung des endoplasmatischen Retikulums, das in kleine Vakuolen zerfällt, die dann zur Peripherie der Zelle divergieren. Gleichzeitig verlieren Ribosomen die Verbindung zu den ER-Membranen. Auch die Zisternen des Golgi-Apparats ändern ihre perinukleäre Lokalisierung und zerfallen in einzelne Dictyosomen, die in keiner bestimmten Reihenfolge im Zytoplasma verteilt sind.
Prometaphase
Das Ende der Prophase und der Beginn der Prometaphase sind normalerweise durch den Zerfall der Kernmembran gekennzeichnet. Ganze Zeile Lamina-Proteine werden phosphoryliert, wodurch die Kernhülle in kleine Vakuolen fragmentiert wird und die Porenkomplexe verschwinden. Nach der Zerstörung der Kernmembran befinden sich die Chromosomen ohne besondere Reihenfolge im Kernbereich. Doch bald beginnen sie alle, sich zu bewegen.
In der Prometaphase wird eine intensive, aber zufällige Bewegung der Chromosomen beobachtet. Zunächst driften einzelne Chromosomen schnell mit einer Geschwindigkeit von bis zu 25 μm/min zum nächstgelegenen Pol der mitotischen Spindel. In der Nähe der Teilungspole steigt die Wahrscheinlichkeit einer Wechselwirkung neu synthetisierter Spindel-Mikrotubuli-Plus-Enden mit Chromosomen-Kinetochoren. Durch diese Wechselwirkung werden Kinetochor-Mikrotubuli vor spontaner Depolymerisation stabilisiert und ihr Wachstum sorgt teilweise für die Entfernung des mit ihnen verbundenen Chromosoms in Richtung vom Pol zur Äquatorialebene der Spindel. Auf der anderen Seite wird das Chromosom von Mikrotubulisträngen überholt, die vom gegenüberliegenden Pol der mitotischen Spindel stammen. Durch die Interaktion mit Kinetochoren nehmen sie auch an der Chromosomenbewegung teil. Dadurch werden Schwesterchromatiden mit entgegengesetzten Polen der Spindel assoziiert. Die Kraft, die Mikrotubuli von verschiedenen Polen aus entwickeln, stabilisiert nicht nur die Wechselwirkung dieser Mikrotubuli mit Kinetochoren, sondern bringt letztendlich auch jedes Chromosom in die Ebene der Metaphasenplatte.
In Säugetierzellen erfolgt die Prometaphase normalerweise innerhalb von 10–20 Minuten. Bei Heuschrecken-Neuroblasten dauert dieses Stadium nur 4 Minuten, bei Haemanthus-Endosperm und Molch-Fibroblasten etwa 30 Minuten.
Metaphase
Am Ende der Prometaphase befinden sich die Chromosomen in der Äquatorialebene der Spindel in etwa gleichen Abständen von beiden Teilungspolen und bilden eine Metaphaseplatte. Die Morphologie der Metaphaseplatte in tierischen Zellen zeichnet sich in der Regel durch eine geordnete Anordnung der Chromosomen aus: Die Zentromerregionen sind zur Mitte der Spindel gerichtet, und die Arme sind zur Peripherie der Zelle gerichtet. In Pflanzenzellen liegen Chromosomen oft ohne strenge Reihenfolge in der Äquatorialebene der Spindel.
Die Metaphase nimmt einen erheblichen Teil der Mitoseperiode ein und zeichnet sich durch einen relativ stabilen Zustand aus. Während dieser ganzen Zeit werden die Chromosomen aufgrund der ausgeglichenen Spannungskräfte der Kinetochor-Mikrotubuli in der Äquatorialebene der Spindel gehalten und führen oszillierende Bewegungen mit unbedeutender Amplitude in der Ebene der Metaphasenplatte aus.
In der Metaphase sowie in anderen Phasen der Mitose setzt sich die aktive Erneuerung der Spindelmikrotubuli durch intensive Montage und Depolymerisation von Tubulinmolekülen fort. Trotz einer gewissen Stabilisierung der Bündel von Kinetochor-Mikrotubuli kommt es zu einer ständigen Neuanordnung interpolarer Mikrotubuli, deren Anzahl in der Metaphase ein Maximum erreicht.
Am Ende der Metaphase ist eine deutliche Trennung der Schwesterchromatiden zu beobachten, deren Verbindung nur in den zentromeren Regionen aufrechterhalten bleibt. Die Chromatidarme liegen parallel zueinander und die Lücke zwischen ihnen wird deutlich sichtbar.
Anaphase
Die Anaphase ist das kürzeste Stadium der Mitose, das mit der plötzlichen Trennung und anschließenden Trennung der Schwesterchromatiden in Richtung entgegengesetzter Pole der Zelle beginnt. Chromatiden divergieren mit einer gleichmäßigen Geschwindigkeit von 0,5–2 µm/min und nehmen häufig eine V-Form an. Ihre Bewegung wird durch erhebliche Kräfte angetrieben, die auf 10 Dyn pro Chromosom geschätzt werden, was dem 10.000-fachen der Kraft entspricht, die erforderlich ist, um ein Chromosom einfach mit der beobachteten Geschwindigkeit durch das Zytoplasma zu bewegen.
Typischerweise besteht die Chromosomentrennung in der Anaphase aus zwei relativ unabhängigen Prozessen, die Anaphase A und Anaphase B genannt werden.
Anaphase A ist durch die Trennung von Schwesterchromatiden in entgegengesetzte Pole der Zellteilung gekennzeichnet. Für ihre Bewegung sind dieselben Kräfte verantwortlich, die zuvor die Chromosomen in der Ebene der Metaphaseplatte hielten. Der Prozess der Chromatidtrennung geht mit einer Verringerung der Länge der depolymerisierenden Kinetochor-Mikrotubuli einher. Darüber hinaus wird ihr Zerfall hauptsächlich im Bereich der Kinetochoren, von den Plusenden her, beobachtet. Vermutlich liegt eine Depolymerisation der Mikrotubuli an Kinetochoren oder im Bereich der Teilungspole vor eine notwendige Bedingung für die Bewegung von Schwesterchromatiden, da deren Bewegung stoppt, wenn Taxol oder schweres Wasser hinzugefügt werden, die eine stabilisierende Wirkung auf Mikrotubuli haben. Der Mechanismus, der der Chromosomensegregation in der Anaphase A zugrunde liegt, ist weiterhin unbekannt.
Während der Anaphase B divergieren die Pole der Zellteilung selbst, und im Gegensatz zur Anaphase A erfolgt dieser Prozess durch die Ansammlung polarer Mikrotubuli an den Plusenden. Die polymerisierenden antiparallelen Filamente der Spindel erzeugen bei ihrer Wechselwirkung teilweise eine Kraft, die die Pole auseinanderdrückt. Das Ausmaß der relativen Bewegung der Pole in diesem Fall sowie der Grad der Überlappung der polaren Mikrotubuli in der äquatorialen Zone der Zelle variieren stark zwischen Individuen verschiedener Arten. Zusätzlich zu den Schubkräften werden die Teilungspole durch Zugkräfte von astralen Mikrotubuli beeinflusst, die durch die Wechselwirkung mit Dynein-ähnlichen Proteinen auf der Plasmamembran der Zelle entstehen.
Die Reihenfolge, Dauer und der relative Beitrag jedes der beiden Prozesse, aus denen sich die Anaphase zusammensetzt, können äußerst unterschiedlich sein. So beginnt in Säugetierzellen die Anaphase B unmittelbar nach Beginn der Chromatiddivergenz zu entgegengesetzten Polen und setzt sich fort, bis sich die mitotische Spindel im Vergleich zur Metaphase um das 1,5- bis 2-fache verlängert. In einigen anderen Zellen beginnt die Anaphase B erst, nachdem die Chromatiden die Teilungspole erreicht haben. Bei einigen Protozoen verlängert sich die Spindel während der Anaphase B um das 15-fache im Vergleich zur Metaphase. Anaphase B fehlt in Pflanzenzellen.
Telophase
Die Telophase gilt als letztes Stadium der Mitose; Als Beginn gilt der Moment, in dem die getrennten Schwesterchromatiden an den entgegengesetzten Polen der Zellteilung enden. In der frühen Telophase wird eine Dekondensation der Chromosomen und damit eine Vergrößerung ihres Volumens beobachtet. In der Nähe der gruppierten Einzelchromosomen beginnt die Verschmelzung der Membranvesikel, wodurch der Wiederaufbau der Kernhülle beginnt. Das Material zum Aufbau der Membranen neu gebildeter Tochterkerne sind Fragmente der zunächst zerfallenen Kernmembran der Mutterzelle sowie Elemente des endoplasmatischen Retikulums. Dabei binden sich einzelne Vesikel an die Oberfläche der Chromosomen und verschmelzen miteinander. Die äußeren und inneren Kernmembranen werden nach und nach wiederhergestellt, die Kernschicht und die Kernporen werden wiederhergestellt. Während des Prozesses der Wiederherstellung der Kernmembran verbinden sich wahrscheinlich einzelne Membranvesikel mit der Oberfläche von Chromosomen, ohne spezifische Nukleotidsequenzen zu erkennen, da Experimente gezeigt haben, dass die Wiederherstellung der Kernmembran um DNA-Moleküle herum erfolgt, die von jedem Organismus, sogar einem bakteriellen Virus, stammen. Im Inneren der neu gebildeten Zellkerne wird Chromatin verteilt, die RNA-Synthese wird wieder aufgenommen und Nukleolen werden sichtbar.
Parallel zu den Prozessen der Bildung der Kerne von Tochterzellen in der Telophase beginnt und endet der Abbau der Spindelmikrotubuli. Die Depolymerisation verläuft in Richtung von den Teilungspolen zur Äquatorialebene der Zelle, von Minus-Enden zu Plus-Enden. In diesem Fall verbleiben die Mikrotubuli am längsten im mittleren Teil der Spindel, die den restlichen Fleming-Körper bilden.
Das Ende der Telophase fällt überwiegend mit der Teilung des Körpers der Mutterzelle – der Zytokinese – zusammen. In diesem Fall werden zwei oder mehr Tochterzellen gebildet. Die Prozesse, die zur Trennung des Zytoplasmas führen, beginnen in der Mitte der Anaphase und können nach Abschluss der Telophase fortgesetzt werden. Mitose geht nicht immer mit einer Teilung des Zytoplasmas einher, daher wird die Zytokinese nicht als separate Phase der mitotischen Teilung klassifiziert und normalerweise als Teil der Telophase betrachtet.
Es gibt zwei Haupttypen der Zytokinese: Teilung durch transversale Zellverengung und Teilung durch Bildung einer Zellplatte. Die Ebene der Zellteilung wird durch die Position der mitotischen Spindel bestimmt und verläuft im rechten Winkel zur Längsachse der Spindel.
Wenn sich eine Zelle durch eine Querverengung teilt, wird der Ort der zytoplasmatischen Teilung vorläufig während der Anaphase festgelegt, wenn ein kontraktiler Ring aus Aktin- und Myosinfilamenten in der Ebene der Metaphasenplatte unter der Zellmembran erscheint. Anschließend bildet sich aufgrund der Aktivität des kontraktilen Rings eine Spaltfurche, die sich allmählich vertieft, bis die Zelle vollständig geteilt ist. Am Ende der Zytokinese löst sich der kontraktile Ring vollständig auf und die Plasmamembran zieht sich um einen verbleibenden Fleming-Körper zusammen, der aus einer Ansammlung von Überresten zweier Gruppen polarer Mikrotubuli besteht, die mit dichtem Matrixmaterial eng zusammengepackt sind.
Die Teilung durch Bildung der Zellplatte beginnt mit der Bewegung kleiner membrangebundener Vesikel in Richtung der Äquatorialebene der Zelle. Hier verschmelzen sie und bilden eine scheibenförmige Struktur, die von einer Membran umgeben ist – der frühen Zellplatte. Die kleinen Vesikel stammen hauptsächlich aus dem Golgi-Apparat und bewegen sich entlang der verbleibenden Pol-Mikrotubuli der Spindel in Richtung der Äquatorialebene und bilden eine zylindrische Struktur, die als Phragmoplast bezeichnet wird. Während sich die Zellplatte ausdehnt, bewegen sich gleichzeitig die Mikrotubuli des frühen Phragmoplasten an die Peripherie der Zelle, wo aufgrund neuer Membranvesikel das Wachstum der Zellplatte bis zu ihrer endgültigen Verschmelzung mit der Membran der Mutterzelle fortgesetzt wird. Nach der endgültigen Trennung der Tochterzellen werden Zellulose-Mikrofibrillen in der Zellplatte abgelagert, wodurch die Bildung einer starren Zellwand abgeschlossen wird.
Um den Abschluss jeder Phase des Zellzyklus festzustellen, ist das Vorhandensein von Kontrollpunkten erforderlich. Wenn eine Zelle einen Kontrollpunkt „passiert“, „bewegt“ sie sich weiter durch den Zellzyklus. Wenn bestimmte Umstände, wie zum Beispiel eine DNA-Schädigung, die Zelle daran hindern, einen Kontrollpunkt zu passieren, der mit einer Art Kontrollpunkt verglichen werden kann, dann stoppt die Zelle und eine weitere Phase des Zellzyklus findet nicht statt, zumindest bis die Hindernisse beseitigt sind Dadurch wird verhindert, dass die Zelle den Kontrollpunkt passiert.
Mitotische Zellteilung
Mitose(vom griechischen Mitos – Faden), auch Karyokinese oder indirekte Zellteilung genannt, ist ein universeller Mechanismus der Zellteilung. Die Mitose folgt auf die G2-Periode und schließt den Zellzyklus ab.
Es dauert 1-3 Stunden und sorgt für eine gleichmäßige Verteilung des genetischen Materials in den Tochterzellen. Die Mitose umfasst 4 Hauptphasen: Prophase, Metaphase, Anaphase und Telophase.
Mitose ist einer der grundlegenden Prozesse der Ontogenese. Die mitotische Teilung sorgt für das Wachstum mehrzelliger Eukaryoten, indem sie die Zellpopulationen im Gewebe erhöht.
Durch die mitotische Teilung der Meristemzellen nimmt die Zahl der Pflanzengewebezellen zu. Die Fragmentierung einer befruchteten Eizelle und das Wachstum der meisten Gewebe bei Tieren erfolgen ebenfalls durch mitotische Teilungen.
Basierend auf morphologischen Merkmalen wird die Mitose üblicherweise in Stadien unterteilt: Prophase, Prometaphase, Metaphase, Anaphase, Telophase. Die ersten Beschreibungen der Phasen der Mitose und die Festlegung ihrer Abfolge erfolgten in den 70er-80er Jahren des 19. Jahrhunderts. In den späten 1870er Jahren prägte der deutsche Histologe Walter Flemming den Begriff „Mitose“, um den Prozess der indirekten Zellteilung zu bezeichnen.
Die durchschnittliche Dauer der Mitose beträgt 1-2 Stunden. Die Mitose tierischer Zellen dauert in der Regel 30 bis 60 Minuten und bei Pflanzen 2 bis 3 Stunden. Im Laufe von 70 Jahren kommt es im menschlichen Körper insgesamt zu etwa 10 14 Zellteilungen.
Die ersten unvollständigen Beschreibungen über das Verhalten und die Veränderungen von Kernen in sich teilenden Zellen finden sich in den Arbeiten von Wissenschaftlern aus den frühen 1870er Jahren.
Im Werk des russischen Botanikers Russov aus dem Jahr 1872 werden Metaphase- und Anaphaseplatten, die aus einzelnen Chromosomen bestehen, anschaulich beschrieben und dargestellt.
Ein Jahr später wurde der deutsche Zoologe G.A. Noch klarer und konsequenter, wenn auch natürlich nicht ganz vollständig, beschrieb Schneider die mitotische Teilung am Beispiel des Zerkleinerns von Eiern der Rektumturbellaria Mesostomus. Seine Arbeit wird im Wesentlichen beschrieben und illustriert in richtige Reihenfolge Hauptphasen der Mitose: Prophase, Metaphase, Anaphase (früh und spät). Im Jahr 1874 gründete der Moskauer Botaniker I.D. Auch bei den Sporen von Bärenmoosen und Schachtelhalmen beobachtete Tschistjakow einzelne Phasen der Zellteilung. Trotz der ersten Erfolge gelang es weder Russov noch Schneider noch Chistyakov, eine klare und konsistente Beschreibung der mitotischen Teilung zu geben.
Im Jahr 1875 wurden Werke veröffentlicht, die detailliertere Beschreibungen von Mitosen enthielten. O. Büchli beschrieb die zytologischen Muster in den zerquetschenden Eiern von Spulwürmern und Weichtieren sowie in den Spermatogenesezellen von Insekten.
E. Strassburger untersuchte die mitotische Teilung in den Zellen der Grünalge Spirogyra, in den Mutterzellen von Zwiebelpollen und in den Muttersporenzellen des Mooses. Unter Bezugnahme auf die Arbeiten von O. Büchli und basierend auf seinen eigenen Forschungen machte E. Strassburger auf die Einheitlichkeit der Zellteilungsprozesse in pflanzlichen und tierischen Zellen aufmerksam.
Ende 1878 - Anfang 1879 detaillierte Arbeit Schleicher und W. Flemming. In seinem Werk von 1879 schlug Schleicher den Begriff „Karyokinese“ vor, um die komplexen Prozesse der Zellteilung zu bezeichnen, die Bewegung implizieren Komponenten Kerne. Walter Flemming prägte als Erster den Begriff „Mitose“ für die indirekte Zellteilung, der sich später allgemein durchsetzte. Flemming ist auch für die endgültige Formulierung der Definition der Mitose als eines zyklischen Prozesses verantwortlich, der mit der Teilung der Chromosomen zwischen Tochterzellen endet.
Im Jahr 1880 O.V. Baranetsky begründete die Spiralstruktur der Chromosomen. Im Zuge weiterer Forschungen wurden Ideen zur Spiralisierung und Despiralisierung von Chromosomen während des Mitosezyklus entwickelt.
Zu Beginn des 20. Jahrhunderts wurden Chromosomen als Träger identifiziert erbliche Informationen, was später eine Erklärung lieferte biologische Rolle Mitose, die aus der Bildung genetisch identischer Tochterzellen besteht.
In den 1970er Jahren begann die Entschlüsselung und detaillierte Untersuchung der Regulatoren der mitotischen Teilung dank einer Reihe von Experimenten zur Zellfusion in verschiedenen Stadien des Zellzyklus. Wenn in diesen Experimenten eine Zelle in der M-Phase mit einer Zelle in einem der Interphase-Stadien (G 1, S oder G 2) kombiniert wurde, gingen die Interphase-Zellen in den mitotischen Zustand über (die Chromosomenkondensation begann und die Kernmembran zerfiel).
Als Ergebnis wurde der Schluss gezogen, dass die Zytoplasmazelle einen Faktor (oder Faktoren) enthält, der die Mitose stimuliert, oder mit anderen Worten einen M-stimulierenden Faktor (MSF, vom englischen M-Phase-Promoting Factor, MPF).
Erstmals wurde der „Mitose-Stimulationsfaktor“ in reifen, unbefruchteten Krallenfrosch-Eiern in der M-Phase des Zellzyklus entdeckt. Das in die Eizelle injizierte Zytoplasma einer solchen Eizelle führte zu einem vorzeitigen Übergang in die M-Phase und zum Beginn der Reifung der Eizelle (ursprünglich stand die Abkürzung MPF für Maturation Promoting Factor, was übersetzt „Reifungsförderung“ bedeutet). Faktor"). Im Zuge weiterer Experimente wurde gleichzeitig eine universelle Bedeutung festgestellt hochgradig Konservatismus des „Mitose-Stimulationsfaktors“: Extrakte, die aus mitotischen Zellen sehr unterschiedlicher Organismen hergestellt wurden, überführten sie beim Einbringen in Krallenfrosch-Oozyten in die M-Phase.
Nachfolgende Studien ergaben, dass der Mitose-stimulierende Faktor ein heterodimerer Komplex ist, der aus einem Cyclin-Protein und einer Cyclin-abhängigen Proteinkinase besteht. Cyclin ist regulatorisches Protein und kommt in allen Eukaryoten vor. Seine Konzentration steigt während des Zellzyklus periodisch an und erreicht in der Metaphase der Mitose ein Maximum. Mit Beginn der Anaphase wird ein starker Rückgang der Cyclinkonzentration beobachtet, da dieser mithilfe komplexer proteolytischer Proteinkomplexe – Proteasomen – abgebaut wird. Cyclin-abhängige Proteinkinase ist ein Enzym (Phosphorylase), das Proteine modifiziert, indem es eine Phosphatgruppe von ATP auf die Aminosäuren Serin und Threonin überträgt. Mit der Aufklärung der Rolle und Struktur des Hauptregulators der mitotischen Teilung begann die bis heute andauernde Erforschung der feinen Regulationsmechanismen der Mitose.
Die Entwicklung einer einheitlichen Typologie und Klassifizierung von Mitosen wird durch eine ganze Reihe von Merkmalen erschwert, die in verschiedenen Kombinationen zu Vielfalt und Heterogenität in den Mustern der mitotischen Teilung führen. Gleichzeitig sind bestimmte Klassifizierungsoptionen, die für einige Taxa entwickelt wurden, für andere inakzeptabel, da sie die Besonderheiten ihrer Mitosen nicht berücksichtigen. Beispielsweise erweisen sich bestimmte Varianten der Klassifizierung von Mitosen, die für tierische oder pflanzliche Organismen charakteristisch sind, für Algen als inakzeptabel.
Eines der Schlüsselmerkmale, die verschiedenen Typologien und Klassifikationen der mitotischen Teilung zugrunde liegen, ist das Verhalten der Kernhülle. Erfolgt die Bildung der Spindel und die mitotische Teilung selbst im Kern ohne Zerstörung der Kernmembran, spricht man von dieser Art der Mitose geschlossen. Mitose mit Zerfall der Kernmembran wird dementsprechend als offen bezeichnet, Mitose mit Zerfall der Kernmembran nur an den Spindelpolen unter Bildung von „Polarfenstern“ als halbgeschlossen.
Ein weiteres charakteristisches Merkmal ist die Art der Symmetrie der mitotischen Spindel. Bei der Pleuromitose ist die Spindel beidseitig symmetrisch oder asymmetrisch und besteht in der Regel aus zwei in Metaphase-Anaphase in einem Winkel zueinander stehenden Halbspindeln. Die Kategorie der Orthomitosen zeichnet sich durch eine bipolare Symmetrie der Spindel aus, und in der Metaphase wird häufig eine erkennbare Äquatorialplatte beobachtet.
Im Rahmen der angegebenen Symptome ist die typische offene Orthomitose am zahlreichsten, im Folgenden werden beispielsweise die Prinzipien und Stadien der mitotischen Teilung erörtert. Diese Art der Mitose ist charakteristisch für Tiere, höhere Pflanzen und einige Protozoen.
Prophase beginnt mit der Verdichtung von Chromosomen, die unter dem Lichtmikroskop als fadenförmige Strukturen sichtbar werden. Jedes Chromosom besteht aus zwei parallelen Schwesterchromatiden, die am Zentromer verbunden sind. Der Nukleolus und die Kernhülle verschwinden am Ende der Phase (letztere zerfallen in Membranvesikel, ähnlich den Elementen des EPS, und der Porenkomplex und die Lamina dissoziieren in Untereinheiten). Karyoplasma ist mit Zytoplasma vermischt.
Zentriolen wandern zu entgegengesetzten Polen der Zelle und bilden die Filamente der mitotischen Spindel (Achromatin). Im Bereich des Zentromers bilden sich spezielle Proteinkomplexe – Kinetochoren, an denen einige Mikrotubuli der Spindel befestigt sind (Kinetochor-Mikrotubuli); Es wurde gezeigt, dass Kinetochoren selbst in der Lage sind, den Aufbau von Mikrotubuli zu induzieren und daher als Organisationszentren für Mikrotubuli dienen können. Die übrigen Mikrotubuli der Spindel werden Pol-Mikrotubuli genannt, da sie sich von einem Pol der Zelle zum anderen erstrecken; Die außerhalb der Spindel liegenden Mikrotubuli, die radial von den Zellzentren zum Plasmalemma divergieren, werden Astral- oder Mikrotubuli (Fäden) der Strahlung genannt.
Die Metaphase entspricht dem maximalen Kondensationsgrad der Chromosomen, die sich im Äquatorbereich der mitotischen Spindel aneinanderreihen und ein Bild der Äquatorplatte (Metaphase) (Seitenansicht) oder des Muttersterns (Ansicht von den Polen) bilden. Chromosomen bewegen sich zur Äquatorialebene und werden dort durch die ausgeglichene Spannung der Kinetochor-Mikrotubuli gehalten. Am Ende dieser Phase sind die Schwesterchromatiden durch eine Spalte getrennt, verbleiben aber im Zentromerbereich.
Die Anaphase beginnt mit der synchronen Aufspaltung aller Chromosomen in Schwesterchromatiden (im Zentromerbereich) und der Bewegung der Tochterchromosomen zu den gegenüberliegenden Polen der Zelle, die entlang der Spindelmikrotubuli mit einer Geschwindigkeit von 0,2–0,5 μm/min erfolgt. Das Signal für den Beginn der Anaphase beinhaltet einen starken (um eine Größenordnung) Anstieg der Konzentration von Calciumkationen im Hyaloplasma, die von Membranvesikeln abgesondert werden, die an den Spindelpolen Cluster bilden. Der Mechanismus der Chromosomenbewegung in der Anaphase ist nicht vollständig aufgeklärt, es wurde jedoch festgestellt, dass es in der Spindelregion neben Aktin auch Proteine wie Myosin und Dynein sowie eine Reihe regulatorischer Proteine gibt. Einigen Beobachtungen zufolge wird es durch die Verkürzung (Demontage) von Mikrotubuli verursacht, die an Kinetochoren befestigt sind. Die Anaphase ist durch eine Verlängerung der mitotischen Spindel aufgrund einer gewissen Divergenz der Zellpole gekennzeichnet. Es endet mit der Ansammlung zweier identischer Chromosomensätze an den Polen der Zelle, die Bilder von Sternen bilden (Stadium der Tochtersterne). Am Ende der Anaphase beginnt sich aufgrund der Kontraktion der um den Zellumfang konzentrierten Aktin-Mikrofilamente (kontraktiler Ring) eine zelluläre Verengung zu bilden, die, wenn sie sich vertieft, in der nächsten Phase zur Zytotomie führt.
Die Telophase ist das letzte Stadium der Mitose, in dem die Kerne der Tochterzellen rekonstruiert werden und ihre Trennung abgeschlossen ist. Um die kondensierten Chromosomen von Tochterzellen aus Membranvesikeln (nach anderen Quellen aus dem EPS) wird das Karyolemma wiederhergestellt, mit dem die sich bildende Lamina verbunden ist, und es entstehen wieder Nukleolen, die aus Abschnitten der entsprechenden Chromosomen gebildet werden. Die Zellkerne vergrößern sich allmählich, und die Chromosomen despiralieren zunehmend und verschwinden und werden durch das Chromatinmuster des Interphasekerns ersetzt. Gleichzeitig vertieft sich die Zellverengung und die Zellen bleiben für einige Zeit durch eine sich verjüngende Zytoplasmabrücke verbunden, die ein Bündel von Mikrotubuli (Mediankörper) enthält. Die weitere Ligation des Zytoplasmas endet mit der Bildung von zwei Tochterzellen. In der Telophase werden Organellen zwischen den Tochterzellen verteilt; Die Einheitlichkeit dieses Prozesses wird durch die Tatsache erleichtert, dass einige Organellen recht zahlreich sind (z. B. Mitochondrien), während andere (wie das ER und der Golgi-Komplex) während der Mitose in kleine Fragmente und Vesikel zerfallen.
Atypische Mitosen treten auf, wenn der Mitoseapparat beschädigt ist, und sind durch eine ungleichmäßige Verteilung des genetischen Materials zwischen den Zellen gekennzeichnet – Aneuploidie (aus dem Griechischen an – nicht, eu – richtig, ploon – Falte); In vielen Fällen findet keine Zytotomie statt, was zur Bildung von Riesenzellen führt. Atypische Mitosen sind charakteristisch für bösartige Tumoren und bestrahltes Gewebe. Je höher ihre Häufigkeit und je größer der Grad der Aneuploidie, desto bösartiger ist der Tumor. Eine Störung der normalen mitotischen Zellteilung kann durch Chromosomenanomalien verursacht werden, die als Chromosomenaberrationen (von lateinisch Aberratio – Abweichung) bezeichnet werden. Zu den Varianten chromosomaler Aberrationen gehören die Adhäsion von Chromosomen, deren Bruch in Fragmente, der Verlust eines Abschnitts, der Austausch von Fragmenten, die Verdoppelung einzelner Chromosomenabschnitte usw. Chromosomenaberrationen können spontan auftreten, entwickeln sich jedoch häufiger als Folge der Einwirkung von Mutagene und ionisierende Strahlung auf Zellen.
Karyotypisierung – eine diagnostische Studie zur Beurteilung des Karyotyps (Chromosomensatzes), die durch Untersuchung der Chromosomen in der Metaphaseplatte durchgeführt wird. Zur Karyotypisierung wird eine Zellkultur gewonnen, in die Colchicin eingebracht wird, eine Substanz, die die Bildung der Mitosespindel blockiert. Aus solchen Zellen werden Chromosomen extrahiert, die dann gefärbt und identifiziert werden. Ein normaler menschlicher Karyotyp wird durch 46 Chromosomen repräsentiert – 22 Autosomenpaare und zwei Geschlechtschromosomen (XY bei Männern und XX bei Frauen). Mit der Karyotypisierung können Sie eine Reihe von damit verbundenen Krankheiten diagnostizieren Chromosomenanomalien, insbesondere Down-Syndrom (Trisomie 21. Chromosom), Edwards-Syndrom (Trisomie 18. Chromosom), Patau-Syndrom (Trisomie 13. Chromosom) sowie eine Reihe von Syndromen, die mit Anomalien der Geschlechtschromosomen verbunden sind – Klinefelter-Syndrom (Genotyp - XXY), Turner (Genotyp - XO) und andere.
Es wird angenommen, dass sich der komplexe mitotische Prozess höherer Organismen schrittweise aus den Teilungsmechanismen von Prokaryoten entwickelte. Diese Annahme wird durch die Tatsache bestätigt, dass Prokaryoten etwa eine Milliarde Jahre früher auftauchten als die ersten Eukaryoten. Darüber hinaus sind ähnliche Proteine an der Mitose von Eukaryoten und der binären Spaltung von Prokaryoten beteiligt.
Mögliche Zwischenstadien zwischen binärer Spaltung und Mitose lassen sich bei einzelligen Eukaryoten nachweisen, bei denen die Kernmembran bei der Teilung nicht zerstört wird. Bei den meisten anderen Eukaryoten, einschließlich Pflanzen und Tieren, wird die Spindel außerhalb des Kerns gebildet und die Kernhülle wird während der Mitose zerstört. Obwohl die Mitose bei einzelligen Eukaryoten noch nicht gut verstanden ist, kann man davon ausgehen, dass sie aus der binären Spaltung entstand und schließlich das in mehrzelligen Organismen anzutreffende Komplexitätsniveau erreichte.
Bei vielen einfachen Eukaryoten blieb die Mitose ebenfalls ein Prozess, der mit der Membran verbunden war, jedoch nicht mit Plasma, sondern mit Kern.
Die wichtigsten Regulationsmechanismen der Mitose sind die Prozesse der Phosphorylierung und Proteolyse.
Reversible Phosphorylierungs- und Dephosphorylierungsreaktionen ermöglichen reversible Ereignisse der Mitose, wie z. B. den Zusammenbau/Zerfall der Spindel oder den Zerfall/die Reparatur der Kernhülle. Die Proteolyse liegt den irreversiblen Ereignissen der Mitose zugrunde, beispielsweise der Trennung von Schwesterchromatiden in der Anaphase oder der Zerstörung mitotischer Zykline in späten Stadien der Mitose.
Die Teilung aller eukaryotischen Zellen ist mit der Bildung eines speziellen Zellteilungsapparates verbunden.
Zytoskelettstrukturen wird häufig eine aktive Rolle bei der mitotischen Zellteilung zugeschrieben. Universell für tierische und pflanzliche Zellen ist die bipolare mitotische Spindel, die aus Mikrotubuli und zugehörigen Proteinen besteht. Die Spindel sorgt für eine streng identische Verteilung der Chromosomen zwischen den Teilungspolen, in deren Bereich sich in der Telophase die Kerne von Tochterzellen bilden.
Der Prozess der Mitose gewährleistet eine streng gleichmäßige Verteilung der Chromosomen zwischen zwei Tochterkernen, sodass in einem vielzelligen Organismus alle Zellen genau die gleichen (in Anzahl und Beschaffenheit) Chromosomensätze aufweisen.
Chromosomen enthalten genetische Informationen, die in der DNA kodiert sind, und daher stellt der regelmäßige, geordnete mitotische Prozess auch sicher, dass alle Informationen vollständig an jeden der Tochterkerne übertragen werden; Dadurch verfügt jede Zelle über alle genetischen Informationen, die zur Entwicklung aller Eigenschaften des Organismus erforderlich sind. In diesem Zusammenhang wird deutlich, warum sich aus einer Zelle einer voll differenzierten erwachsenen Pflanze unter geeigneten Bedingungen eine ganze Pflanze entwickeln kann. Wir haben die Mitose in einer diploiden Zelle beschrieben, aber dieser Prozess läuft in ähnlicher Weise in haploiden Zellen ab, beispielsweise in den Zellen der gametophytischen Generation von Pflanzen.
Durch die Teilung somatischer Zellen wächst der Welpe und nimmt an Größe zu Mitose. Mitose ist die indirekte Teilung einer somatischen Zelle, bei der komplexe Veränderungen in ihrem Zellkern und Zytoplasma auftreten. Nach der Befruchtung (Ovogamie) einer Eizelle durch ein Spermium (Fusion oder Kopulation von Gameten), a Zygote(Oozyste) ist ein neuer Organismus, der nur aus einer Zelle besteht. Der Prozess des Wachstums und der Entwicklung eines neuen Organismus beginnt ab dem Moment der ersten mitotischen Teilung dieser Zelle (mütterlicherseits), wenn daraus zwei ihr völlig ähnliche Tochterzellen (genauer gesagt Schwesterzellen) entstehen, und dauert bis zum Tod
Abb. 1 Zellstruktur
Während des Mitoseprozesses kommt es zu:
1- Verdoppelung der Chromosomensubstanz;
2- Veränderung des physikalischen Zustands und der chemischen Organisation der Chromosomen;
3- Divergenz der Schwesterchromosomen zu den Zellpolen;
4- anschließende Teilung des Zytoplasmas und vollständige Wiederherstellung von zwei Kernen in neuen Zellen.
Mitose umfasst den Lebenszyklus nuklearer Gene: Duplikation, Verteilung und Funktion. Der Zeitraum zwischen Zellteilungen wird genannt Interphase, in deren Verlauf aktive Lebensprozesse und die Vorbereitung auf die nächste Teilung stattfinden. Der gesamte Zyklus der Veränderungen in der Zelle
von einer Abteilung zur anderen heißt Mitotischer Zyklus. Letztere besteht aus zwei Hauptperioden – der Interphase und der Mitose selbst.
Durch die Mitose entstehen aus einer Zelle zwei Zellen mit identischen Chromosomen. Somit gewährleistet die Mitose die Kontinuität und Konstanz der Anzahl und Menge, also die qualitative Spezifität der Chromosomen in aufeinanderfolgenden Generationen sich teilender Zellen (siehe Abb. 2).
In der Interphase, dem Zeitraum zwischen zwei aufeinanderfolgenden Zellteilungen, erfolgt die Replikation (Autoduplikation oder Selbstduplikation) der DNA im Zellkern und damit die Anzahl der Chromosomen in der Zelle (d. h. die Bildung von Schwesterchromatiden, die durch das Zentromer zusammengehalten werden). , ein Körper, der als mechanisches Zentrum des Chromosoms fungiert) sowie dessen Despiralisierung.
In der Metaphase, der zentralen Phase der Kernteilung, verwandelt sich ein Chromosom, das aus zwei Chromatiden besteht, in zwei Tochterchromosomen.
Reis. 2. Mitose
 1 - drei Chromosomenpaare; 2 - Replikation von Chromosomen unter Bildung von Schwesterchromatiden, die am Zentromer verbunden sind; 3 - an den Polen der Kernhülle sind Zentrosomen mit Asteralstrahlen sichtbar, die in den zentromeren Bereich der Chromatiden gehen und entlang des Äquators ausgerichtet sind, um die Zentromere der Schwesterchromatiden zu trennen und sie zu verschiedenen Polen zu bewegen; 4 - Despiralisierung der Chromosomen, Wiederherstellung der Kernmembran und Bildung eines Zellseptums mit der Bildung von zwei mit der Mutter identischen Tochterzellen mit genau der gleichen Anzahl an Chromosomen wie in dieser
1 - drei Chromosomenpaare; 2 - Replikation von Chromosomen unter Bildung von Schwesterchromatiden, die am Zentromer verbunden sind; 3 - an den Polen der Kernhülle sind Zentrosomen mit Asteralstrahlen sichtbar, die in den zentromeren Bereich der Chromatiden gehen und entlang des Äquators ausgerichtet sind, um die Zentromere der Schwesterchromatiden zu trennen und sie zu verschiedenen Polen zu bewegen; 4 - Despiralisierung der Chromosomen, Wiederherstellung der Kernmembran und Bildung eines Zellseptums mit der Bildung von zwei mit der Mutter identischen Tochterzellen mit genau der gleichen Anzahl an Chromosomen wie in dieser
In der Anaphase kommt es zur Teilung und Divergenz der Tochterchromosomen zu den Zellpolen, das heißt, ihre richtige Anzahl wird wiederhergestellt. In der Telophase, dem Endstadium der Zellteilung, nehmen die Chromosomen das gleiche Aussehen wie vor Beginn der Teilung an und die DNA-Menge in jedem Tochterkern ist im Vergleich zu früheren Stadien halbiert. Somit enthalten beide Tochterzellen gleiche Mengen an Zytoplasma und identische Chromosomensätze und sind bereit für die Mitose.
Nicht alle Körperzellen des Körpers teilen sich ständig. Im Verlauf der Embryonalentwicklung kommt es zur Differenzierung von Organen und Geweben, die sich entlang ihres eigenen spezifischen, genetisch festgelegten Weges entwickeln. Daher verwandeln sich einige Zellen in Gehirnzellen, andere in Blutzellen usw. Darüber hinaus teilen sich einige von ihnen ständig, während andere erst in einem bestimmten Entwicklungsstadium oder bei Bedarf beispielsweise für die Regeneration zuständig sind
(restaurative) Prozesse.