Histologie generală. Embriologie generală şi comparativă Metoda de gastrulare la păsări
Citeste si
În om implantare submersibil sau interstițial. Într-o zi, blastocistul se scufundă în endometru aproape jumătate, iar după 40 de ore - complet. Defectul format în membrana mucoasă a uterului este restabilit în 5 zile. În mod convențional, implantarea constă din două faze - aderența (aderența) blastocistului la mucoasa uterină și imersarea (invazia) blastocistului în profunzimea mucoasei.
În a 6-a zi embriogeneza blastocistul este atașat de epiteliul endometrial (de obicei prin polul embrionar în regiunea peretelui posterior sau ventral la unghiul uterin). În aderența blastocistului, un rol important joacă substanțe din grupa integrinei, produse de celulele epiteliale ale mucoasei uterine. În mod normal, la femei, între 19-24 de zile ale ciclului menstrual – adică la cel mai optim moment pentru interacțiunea cu blastocistul, există o expresie activă a genei integrinei în celulele epiteliale ale mucoasei uterine.
gastrulatie
La fătul uman gastrulația este inițiată la sfârșitul primei săptămâni de dezvoltare, imediat după scindarea și îndepărtarea zonei transparente, când embrionul începe să se scufunde în peretele uterin.
gastrulatie prima faza continuă pe parcursul celei de-a 2-a săptămâni de dezvoltare. Materialul masei celulare interioare este împărțit prin delaminare în două foi - epiblast (partea superioară) și hipoblast (partea inferioară). Celulele hipoblastice sunt cubice mici, citoplasma lor are un aspect spumant, formează un strat subțire sub epiblast și mărginesc cavitatea blastocistului. Celulele epiblastice sunt mai înalte și arată ca epiteliul prismatic pseudostratificat.
Celulele epiblastice sunt separate prin intermediul unor cavități mici, la confluența cărora se formează ulterior cavitatea amniotică. În partea inferioară a cavității amniotice, rămâne un mic grup de celule epiblastice - materialul viitorului embrion (embrioblast) și organele extraembrionare.
Faza a 2-a de gastrulatie apare în a 3-a săptămână de dezvoltare, se realizează prin imigrare și se termină cu formarea a trei straturi germinale - ectoderm, mezoderm, endoderm. În viitor, țesuturile embrionului și ale organelor extraembrionare vor apărea din materialul acestor foi. Evenimentele care au loc în faza a 2-a a gastrulației seamănă cu cele din embrionul de pui și reprezentanții mamiferelor.
Iniţiere imigrația este asociată cu proliferarea celulară activă și neuniformă în diferite părți ale epiblastului, ceea ce duce la formarea de fluxuri celulare și formarea de structuri pe suprafața epiblastului, cunoscute sub numele de dâra primară și nodul lui Hensen. Discul germinal rotunjit si plat in faza a 2-a de gastrulatie se transforma intr-unul alungit cu craniul expandat si capete caudale mai inguste.
Semnificație morfogenetică deosebităîn embriogeneza timpurie, aparține procesului capului (notocorda). Procesul capului crește în direcția craniană între epi- și hipoblast până ajunge la placa precordală, care este un „indicator” al viitoarei guri a embrionului. Procesul capului dă naștere dezvoltării notocordului embrionului - un fel de tijă celulară care determină axa primară a embrionului și îi conferă „rigiditate”.
Coardă formează axa scheletului embrionului uman și stă la baza dezvoltării oaselor scheletului axial (coloana vertebrală, coaste, stern, craniu). Coloana vertebrală se va forma în jurul coardei în viitor. Notocorda are un efect inductor important asupra diferențierii ectodermului adiacent acestuia de sus în placa neurală și apoi în tubul neural, din care se vor dezvolta creierul și măduva spinării. Până la sfârșitul celei de-a treia săptămâni de embriogeneză, notocordul este aproape complet format și se extinde de la membrana orofaringiană până la capătul caudal al embrionului.
EMBRIOLOGIE. Capitolul 21. BAZELE EMBRIOLOGIEI UMANE
EMBRIOLOGIE. Capitolul 21. BAZELE EMBRIOLOGIEI UMANE
Embriologie (din greacă. embrionară- embrion, logos- doctrină) - știința legilor dezvoltării embrionilor.
Embriologia medicală studiază modelele de dezvoltare ale embrionului uman. Se acordă o atenție deosebită surselor embrionare și proceselor regulate de dezvoltare a țesuturilor, caracteristicilor metabolice și funcționale ale sistemului mamă-placenta-făt și perioadelor critice ale dezvoltării umane. Toate acestea sunt de mare importanță pentru practica medicală.
Cunoașterea embriologiei umane este necesară pentru toți medicii, în special pentru cei care lucrează în domeniul obstetrică și pediatrie. Acest lucru ajută la diagnosticarea tulburărilor din sistemul mamă-făt, identificând cauzele deformărilor și bolilor la copii după naștere.
În prezent, cunoștințele despre embriologia umană sunt folosite pentru a descoperi și elimina cauzele infertilității, transplantul de organe fetale și dezvoltarea și utilizarea contraceptivelor. În special, problemele culturii ouălor, fertilizarea in vitro și implantarea embrionilor în uter au devenit actuale.
Procesul de dezvoltare embrionară umană este rezultatul unei evoluții îndelungate și reflectă într-o anumită măsură trăsăturile dezvoltării altor reprezentanți ai lumii animale. Prin urmare, unele dintre etapele timpurii ale dezvoltării umane sunt foarte asemănătoare cu stadiile similare în embriogeneza cordatelor organizate inferioare.
Embriogeneza umană este o parte a ontogenezei sale, incluzând următoarele etape principale: I - fertilizarea și formarea zigoților; II - zdrobirea și formarea blastulei (blastocist); III - gastrulația - formarea straturilor germinale și a unui complex de organe axiale; IV - histogeneza si organogeneza organelor germinale si extraembrionare; V - sistemogeneza.
Embriogeneza este strâns legată de progeneza și perioada postembrionară timpurie. Astfel, dezvoltarea tesuturilor incepe in perioada embrionara (histogeneza embrionara) si continua dupa nasterea unui copil (histogeneza postembrionara).
21.1. PROGENEZĂ
Aceasta este perioada de dezvoltare și maturare a celulelor germinale - ovule și spermatozoizi. Ca urmare a progenezei, un set haploid de cromozomi apare în celulele germinale mature, se formează structuri care oferă capacitatea de a fertiliza și de a dezvolta un nou organism. Procesul de dezvoltare a celulelor germinale este analizat în detaliu în capitolele privind sistemele reproductive masculine și feminine (vezi capitolul 20).
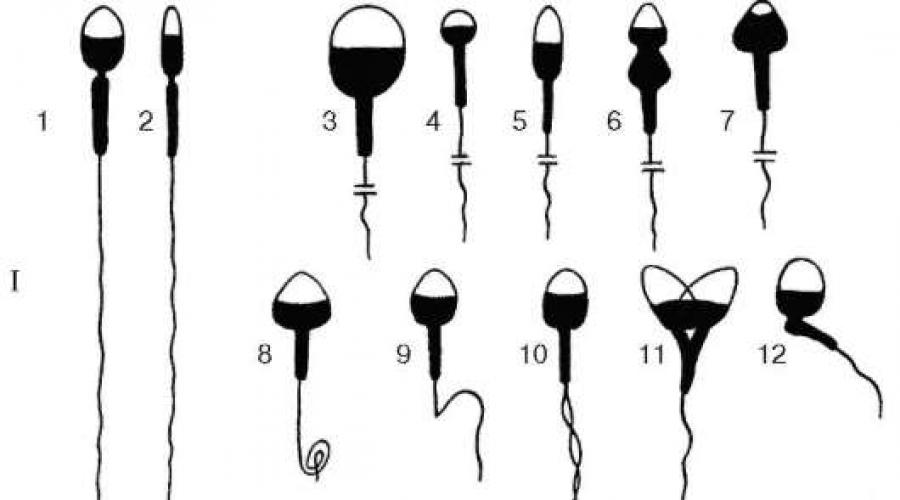
Orez. 21.1. Structura celulei germinale masculine:
I - cap; II - coada. 1 - receptor;
2 - acrozom; 3 - „caz”; 4 - centriol proximal; 5 - mitocondrie; 6 - strat de fibrile elastice; 7 - axonă; 8 - inel terminal; 9 - fibrile circulare
Principalele caracteristici ale celulelor germinale umane mature
celule reproductive masculine
Spermatozoizii umani sunt produși pe toată perioada sexuală activă în cantități mari. Descriere detaliata spermatogeneza - vezi capitolul 20.
Motilitatea spermatozoizilor se datorează prezenței flagelilor. Viteza de mișcare a spermatozoizilor la om este de 30-50 microni/s. Mișcarea intenționată este facilitată de chemotaxie (mișcare către sau departe de un stimul chimic) și reotaxie (mișcare împotriva fluxului de fluid). La 30-60 de minute după actul sexual, spermatozoizii se găsesc în cavitatea uterină, iar după 1,5-2 ore - în partea distală (ampulară) a trompei, unde se întâlnesc cu ovulul și fecundarea. Spermatozoizii își păstrează capacitatea de fertilizare până la 2 zile.
Structura. Celulele sexuale masculine umane - spermatozoizi, sau sperma-mii, aproximativ 70 de microni lungime, au cap și coadă (Fig. 21.1). Membrana plasmatică a spermatozoidului din zona capului conține un receptor, prin care are loc interacțiunea cu ovulul.
Capul spermatozoidului include un nucleu mic dens cu un set haploid de cromozomi. Jumătatea anterioară a nucleului este acoperită cu un sac plat caz sperma. În ea se află acrozom(din greaca. asron- sus, soma- corp). Acrozomul conține un set de enzime, printre care un loc important revine hialuronidazei și proteazelor, care sunt capabile să dizolve membranele care acoperă ovulul în timpul fertilizării. Cazul și acrozomul sunt derivați ai complexului Golgi.

Orez. 21.2. Compoziția celulară a ejaculatului uman este normală:
I - celule sexuale masculine: A - mature (după L.F. Kurilo și alții); B - imatur;
II - celule somatice. 1, 2 - spermatozoid tipic (1 - fata intreaga, 2 - profil); 3-12 - cele mai frecvente forme de atipie a spermatozoizilor; 3 - cap macro; 4 - microcap; 5 - cap alungit; 6-7 - anomalie în forma capului și a acrozomului; 8-9 - anomalie a flagelului; 10 - spermatozoizi biflagelati; 11 - capete fuzionate (sperma cu două capete); 12 - anomalie a gâtului spermatozoizilor; 13-18 - celule sexuale masculine imature; 13-15 - spermatocite primare în profaza primei diviziuni a meiozei - proleptoten, pachiten, respectiv diploten; 16 - spermatocitul primar în metafaza meiozei; 17 - spermatide tipice (A- din timp; b- târziu); 18 - spermatid binuclear atipic; 19 - celule epiteliale; 20-22 - leucocite
Nucleul spermatozoidului uman conține 23 de cromozomi, dintre care unul sexual (X sau Y), restul sunt autozomi. 50% dintre spermatozoizi conțin cromozomul X, 50% - cromozomul Y. Masa cromozomului X este ceva mai mare decât masa cromozomului Y, prin urmare, aparent, spermatozoizii care conțin cromozomul X sunt mai puțin mobili decât spermatozoizii care conțin cromozomul Y.
În spatele capului există o îngustare inelară, care trece în secțiunea cozii.
secțiunea de coadă (flagel) Spermatozoidul este format dintr-o parte de legătură, intermediară, principală și terminală. În partea de legătură (pars conjungens), sau gât (colul uterin) se situează centriolii - proximali, adiacente nucleului, iar resturile centriolului distal, coloane striate. Aici începe filetul axial (axonem), continuând în părțile intermediare, principale și terminale.
Parte intermediară (pars intermedia) contine 2 microtubuli centrali si 9 perechi de microtubuli periferici inconjurati de mitocondrii dispuse spiralat (teaca mitocondriala - vagin mitocondrial). Proeminențele pereche, sau „mânere”, constând dintr-o altă proteină, dineina, care are activitate ATP-azei, pleacă de la microtubuli (vezi capitolul 4). Dineina descompune ATP produs de mitocondrii și transformă energia chimică în energie mecanică, datorită căreia se realizează mișcarea spermatozoizilor. În cazul unei absențe determinate genetic a dineinei, spermatozoizii sunt imobilizați (una dintre formele sterilității masculine).
Printre factorii care afectează viteza de mișcare a spermatozoizilor, temperatura, pH-ul mediului etc. sunt de mare importanță.
parte principală (pars principali) Structura cozii seamănă cu un ciliu cu un set caracteristic de microtubuli în axonem (9 × 2) + 2, înconjurat de fibrile orientate circular care conferă elasticitate și o plasmălemă.
Terminal, sau partea finală sperma (pars terminalis) conţine un axonem care se termină în microtubuli deconectaţi şi o scădere treptată a numărului acestora.
Mișcările cozii sunt de tip bici, ceea ce se datorează contracției succesive a microtubulilor de la prima la a noua pereche (prima este considerată o pereche de microtubuli, care se află într-un plan paralel cu cei doi centrali).
În practica clinică, în studiul spermatozoizilor se numără diverse forme de spermatozoizi, numărându-se procentul acestora (spermograma).
Potrivit Organizației Mondiale a Sănătății (OMS), următorii indicatori sunt caracteristici normale ale spermei umane: concentrația spermatozoizilor - 20-200 milioane / ml, conținutul în ejaculat este mai mult de 60% din formele normale. Alături de acestea din urmă, spermatozoidul uman conține întotdeauna și anormale - biflagelate, cu dimensiunile capului defecte (macro și microforme), cu cap amorf, cu fuzionate.
capete, forme imature (cu resturi de citoplasmă în gât și coadă), cu defecte de flagel.
În ejaculatul bărbaților sănătoși predomină spermatozoizii tipici (Fig. 21.2). Numărul diferitelor tipuri de spermatozoizi atipici nu trebuie să depășească 30%. În plus, există forme imature de celule germinale - spermatide, spermatocite (până la 2%), precum și celule somatice - epiteliocite, leucocite.
Dintre spermatozoizii din ejaculat, celulele vii ar trebui să fie de 75% sau mai mult și active mobile - 50% sau mai mult. Parametrii normativi stabiliți sunt necesari pentru evaluarea abaterilor de la normă în diferite forme de infertilitate masculină.
Într-un mediu acid, spermatozoizii își pierd rapid capacitatea de mișcare și fertilizare.
celule sexuale feminine
ouă, sau ovocite(din lat. ovul- ou), se coace într-o cantitate nemăsurat mai mică decât spermatozoizii. La o femeie în timpul ciclului sexual (24-28 de zile), de regulă, un ou se maturizează. Astfel, în perioada fertilă se formează aproximativ 400 de ouă.
Eliberarea unui ovocit dintr-un ovar se numește ovulație (vezi capitolul 20). Ovocitul eliberat din ovar este înconjurat de o coroană de celule foliculare, al căror număr ajunge la 3-4 mii. formă sferică, mai mare decât cel al spermatozoizilor, volumul citoplasmei nu are capacitatea de a se deplasa independent.
Clasificarea ovocitelor se bazează pe semne de prezență, cantitate și distribuție. gălbenuș (lecithos), care este o incluziune proteino-lipidică în citoplasmă, folosită pentru a hrăni embrionul. Distinge fără gălbenuș(alecital), mic-gălbenuș(oligolecital), galbenus mediu(mesolecital), gălbenuș multiplu ouă (polilecitale). Ouăle mici de gălbenuș sunt împărțite în primare (în cele non-craniene, de exemplu, lancelete) și secundare (la mamifere placentare și la oameni).
De regulă, în ouăle cu gălbenușuri mici, incluziunile de gălbenuș (granule, farfurii) sunt distribuite uniform, așa că sunt numite isolecital(gr. isos- egal). ou uman tip izolecital secundar(ca și la alte mamifere) conține nr un numar mare de granule de galbenus dispuse mai mult sau mai putin uniform.
La om, prezența unei cantități mici de gălbenuș în ou se datorează dezvoltării embrionului în corpul mamei.
Structura. Oul uman are un diametru de aproximativ 130 de microni. O zonă transparentă (lucitoare) este adiacentă lemei plasmatice (zona pellucida- Zp) și apoi un strat de celule epiteliale foliculare (Fig. 21.3).
Nucleul celulei reproducătoare feminine are un set haploid de cromozomi cu un cromozom X-sex, un nucleol bine definit și există multe complexe de pori în învelișul nucleului. În perioada de creștere a ovocitelor, în nucleu au loc procese intensive de sinteză a ARNm și ARNr.

Orez. 21.3. Structura celulei reproductive feminine:
1 - miez; 2 - plasmalema; 3 - epiteliul folicular; 4 - coroana radianta; 5 - granule corticale; 6 - incluziuni de galbenus; 7 - zona transparenta; 8 - receptor Zp3
În citoplasmă se dezvoltă aparatul de sinteză a proteinelor (reticulul endoplasmatic, ribozomi) și complexul Golgi. Numărul de mitocondrii este moderat, sunt situate în apropierea nucleului, unde există o sinteză intensivă a gălbenușului, centrul celular este absent. Complexul Golgi pe primele etape dezvoltarea este situată în apropierea nucleului, iar în procesul de maturare a oului este deplasat la periferia citoplasmei. Iată derivatele acestui complex - granule corticale (granula corticala), al cărui număr ajunge la 4000, iar dimensiunea este de 1 micron. Conțin glicozaminoglicani și diverse enzime (inclusiv cele proteolitice), participă la reacția corticală, protejând oul de polispermie.
Dintre incluziuni, ovoplasmele merită o atenție specială granule de galbenus, care conțin proteine, fosfolipide și carbohidrați. Fiecare granulă de gălbenuș este înconjurată de o membrană, are o parte centrală densă, constând din fosfovitină (fosfoproteină) și o parte periferică mai liberă, constând din lipovitelină (lipoproteină).
Zona transparentă (zona pellucida- Zp) constă din glicoproteine și glicozaminoglicani - acizi condroitin sulfuric, hialuronic și sialic. Glicoproteinele sunt reprezentate de trei fracții - Zpl, Zp2, Zp3. Fracțiile Zp2 și Zp3 formează filamente de 2–3 µm lungime și 7 nm grosime, care
interconectate folosind fracția Zpl. Fracția Zp3 este receptor spermatozoizii, iar Zp2 previne polispermia. Zona limpede conține zeci de milioane de molecule de glicoproteină Zp3, fiecare cu mai mult de 400 de resturi de aminoacizi conectate la multe ramuri de oligozaharide. Celulele epiteliale foliculare participă la formarea zonei transparente: procesele celulelor foliculare pătrund în zona transparentă, îndreptându-se spre plasmolema oului. Plasmolema oului, la rândul său, formează microvilozități situate între procesele celulelor epiteliale foliculare (vezi Fig. 21.3). Acestea din urmă îndeplinesc funcții trofice și de protecție.
21.2. Embriogeneza
Dezvoltarea intrauterină umană durează în medie 280 de zile (10 luni lunare). Se obișnuiește să se distingă trei perioade: inițială (săptămâna I), embrionară (săptămâna 2-8), fetală (de la a 9-a săptămână de dezvoltare până la nașterea unui copil). Până la sfârșitul perioadei embrionare, depunerea principalelor rudimente embrionare ale țesuturilor și organelor este finalizată.
Fertilizarea și formarea zigoților
Fertilizare (fertilizare)- fuziunea celulelor germinale masculine și feminine, în urma căreia se restabilește setul diploid de cromozomi caracteristic acestui tip de animal și apare o celulă nouă calitativ - un zigot (un ou fertilizat sau un embrion unicelular) .
La om, volumul ejaculatului - spermatozoizii erupți - este în mod normal de aproximativ 3 ml. Pentru a asigura fertilizarea, numărul total de spermatozoizi din material seminal trebuie să fie de cel puțin 150 de milioane, iar concentrația - 20-200 de milioane / ml. În tractul genital al unei femei, după copulație, numărul lor scade în direcția de la vagin la partea ampulară a trompei uterine.
În procesul de fertilizare se disting trei faze: 1) interacţiunea la distanţă şi convergenţa gameţilor; 2) interacțiunea de contact și activarea oului; 3) pătrunderea spermatozoizilor în ovul și fuziunea ulterioară - singamie.
Primă fază- interacțiunea la distanță - este asigurată de chemotaxie - un set de factori specifici care cresc probabilitatea întâlnirii cu celulele germinale. Se joacă un rol important în acest sens gamonii- chimicale produse de celulele germinale (Fig. 21.4). De exemplu, ouăle secretă peptide care ajută la atragerea spermatozoizilor.
Imediat după ejaculare, spermatozoizii nu sunt capabili să pătrundă în ovul până nu are loc capacitate - dobândirea capacității de fertilizare de către spermatozoizi sub acțiunea secretului tractului genital feminin, care durează 7 ore.În procesul de capacitare, glicoproteinele și proteinele sunt îndepărtat din plasmolema spermatozoizilor în plasma seminal acrozomului, ceea ce contribuie la reacția acrozomală.

Orez. 21.4. Interacțiunea la distanță și de contact dintre spermatozoizi și ovul: 1 - spermatozoizii și receptorii săi pe cap; 2 - separarea carbohidraților de la suprafața capului în timpul capacității; 3 - legarea receptorilor spermatozoizilor de receptorii ovulelor; 4 - Zp3 (a treia fracțiune de glicoproteine a zonei transparente); 5 - plasmomolema oului; GGI, GGII - gynogamons; AGI, AGII - androgamone; Gal - glicoziltransferaza; NAG - N-acetilglucozamină
În mecanismul capacităţii, factorii hormonali sunt de mare importanţă, în primul rând progesteronul (hormonul corpului galben), care activează secreţia celulelor glandulare ale trompelor uterine. În timpul capacității, colesterolul din membrana plasmatică a spermatozoizilor se leagă de albumina din tractul genital feminin și sunt expuși receptorii celulelor germinale. Fertilizarea are loc în ampula trompei uterine. Fertilizarea este precedată de inseminare - interacțiunea și convergența gameților (interacțiune la distanță), datorită chimiotaxiei.
Faza a doua fertilizarea – interacțiunea de contact. Numeroase spermatozoizi se apropie de ovul și vin în contact cu membrana acestuia. Oul începe să se rotească în jurul axei sale cu o viteză de 4 rotații pe minut. Aceste mișcări sunt cauzate de bătaia cozilor spermatozoizilor și durează aproximativ 12 ore Spermatozoizii, în contact cu ovulul, pot lega zeci de mii de molecule de glicoproteină Zp3. Aceasta marchează începutul reacției acrozomale. Reacția acrozomală se caracterizează printr-o creștere a permeabilității plasmolemei spermatozoizilor la ionii de Ca 2 +, depolarizarea acesteia, ceea ce contribuie la fuziunea plasmolemei cu membrana acrozomală anterioară. Zona transparentă este în contact direct cu enzimele acrozomale. Enzimele îl distrug, sperma trece prin zona transparentă și

Orez. 21.5. Fertilizarea (conform lui Wasserman cu modificări):
1-4 - etape ale reacției acrozomale; 5 - zona pellucida(zona transparenta); 6 - spaţiul perivitelin; 7 - membrana plasmatica; 8 - granulă corticală; 8a - reacție corticală; 9 - pătrunderea spermatozoizilor în ovul; 10 - zona de reactie
intră în spațiul perivitelin, situat între zona transparentă și plasmolema oului. După câteva secunde, proprietățile plasmolemei celulei ou se schimbă și începe reacția corticală, iar după câteva minute se schimbă proprietățile zonei transparente (reacție zonală).
Inițierea celei de-a doua faze a fertilizării are loc sub influența polizaharidelor sulfatate ale zonei pellucide, care provoacă intrarea ionilor de calciu și sodiu în cap, spermatozoizi, înlocuindu-i cu ioni de potasiu și hidrogen și ruperea membranei acrozomului. Atașarea spermatozoizilor la ovul are loc sub influența grupului de carbohidrați al fracțiunii glicoproteice din zona transparentă a oului. Receptorii spermatozoizilor sunt o enzimă glicoziltransferază situată pe suprafața acrozomului capului, care

Orez. 21.6. Fazele de fertilizare și începutul zdrobirii (schemă):
1 - ovoplasmă; 1a - granule corticale; 2 - miez; 3 - zona transparenta; 4 - epiteliul folicular; 5 - spermatozoizi; 6 - corpuri de reducere; 7 - finalizarea diviziunii mitotice a ovocitului; 8 - tuberculul de fertilizare; 9 - înveliș de fertilizare; 10 - pronucleul feminin; 11 - pronucleul masculin; 12 - sincarion; 13 - prima diviziune mitotică a zigotului; 14 - blastomeri
„recunoaște” receptorul celulei germinale feminine. Membranele plasmatice de la locul de contact al celulelor germinale se îmbină și are loc plasmogamia - unirea citoplasmelor ambilor gameți.
La mamifere, doar un spermatozoid intră în ovul în timpul fertilizării. Un astfel de fenomen se numește monospermie. Fertilizarea este facilitată de sute de alți spermatozoizi implicați în inseminare. Enzimele secretate de acrozomi - spermolizinele (tripsină, hialuronidază) - distrug coroana radiantă, descompun glicozaminoglicanii din zona transparentă a oului. Celulele epiteliale foliculare detașate se lipesc împreună într-un conglomerat, care, urmând ovulul, se deplasează de-a lungul trompei uterine din cauza pâlpâirii cililor celulelor epiteliale ale membranei mucoase.

Orez. 21.7. Oul uman și zigotul (conform lui B.P. Khvatov):
A- ovul uman dupa ovulatie: 1 - citoplasma; 2 - miez; 3 - zona transparenta; 4 - celule epiteliale foliculare care formează o coroană radiantă; b- zigotul uman în stadiul de convergenţă a nucleilor masculin şi feminin (pronuclei): 1 - nucleu feminin; 2 - miez masculin
A treia fază. Capul și partea intermediară a regiunii caudale pătrund în ovoplasmă. După intrarea spermatozoidului în ovul, la periferia ovoplasmei, acesta devine mai dens (reacție zonală) și se formează înveliș de fertilizare.
Reacție corticală- fuziunea plasmolemei oului cu membranele granulelor corticale, în urma căreia conținutul granulelor pătrunde în spațiul perivitelin și acționează asupra moleculelor de glicoproteină ale zonei transparente (Fig. 21.5).
Ca urmare a acestei reacții de zonă, moleculele Zp3 sunt modificate și își pierd capacitatea de a fi receptori pentru spermatozoizi. Se formează o înveliș de fertilizare de 50 nm grosime, care împiedică polispermia - pătrunderea altor spermatozoizi.
Mecanismul reacției corticale implică afluxul de ioni de sodiu prin segmentul plasmalemei spermatozoidului, care este înglobat în plasmalema ovulului după terminarea reacției acrozomale. Ca rezultat, potențialul negativ de membrană al celulei devine slab pozitiv. Afluxul de ioni de sodiu determină eliberarea ionilor de calciu din depozitele intracelulare și o creștere a conținutului acestuia în hialoplasma oului. Aceasta este urmată de exocitoza granulelor corticale. Enzimele proteolitice eliberate din acestea rup legăturile dintre zona transparentă și plasmolema ovulului, precum și dintre spermatozoizi și zona transparentă. În plus, se eliberează o glicoproteină care leagă apa și o atrage în spațiul dintre plasmalemă și zona transparentă. Ca urmare, se formează un spațiu perivitelin. In cele din urma,
se eliberează un factor care contribuie la întărirea zonei transparente și la formarea unei învelișuri de fertilizare din aceasta. Datorită mecanismelor de prevenire a polispermiei, doar un nucleu haploid al spermatozoidului are posibilitatea de a fuziona cu un nucleu haploid al oului, ceea ce duce la restabilirea setului diploid caracteristic tuturor celulelor. Pătrunderea spermatozoidului în ovul după câteva minute îmbunătățește semnificativ procesele de metabolism intracelular, care este asociat cu activarea sistemelor sale enzimatice. Interacțiunea spermatozoizilor cu ovulul poate fi blocată de anticorpi împotriva substanțelor incluse în zona transparentă. Pe această bază se caută metode de contracepție imunologică.
După convergența pronucleilor feminin și masculin, care durează aproximativ 12 ore la mamifere, se formează un zigot - un embrion unicelular (Fig. 21.6, 21.7). În stadiul zigot, zone presupuse(lat. prezumtie- probabilitate, presupunere) ca surse de dezvoltare a secțiunilor corespunzătoare ale blastulei, din care se formează ulterior straturile germinale.
21.2.2. Clivajul și formarea blastulei
Despărțirea (fisiune)- diviziunea mitotică secvențială a zigotului în celule (blastomere) fără creșterea celulelor fiice până la dimensiunea mamei.
Blastomerele rezultate rămân unite într-un singur organism al embrionului. În zigot, se formează un fus mitotic între retrageri

Orez. 21.8. Embrionul uman în stadiile incipiente de dezvoltare (după Hertig și Rock):
A- stadiul a două blastomere; b- blastocist: 1 - embrioblast; 2 - trofoblast;
3 - cavitatea blastocistului

Orez. 21.9. Clivajul, gastrulația și implantarea embrionului uman (schemă): 1 - zdrobire; 2 - morula; 3 - blastocist; 4 - cavitatea blastocistului; 5 - embrio-blast; 6 - trofoblast; 7 - nodul germinativ: A - epiblast; b- hipoblast; 8 - înveliș de fertilizare; 9 - veziculă amniotică (ectodermică); 10 - mezenchim extraembrionar; 11 - ectoderm; 12 - endoderm; 13 - citotrofoblast; 14 - simplastotrofoblast; 15 - disc germinativ; 16 - goluri cu sânge matern; 17 - corion; 18 - picior amniotic; 19 - veziculă de gălbenuș; 20 - membrana mucoasă a uterului; 21 - oviduct
deplasându-se spre poli prin centrioli introduși de spermatozoid. Pronucleii intră în stadiul de profază cu formarea unui set combinat diploid de cromozomi de ovul și spermatozoizi.
După ce a trecut prin toate celelalte faze ale diviziunii mitotice, zigotul este împărțit în două celule fiice - blastomeri(din greaca. blastos- germeni, meros- Parte). Datorită absenței virtuale a perioadei G 1, în care celulele formate ca urmare a diviziunii cresc, celulele sunt mult mai mici decât cea maternă, prin urmare, dimensiunea embrionului în ansamblu în această perioadă, indiferent de numărul celulelor sale constitutive, nu depășește dimensiunea celulei originale - zigotul. Toate acestea au făcut posibilă apelarea procesului descris strivire(adică măcinarea) și celulele formate în procesul de zdrobire - blastomeri.
Clivajul zigotului uman începe la sfârșitul primei zile și este caracterizat ca complet neuniform asincron.În primele zile s-a întâmplat
merge încet. Prima zdrobire (diviziune) a zigotului se finalizează după 30 de ore, rezultând formarea a doi blastomeri acoperiți cu o membrană de fertilizare. Stadiul a două blastomere este urmat de stadiul a trei blastomere.
De la prima zdrobire a zigotului, se formează două tipuri de blastomeri - „întunecat” și „luminos”. „Ușoare”, mai mici, blastomerele sunt zdrobite mai repede și sunt aranjate într-un singur strat în jurul „întunericului” mare, care se află în mijlocul embrionului. Din blastomerele superficiale „ușoare” ia naștere ulterior trofoblast, legând embrionul cu corpul mamei și asigurarea nutriției acestuia. Interioare, „întunecate”, se formează blastomeri embrioblast, din care se formează corpul embrionului și organele extraembrionare (amnios, sacul vitelin, alantoide).
Începând din a 3-a zi, clivajul se desfășoară mai repede, iar în a 4-a zi embrionul este format din 7-12 blastomeri. După 50-60 de ore, se formează o acumulare densă de celule - morula, iar în ziua a 3-a-4 începe formarea blastociste- un balon gol umplut cu lichid (vezi Fig. 21.8; Fig. 21.9).
Blastocistul trece prin trompele uterine în uter în decurs de 3 zile și intră în cavitatea uterină după 4 zile. Blastocistul este liber în cavitatea uterină (blastocist liber)în termen de 2 zile (a 5-a și a 6-a zi). Până în acest moment, blastocistul crește în dimensiune datorită creșterii numărului de blastomere - celule embrioblast și trofoblaste - până la 100 și datorită absorbției crescute a secreției glandelor uterine de către trofoblast și producției active de lichid de către celulele trofoblaste. (vezi Fig. 21.9). Trofoblastul în primele 2 săptămâni de dezvoltare asigură hrana embrionului datorită produselor de degradare a țesuturilor materne (tip de nutriție histiotrofic),
Embrioblastul este situat sub forma unui mănunchi de celule germinale („mănunchi germinativ”), care este atașat intern de trofoblast la unul dintre polii blastocistului.
21.2.4. Implantare
Implantare (lat. implantare- ingrowth, rooting) - introducerea embrionului în membrana mucoasă a uterului.
Există două etape de implantare: adeziune(adeziune) când embrionul se atașează de suprafața interioară a uterului și invazie(imersie) - introducerea embrionului în țesutul membranei mucoase a uterului. În a 7-a zi, apar modificări în trofoblast și embrioblast asociate cu pregătirea pentru implantare. Blastocistul reține membrana de fertilizare. În trofoblast, numărul de lizozomi cu enzime crește, ceea ce asigură distrugerea (liza) țesuturilor peretelui uterin și contribuie astfel la introducerea embrionului în grosimea membranei sale mucoase. Microvilozitățile care apar în trofoblast distrug treptat membrana de fertilizare. Nodulul germinal se aplatizează și devine
V scut germinativ,în care încep pregătirile pentru prima etapă a gastrulației.
Implantarea durează aproximativ 40 de ore (vezi Fig. 21.9; Fig. 21.10). Concomitent cu implantarea, începe gastrulația (formarea straturilor germinale). Acest prima perioadă critică dezvoltare.
În prima etapă trofoblastul este atașat de epiteliul mucoasei uterine și în acesta se formează două straturi - citotrofoblastȘi simplastotrofoblast. În a doua etapă simplastotrofoblastul, producând enzime proteolitice, distruge mucoasa uterină. În același timp, cel vilozități trofoblastul, care pătrunde în uter, distruge secvențial epiteliul acestuia, apoi țesutul conjunctiv subiacent și pereții vaselor, iar trofoblastul intră în contact direct cu sângele vaselor materne. Format fosa de implantare,în care apar zone de hemoragii în jurul embrionului. Nutriția embrionului se realizează direct din sângele matern (tip de nutriție hematotrof). Din sângele mamei, fătul primește nu numai toate substanțele nutritive, ci și oxigenul necesar pentru respirație. În același timp, în mucoasa uterină din celulele țesutului conjunctiv bogate în glicogen, se formează deciduală celule. După ce embrionul este complet scufundat în fosa de implantare, orificiul format în mucoasa uterină este umplut cu sânge și produse de distrugere a țesuturilor mucoasei uterine. Ulterior, defectul mucoasei dispare, epiteliul este restabilit prin regenerare celulară.
Alimentația de tip hematotrof, care o înlocuiește pe cea histiotrofă, este însoțită de trecerea la o etapă calitativ nouă a embriogenezei - a doua fază a gastrulației și depunerea organelor extraembrionare.
21.3. GASTRULAȚIE ȘI ORGANOGENEZĂ
Gastrulația (din lat. gaster- stomac) - proces complex de modificări chimice și morfogenetice, însoțite de reproducere, creștere, mișcare direcționată și diferențiere a celulelor, având ca rezultat formarea straturilor germinale: exterioare (ectoderm), mijlociu (mezoderm) și interioare (endoderm) - surse de dezvoltare a complexului de organe axiale și muguri de țesut embrionar.
Gastrularea la om are loc în două etape. Primul stagiu(fapte-națiune) cade în ziua a 7-a, și a doua faza(imigrație) - în a 14-15-a zi de dezvoltare intrauterină.
La delaminare(din lat. lamina- farfurie), sau despicare, din materialul nodulului germinal (embrioblast) se formează două foi: foaia exterioară - epiblastși intern - hipoblast, cu fața în cavitatea blastocistului. Celulele epiblastice arată ca epiteliul prismatic pseudostratificat. Celule hipoblastice - mici cubice, cu cito-

Orez. 21.10. Embrioni umani 7,5 și 11 zile de dezvoltare în procesul de implantare în mucoasa uterină (după Hertig și Rocca):
A- 7,5 zile de dezvoltare; b- 11 zile de dezvoltare. 1 - ectodermul embrionului; 2 - endodermul embrionului; 3 - veziculă amniotică; 4 - mezenchim extraembrionar; 5 - citotrofoblast; 6 - simplastotrofoblast; 7 - glanda uterină; 8 - goluri cu sânge matern; 9 - epiteliul membranei mucoase a uterului; 10 - placa proprie a membranei mucoase a uterului; 11 - vilozități primare
plasma, formează un strat subțire sub epiblast. O parte din celulele epiblastice formează ulterior un perete sacul amniotic, care începe să se formeze în a 8-a zi. În zona inferioară a veziculei amniotice, rămâne un mic grup de celule epiblastice - materialul care va merge la dezvoltarea corpului embrionului și a organelor extraembrionare.
După delaminare, celulele sunt evacuate din foile exterioare și interioare în cavitatea blastocistului, care marchează formarea. mezenchimul extraembrionar. Până în a 11-a zi, mezenchimul crește până la trofoblast și se formează corionul - membrana viloasă a embrionului cu vilozități coriale primare (vezi Fig. 21.10).
A doua faza gastrulația are loc prin imigrarea (mișcarea) celulelor (Fig. 21.11). Mișcarea celulelor are loc în zona fundului veziculei amniotice. Fluxurile celulare apar în direcția din față în spate, spre centru și în profunzime ca urmare a reproducerii celulare (vezi Fig. 21.10). Acest lucru are ca rezultat formarea unei dungi primare. La capătul capului, dâra primară se îngroașă, formându-se primar, sau cap, nod(Fig. 21.12), de unde provine procesul capului. Procesul capului crește în direcția craniană între epi- și hipoblast și, în continuare, dă naștere la dezvoltarea notocordului embrionului, care determină axa embrionului, este baza pentru dezvoltarea oaselor scheletului axial. În jurul orei, coloana vertebrală se formează în viitor.
Materialul celular care se deplasează de la stria primară în spațiul dintre epiblast și hipoblast este situat paracordal sub formă de aripi mezo-dermale. O parte din celulele epiblastice este introdusă în hipoblast, participând la formarea endodermului intestinal. Ca rezultat, embrionul dobândește o structură cu trei straturi sub forma unui disc plat, constând din trei straturi germinale: ectoderm, mezodermȘi endoderm.
Factori care afectează mecanismele de gastrulație. Metodele și viteza de gastrulare sunt determinate de o serie de factori: gradientul metabolic dorsoventral, care determină asincronia reproducerii, diferențierii și mișcării celulelor; tensiunea superficială a celulelor și contactele intercelulare care contribuie la deplasarea grupurilor celulare. Un rol important îl au factorii inductivi. Conform teoriei centrelor organizatorice propusă de G. Spemann, în anumite părți ale embrionului apar inductori (factori de organizare), care au un efect inductor asupra altor părți ale embrionului, determinând dezvoltarea lor într-o anumită direcție. Există inductori (organizatori) de mai multe ordine care acționează secvenţial. De exemplu, s-a dovedit că organizatorul de ordinul întâi induce dezvoltarea plăcii neurale din ectoderm. În placa neurală apare un organizator de ordinul doi, care contribuie la transformarea unei secțiuni a plăcii neurale într-o cupă oculară etc.
În prezent, natura chimică a multor inductori (proteine, nucleotide, steroizi etc.) a fost elucidată. A fost stabilit rolul joncțiunilor gap în interacțiunile intercelulare. Sub acțiunea inductorilor care emană dintr-o celulă, celula indusă, care are capacitatea de a răspunde în mod specific, schimbă calea de dezvoltare. O celulă care nu este supusă acțiunii de inducție își păstrează potențele anterioare.
Diferențierea straturilor germinale și a mezenchimului începe la sfârșitul săptămânii a 2-a - începutul săptămânii a 3-a. O parte a celulelor este transformată în rudimente ale țesuturilor și organelor embrionului, cealaltă - în organe extraembrionare (vezi Capitolul 5, Schema 5.3).

Orez. 21.11. Structura unui embrion uman de 2 săptămâni. A doua etapă a gastrulației (schema):
A- sectiunea transversala a embrionului; b- discul germinal (vedere din laterala veziculei amniotice). 1 - epiteliul corionic; 2 - mezenchimul corion; 3 - goluri umplute cu sânge matern; 4 - baza vilozităților secundare; 5 - picior amniotic; 6 - veziculă amniotică; 7 - veziculă de gălbenuș; 8 - scut germinativ în procesul de gastrulare; 9 - banda primara; 10 - rudiment al endodermului intestinal; 11 - epiteliu gălbenuș; 12 - epiteliul membranei amniotice; 13 - nod primar; 14 - proces precordal; 15 - mezoderm extraembrionar; 16 - ectoderm extraembrionar; 17 - endoderm extraembrionar; 18 - ectoderm germinal; 19 - endoderm germinal

Orez. 21.12. Embrion uman 17 zile ("Crimeea"). Reconstituire grafică: A- disc embrionar (vedere de sus) cu proiecția angajărilor axiale și sistemul cardiovascular definitiv; b- secțiune sagitală (de mijloc) prin filele axiale. 1 - proiecția marcajelor bilaterale ale endocardului; 2 - proiecția anlagelor bilaterale ale celomului pericardic; 3 - proiecția anlagerilor bilaterale ale vaselor de sânge corporale; 4 - picior amniotic; 5 - vasele de sânge din piciorul amniotic; 6 - insule de sânge în peretele sacului vitelin; 7 - golful alantois; 8 - cavitatea veziculei amniotice; 9 - cavitatea sacului vitelin; 10 - trofoblast; 11 - proces cordal; 12 - nod de cap. Simboluri: bandă primară - hașurare verticală; nodulul cefalic primar este indicat prin încrucișări; ectoderm - fără umbrire; endoderm - linii; mezodermul extraembrionar - puncte (conform lui N. P. Barsukov și Yu. N. Shapovalov)
Diferențierea straturilor germinale și a mezenchimului, ducând la apariția primordiilor de țesut și organ, are loc nesimultan (heterocron), dar interconectate (integrativ), rezultând în formarea primordiilor tisulare.
21.3.1. Diferențierea ectodermului
Pe măsură ce ectodermul se diferențiază, se formează părți embrionare - ectoderm dermic, neuroectoderm, placode, placă precordală și ectoderm extragermeni, care este sursa formării căptușelii epiteliale a amnionului. Partea mai mică a ectodermului situat deasupra notocordului (neuroectoderm), dă naștere diferențierii tub neural Și creastă neurală. Ectodermul pielii dă naștere epiteliului scuamos stratificat al pielii (epidermă)și derivații săi, epiteliul corneei și conjunctivei ochiului, epiteliul cavității bucale, smalțul și cuticula dinților, epiteliul rectului anal, căptușeala epitelială a vaginului.
Neurulație- procesul de formare a tubului neural - decurge diferit în timp în diferite părți ale embrionului. Închiderea tubului neural începe în regiunea cervicală și apoi se extinde posterior și ceva mai lent în direcția craniană, unde se formează veziculele cerebrale. Aproximativ în a 25-a zi, tubul neural este complet închis, doar două deschideri neînchise la capetele anterior și posterior comunică cu mediul extern - neuropori anteriori si posteriori(Fig. 21.13). Neuroporul posterior corespunde canalul neurointestinal. După 5-6 zile, ambii neuropori cresc excesiv. Din tubul neural se formează neuronii și neuroglia ale creierului și măduvei spinării, retina ochiului și organul mirosului.
Odată cu închiderea pereților laterali ai pliurilor neuronale și formarea tubului neural, apare un grup de celule neuroectodermice, care se formează în joncțiunea ectodermului neural cu restul (pielei). Aceste celule, dispuse mai întâi în rânduri longitudinale de fiecare parte între tubul neural și ectoderm, formează creastă neurală. Celulele crestei neurale sunt capabile de migrare. În trunchi, unele celule migrează în stratul de suprafață al dermei, altele migrează în direcția ventrală, formând neuroni și neuroglia ganglionilor parasimpatici și simpatici, țesut cromafin și medula suprarenală. Unele celule se diferențiază în neuroni și neuroglia a ganglionilor spinali.
Celulele sunt eliberate din epiblast placă precordală, care este inclus în compoziția capului tubului intestinal. Din materialul plăcii precordale se dezvoltă ulterior epiteliul stratificat al părții anterioare a tubului digestiv și derivații acestuia. În plus, din placa precordală se formează epiteliul traheei, plămânilor și bronhiilor, precum și căptușeala epitelială a faringelui și esofagului, derivați ai pungilor branhiale - timusul etc.
Potrivit lui A. N. Bazhanov, sursa de formare a căptușelii esofagului și a tractului respirator este endodermul capului intestinului.

Orez. 21.13. Neurularea la embrionul uman:
A- vedere din spate; b- secțiuni transversale. 1 - neuropor anterior; 2 - neuropor posterior; 3 - ectoderm; 4 - placa neural; 5 - canal neural; 6 - mezoderm; 7 - coarda; 8 - endoderm; 9 - tub neural; 10 - creasta neural; 11 - creier; 12 - măduva spinării; 13 - canalul rahidian

Orez. 21.14. Embrionul uman în stadiul de formare a pliului trunchiului și a organelor extra-respiratorii (conform lui P. Petkov):
1 - simplastotrofoblast; 2 - citotrofoblast; 3 - mezenchim extraembrionar; 4 - locul piciorului amniotic; 5 - intestinul primar; 6 - cavitatea amnionică; 7 - ectoderm amnios; 8 - mezenchimul amniotic extraembrionar; 9 - cavitatea veziculei gălbenușului; 10 - endodermul veziculei gălbenușului; 11 - mezenchimul extraembrionar al sacului vitelin; 12 - alantoida. Săgețile indică direcția de formare a pliului trunchiului
Ca parte a ectodermului germinativ, sunt așezate placode, care sunt sursa dezvoltării structurilor epiteliale ale urechii interne. Din ectodermul extra-respirator se formează epiteliul amnionului și al cordonului ombilical.
21.3.2. Diferențierea endodermului
Diferențierea endodermului duce la formarea endodermului tubului intestinal în corpul embrionului și la formarea endodermului extraembrionar, care formează căptușeala veziculei viteline și alantoidei (Fig. 21.14).
Izolarea tubului intestinal începe cu apariția pliului trunchiului. Acesta din urmă, adâncindu-se, separă endodermul intestinal al viitorului intestin de endodermul extraembrionar al sacului vitelin. În partea posterioară a embrionului, intestinul rezultat include și acea parte a endodermului din care provine excrescența endodermică a alantoidei.
Din endodermul tubului intestinal se dezvoltă un epiteliu tegumentar cu un singur strat al stomacului, intestinelor și glandelor lor. În plus, din aceasta
dermul dezvoltă structuri epiteliale ale ficatului și pancreasului.
Endodermul extraembrionar dă naștere epiteliului sacului vitelin și alantoidei.
21.3.3. diferențierea mezodermului
Acest proces începe în a 3-a săptămână de embriogeneză. Secțiunile dorsale ale mezodermului sunt împărțite în segmente dense situate pe părțile laterale ale coardei - somite. Procesul de segmentare a mezodermului dorsal și de formare a somitelor începe în capul embrionului și se răspândește rapid caudal.
Embrionul din a 22-a zi de dezvoltare are 7 perechi de segmente, pe 25 - 14, pe 30 - 30, iar pe 35 - 43-44 perechi. Spre deosebire de somiți, secțiunile ventrale ale mezodermului (splanchnotom) nu sunt segmentate, ci împărțite în două foi - viscerală și parietală. O mică secțiune a mezodermului, care leagă somiții cu splanchnotomul, este împărțită în segmente - picioare segmentare (nefrogonotom). La capătul posterior al embrionului, segmentarea acestor diviziuni nu are loc. Aici, în loc de picioare segmentare, există un rudiment nefrogen nesegmentat (cordul nefrogen). Canalul paramezonefric se dezvoltă și din mezodermul embrionului.
Somiții se diferențiază în trei părți: miotomul, care dă naștere țesutului muscular scheletic striat, sclerotomul, care este sursa dezvoltării țesuturilor osoase și cartilajului și dermatomul, care formează baza țesutului conjunctiv al pielii - dermul. .
Din picioarele segmentare (nefrogonotomi) se dezvoltă epiteliul rinichilor, gonadelor și canalelor deferente, iar din canalul paramezonefric - epiteliul uterului, trompele uterine (oviducte) și epiteliul mucoasei primare a vaginului.
Foile parietale și viscerale ale splanhnotomului formează căptușeala epitelială a membranelor seroase - mezoteliul. Dintr-o parte a stratului visceral al mezodermului (placa mioepicardică) se dezvoltă învelișul mijlociu și exterior al inimii - miocardul și epicardul, precum și cortexul suprarenal.
Mezenchimul din corpul embrionului este sursa formării multor structuri - celule sanguine și organe hematopoietice, țesut conjunctiv, vase de sânge, țesut muscular neted, microglia (vezi capitolul 5). Din mezodermul extraembrionar se dezvoltă mezenchimul, dând naștere țesutului conjunctiv al organelor extraembrionare - amnios, alantoida, corion, veziculă gălbenușă.
Țesutul conjunctiv al embrionului și organele sale provizorii se caracterizează printr-o hidrofilitate ridicată a substanței intercelulare, o bogăție de glicozaminoglicani în substanța amorfă. Țesutul conjunctiv al organelor provizorii se diferențiază mai repede decât în rudimentele organului, ceea ce se datorează necesității de a stabili o legătură între embrion și corpul mamei și
asigurarea dezvoltării lor (de exemplu, placenta). Diferențierea mezenchimului corion apare precoce, dar nu are loc simultan pe toată suprafața. Procesul este cel mai activ în dezvoltarea placentei. Aici apar și primele structuri fibroase, care joacă un rol important în formarea și întărirea placentei din uter. Odată cu dezvoltarea structurilor fibroase ale stromei vilozităților, se formează succesiv fibre pre-colagen argirofile și apoi fibre de colagen.
În a 2-a lună de dezvoltare la embrionul uman, începe în primul rând diferențierea mezenchimului scheletic și a pielii, precum și a mezenchimului peretelui inimii și a vaselor mari de sânge.
Arterele de tip muscular și elastic de embrioni umani, precum și arterele vilozităților tulpinii (ancoră) ale placentei și ramurile acestora, conțin miocite netede desmin-negative, care au proprietatea unei contracție mai rapidă.
În a 7-a săptămână de dezvoltare a embrionului uman apar mici incluziuni lipidice în mezenchimul pielii și mezenchimul organelor interne, iar mai târziu (8-9 săptămâni) se formează celule adipoase. În urma dezvoltării țesutului conjunctiv al sistemului cardiovascular, țesutul conjunctiv al plămânilor și tubului digestiv se diferențiază. Diferențierea mezenchimului în embrioni umani (11-12 mm lungime) la a 2-a lună de dezvoltare începe cu creșterea cantității de glicogen din celule. În aceleași zone, activitatea fosfatazelor crește, iar mai târziu, în cursul diferențierii, se acumulează glicoproteine, se sintetizează ARN și proteine.
perioadă fructuoasă. Perioada fetală începe din săptămâna a 9-a și se caracterizează prin procese morfogenetice semnificative care apar atât în corpul fătului, cât și al mamei (Tabelul 21.1).
Tabelul 21.1. Un scurt calendar al dezvoltării intrauterine a unei persoane (cu completări conform R. K. Danilov, T. G. Borovoy, 2003)

Continuarea tabelului. 21.1

Continuarea tabelului. 21.1

Continuarea tabelului. 21.1

Continuarea tabelului. 21.1

Continuarea tabelului. 21.1

Continuarea tabelului. 21.1

Continuarea tabelului. 21.1

Sfârșitul mesei. 21.1

21.4. ORGANE EXTRA-GERMALE
Organele extraembrionare care se dezvoltă în procesul de embriogeneză în afara corpului embrionului îndeplinesc o varietate de funcții care asigură creșterea și dezvoltarea embrionului însuși. Unele dintre aceste organe din jurul embrionului sunt, de asemenea, numite membranelor embrionare. Aceste organe includ amniosul, sacul vitelin, alantoida, corionul, placenta (Fig. 21.15).
Sursele de dezvoltare ale țesuturilor organelor extraembrionare sunt trofectodermul și toate cele trei straturi germinale (Schema 21.1). Proprietățile generale ale țesăturii

Orez. 21.15. Dezvoltarea organelor extraembrionare în embrionul uman (schemă): 1 - veziculă amniotică; 1a - cavitatea amnionică; 2 - corpul embrionului; 3 - sacul vitelin; 4 - celomul extraembrionar; 5 - vilozități primare ale corionului; 6 - vilozități secundare ale corionului; 7 - tulpina de alantois; 8 - vilozități terțiare ale corionului; 9 - allan-tois; 10 - cordonul ombilical; 11 - corion neted; 12 - cotiledoane

Schema 21.1. Clasificarea țesuturilor organelor extraembrionare (conform lui V. D. Novikov, G. V. Pravotorov, Yu. I. Sklyanov)
organele ei extraembrionare și diferențele lor față de cele definitive sunt următoarele: 1) dezvoltarea țesuturilor este redusă și accelerată; 2) țesutul conjunctiv conține puține forme celulare, dar multă substanță amorfă bogată în glicozaminoglicani; 3) îmbătrânirea țesuturilor organelor extraembrionare are loc foarte rapid - până la sfârșitul dezvoltării fetale.
21.4.1. Amnion
Amnion- un organ temporar care asigură un mediu acvatic pentru dezvoltarea embrionului. A apărut în evoluție în legătură cu eliberarea vertebratelor din apă pe uscat. În embriogeneza umană, apare în a doua etapă a gastrulației, mai întâi ca o mică veziculă ca parte a epiblastului.
Peretele veziculei amniotice este format dintr-un strat de celule ale ectodermului extraembrionar și mezenchimului extraembrionar, formează țesutul său conjunctiv.
Amnionul crește rapid, iar la sfârșitul săptămânii a 7-a, țesutul său conjunctiv intră în contact cu țesutul conjunctiv al corionului. În același timp, epiteliul amniotic trece la tulpina amniotică, care ulterior se transformă în cordonul ombilical, iar în regiunea inelului ombilical se contopește cu învelișul epitelial al pielii embrionului.
Membrana amniotică formează peretele rezervorului umplut cu lichid amniotic, în care se află fătul (Fig. 21.16). Funcția principală a membranei amniotice este de a produce lichid amniotic, oferind un mediu pentru un organism în curs de dezvoltare și protejându-l de daune mecanice. Epiteliul amniosului, îndreptat spre cavitatea sa, nu numai că eliberează lichid amniotic, ci participă și la reabsorbția acestora. Lichidul amniotic se menține până la sfârșitul sarcinii compoziția necesarăși concentrația de sare. Amnionul îndeplinește și o funcție de protecție, împiedicând agenții nocivi să intre în făt.
Epiteliul amnionului în stadiile incipiente este plat cu un singur strat, format din celule poligonale mari, apropiate unele de altele, printre care există multe care se divid mitotic. La luna a 3-a de embriogeneză epiteliul se transformă într-unul prismatic. Pe suprafața epiteliului există microvilozități. Citoplasma conține întotdeauna mici picături de lipide și granule de glicogen. În părțile apicale ale celulelor există vacuole de diferite dimensiuni, al căror conținut este eliberat în cavitatea amnionică. Epiteliul amnionului din zona discului placentar este prismatic cu un singur strat, uneori cu mai multe rânduri, îndeplinește o funcție predominant secretorie, în timp ce epiteliul amnionului extraplacentar resorbe în principal lichidul amniotic.
În stroma țesutului conjunctiv a membranei amniotice se disting o membrană bazală, un strat de țesut conjunctiv fibros dens și un strat spongios de țesut conjunctiv fibros lax, care leagă

Orez. 21.16. Dinamica relației dintre embrion, organe extraembrionare și membrane uterine:
A- embrion uman 9,5 săptămâni de dezvoltare (micrograf): 1 - amnios; 2 - corion; 3 - formarea placentei; 4 - cordonul ombilical
amnios comun cu corion. În stratul de țesut conjunctiv dens, se pot distinge partea acelulară aflată sub membrana bazală și partea celulară. Acesta din urmă este format din mai multe straturi de fibroblaste, între care există o rețea densă de mănunchiuri subțiri de colagen și fibre reticulare strâns adiacente între ele, formând o rețea de formă neregulată orientată paralel cu suprafața cochiliei.
Stratul spongios este format dintr-un țesut conjunctiv mucos lax cu mănunchiuri rare de fibre de colagen, care sunt o continuare a celor care se află într-un strat de țesut conjunctiv dens, conectând amniosul cu corionul. Această conexiune este foarte fragilă și, prin urmare, ambele cochilii sunt ușor de separat unul de celălalt. Substanța principală a țesutului conjunctiv conține mulți glicozaminoglicani.
21.4.2. Sacul vitelin
Sacul vitelin- cel mai vechi organ extraembrionar din evolutie, care a luat nastere ca organ care depune nutrienti (galbenusul) necesari dezvoltarii embrionului. La om, aceasta este o formațiune rudimentară (veziculă de gălbenuș). Este format din endoderm extraembrionar și mezoderm extraembrionar (mezenchim). Apărând în a 2-a săptămână de dezvoltare la om, vezicula gălbenușă în nutriția embrionului ia

Orez. 21.16. Continuare
b- diagrama: 1 - membrana musculara a uterului; 2- decidua bazalis; 3 - cavitatea amnionică; 4 - cavitatea sacului vitelin; 5 - celomul extraembrionar (cavitatea coriala); 6- decidua capsularis; 7 - decidua parietalis; 8 - cavitatea uterină; 9 - colul uterin; 10 - embrion; 11 - vilozități terțiare ale corionului; 12 - alantoida; 13 - mezenchimul cordonului ombilical: A- vasele de sânge ale vilozităților coriale; b- lacune cu sânge matern (după Hamilton, Boyd și Mossman)
participarea este foarte scurtă, deoarece din a 3-a săptămână de dezvoltare se stabilește o legătură între făt și corpul mamei, adică nutriția hematotrofică. Sacul vitelin al vertebratelor este primul organ în peretele căruia se dezvoltă insule de sânge, formând primele celule sanguine și primele vase de sânge care furnizează oxigen și substanțe nutritive fătului.
Pe măsură ce se formează pliul trunchiului, care ridică embrionul deasupra sacului vitelin, se formează un tub intestinal, în timp ce sacul vitelin este separat de corpul embrionului. Legătura embrionului cu sacul vitelin rămâne sub forma unui funiculu gol, numit tulpină vitelină. Ca organ hematopoietic, sacul vitelin funcționează până în săptămâna 7-8, apoi suferă o dezvoltare inversă și rămâne în cordonul ombilical sub forma unui tub îngust care servește ca conductor al vaselor de sânge către placentă.
21.4.3. Allantois
Alantoida este un proces asemănător unui deget mic în partea caudală a embrionului, care crește în tulpina amniotică. Este derivat din sacul vitelin și este format din endodermul extraembrionar și mezodermul visceral. La om, alantoida nu atinge o dezvoltare semnificativă, dar rolul său în asigurarea nutriției și respirației embrionului este încă mare, deoarece vasele situate în cordonul ombilical cresc de-a lungul acestuia spre corion. Partea proximală a alantoidei este situată de-a lungul tulpinii gălbenușului, iar partea distală, în creștere, crește în golul dintre amnios și corion. Este un organ de schimb de gaze și excreție. Oxigenul este livrat prin vasele alantoidei, iar produșii metabolici ai embrionului sunt eliberați în alantoide. La a 2-a lună de embriogeneză, alantoida este redusă și se transformă într-un cordon de celule, care, împreună cu vezicula vitelină redusă, face parte din cordonul ombilical.
21.4.4. cordon ombilical
Cordonul ombilical sau cordonul ombilical este un cordon elastic care leagă embrionul (fătul) de placentă. Este acoperit de o membrană amniotică care înconjoară un țesut conjunctiv mucos cu vase de sânge (două artere ombilicale și o venă) și vestigii ale sacului vitelin și alantoidei.
Țesutul conjunctiv mucos, numit „jeleu Wharton”, asigură elasticitatea cordonului, protejează vasele ombilicale de compresie, asigurând astfel un aport continuu de nutrienți și oxigen embrionului. Împreună cu aceasta, împiedică pătrunderea agenților nocivi din placentă la embrion prin mijloace extravasculare și astfel îndeplinește o funcție de protecție.
Metodele imunocitochimice au stabilit că în vasele de sânge ale cordonului ombilical, placentă și embrion există celule musculare netede (SMC) eterogene. În vene, spre deosebire de artere, s-au găsit SMC-uri pozitive pentru desmină. Acestea din urmă asigură contracții tonice lente ale venelor.
21.4.5. Chorion
Chorion, sau teaca viloasa, apare pentru prima dată la mamifere, se dezvoltă din trofoblast și mezodermul extraembrionar. Inițial, trofoblastul este reprezentat de un strat de celule care formează vilozități primare. Ele secretă enzime proteolitice, cu ajutorul cărora mucoasa uterină este distrusă și se realizează implantarea. În săptămâna a 2-a, trofoblastul capătă o structură cu două straturi datorită formării în el a stratului celular interior (citotrofoblast) și a stratului exterior simplastic (simplastotrofoblast), care este un derivat al stratului celular. Mezenchimul extraembrionar care apare de-a lungul periferiei embrioblastului (la om în săptămâna 2-3 de dezvoltare) crește până la trofoblast și formează vilozități epiteliomezenchimale secundare cu acesta. Din acest moment, trofoblastul se transformă într-un corion sau membrană viloasă (vezi Fig. 21.16).
La începutul săptămânii a 3-a, capilarele sanguine cresc în vilozitățile corionului și se formează vilozități terțiare. Aceasta coincide cu începutul nutriției hematotrofice a embrionului. Dezvoltarea ulterioară a corionului este asociată cu două procese - distrugerea mucoasei uterine datorită activității proteolitice a stratului exterior (simplastic) și dezvoltarea placentei.
21.4.6. Placenta
Placenta (locul copiilor) omul aparține tipului de placentă viloasă hemocorală discoidală (vezi Fig. 21.16; Fig. 21.17). Acesta este un organ temporar important cu o varietate de funcții care asigură o legătură între făt și corpul mamei. În același timp, placenta creează o barieră între sângele mamei și făt.
Placenta este formată din două părți: germinale sau fetală (pars fetalis)și maternă (pars materna). Partea fetală este reprezentată de un corion ramificat și o membrană amniotică care aderă la corion din interior, iar partea maternă este o mucoasă uterină modificată care este respinsă în timpul nașterii. (decidua bazalis).
Dezvoltarea placentei începe în a 3-a săptămână, când vasele încep să crească în forma vilozităților secundare și terțiare și se termină la sfârșitul lunii a 3-a de sarcină. În săptămâna 6-8 în jurul vaselor

Orez. 21.17. Placenta hemocoriala. Dinamica dezvoltării vilozităților coriale: A- structura placentei (săgețile indică circulația sângelui în vase și într-unul din golurile de unde a fost îndepărtată vilozitatea): 1 - epiteliu amnios; 2 - placa coriala; 3 - vilozități; 4 - fibrinoid; 5 - veziculă de gălbenuș; 6 - cordonul ombilical; 7 - septul placentar; 8 - lacună; 9 - artera spirală; 10 - stratul bazal al endometrului; 11 - miometru; b- structura vilozităților trofoblastice primare (săptămâna I); V- structura vilozităților epitelial-mezenchimatoase secundare a corionului (săptămâna a 2-a); G- structura vilozităților coriale terțiare - epitelial-mezenchimatoase cu vase de sânge (săptămâna a 3-a); d- structura vilozităților coriale (luna a 3-a); e- structura vilozităților coriale (luna a 9-a): 1 - spațiu intervilos; 2 - microvilozități; 3 - simplastotrofoblast; 4 - nuclei simplastotrofoblasti; 5 - citotrofoblast; 6 - nucleul citotrofoblastului; 7 - membrana bazala; 8 - spațiu intercelular; 9 - fibroblast; 10 - macrofage (celule Kashchenko-Hofbauer); 11 - endoteliocit; 12 - lumenul unui vas de sânge; 13 - eritrocit; 14 - membrana bazală a capilarului (după E. M. Schwirst)
elementele de țesut conjunctiv sunt diferențiate. Vitaminele A și C joacă un rol important în diferențierea fibroblastelor și sinteza colagenului de către acestea, fără un aport suficient al căruia puterea legăturii dintre embrion și corpul mamei este întreruptă și se creează amenințarea avortului spontan.
Substanța principală a țesutului conjunctiv al corionului conține o cantitate semnificativă de acizi hialuronic și condroitinsulfuric, care sunt asociate cu reglarea permeabilității placentare.
Odată cu dezvoltarea placentei, are loc distrugerea mucoasei uterine, datorită activității proteolitice a corionului, și schimbarea nutriției histiotrofice în hematotrofă. Aceasta înseamnă că vilozitățile corionului sunt spălate de sângele mamei, care s-a revărsat din vasele distruse ale endometrului în lacune. Cu toate acestea, sângele mamei și al fătului în condiții normale nu se amestecă niciodată.
bariera hematocoriala, care separă ambele fluxuri sanguine, este format din endoteliul vaselor fetale, țesutul conjunctiv care înconjoară vasele, epiteliul vilozităților coriale (citotrofoblast și simplastotrofoblast) și, în plus, din fibrinoid, care acoperă uneori vilozitățile din exterior.
germinal, sau fetal, parte placenta până la sfârșitul lunii a 3-a este reprezentată de o placă corionica ramificată, formată din țesut conjunctiv fibros (colagen), acoperită cu cito- și simplastotrofoblast (o structură multinucleară care acoperă citotrofoblastul reducător). Vilozitățile ramificate ale corionului (tulpină, ancoră) sunt bine dezvoltate doar pe partea orientată spre miometru. Aici trec prin toată grosimea placentei și cu vârfurile lor plonjează în partea bazală a endometrului distrus.
Epiteliul corionic, sau citotrofoblast, în stadiile incipiente de dezvoltare este reprezentat de un epiteliu cu un singur strat cu nuclei ovali. Aceste celule se reproduc prin mitoză. Ei dezvoltă simplastotrofoblast.
Simplastotrofoblastul conține un număr mare de diferite enzime proteolitice și oxidative (ATPaze, alcaline și acide).

Orez. 21.18. Secțiunea vilozităților coriale a unui embrion uman de 17 zile („Crimeea”). Micrografie:
1 - simplastotrofoblast; 2 - citotrofoblast; 3 - mezenchimul corion (după N. P. Barsukov)
- total aproximativ 60), care este asociat cu rolul său în procesele metabolice dintre mamă și făt. Vezicule pinocitare, lizozomi și alte organite sunt detectate în citotrofoblast și în simplast. Începând din luna a 2-a, epiteliul corionic se subțiază și este înlocuit treptat de simplastotrofoblast. În această perioadă, simplastotrofoblastul depășește citotrofoblastul în grosime. În săptămâna a 9-a-10, simplastul devine mai subțire, iar numărul de nuclee din el crește. Pe suprafața simplastului îndreptată spre lacune apar numeroase microvilozități sub forma unei margini de pensulă (vezi Fig. 21.17; Fig. 21.18, 21.19).
Între simplastotrofoblast și trofoblast celular există spații submicroscopice în formă de fante, ajungând pe alocuri până la membrana bazală a trofoblastului, ceea ce creează condiții pentru pătrunderea bilaterală a substanțelor trofice, hormonilor etc.
În a doua jumătate a sarcinii și, mai ales, la sfârșitul acesteia, trofoblastul devine foarte subțire, iar vilozitățile sunt acoperite cu o masă oxifilă asemănătoare fibrinei, care este un produs al coagulării plasmatice și al defalcării trofoblastului („Langhans). fibrinoid”).
Odată cu creșterea vârstei gestaționale, numărul de macrofage și fibroblaste diferențiate producătoare de colagen scade, apărând

Orez. 21.19. Bariera placentară la a 28-a săptămână de sarcină. Micrografie electronică, mărire 45.000 (conform lui U. Yu. Yatsozhinskaya):
1 - simplastotrofoblast; 2 - citotrofoblast; 3 - membrana bazală a trofoblastului; 4 - membrana bazală a endoteliului; 5 - endoteliocit; 6 - eritrocit în capilar
fibrocite. Numărul de fibre de colagen, deși în creștere, rămâne nesemnificativ în majoritatea vilozităților până la sfârșitul sarcinii. Majoritatea celulelor stromale (miofibroblaste) se caracterizează printr-un conținut crescut de proteine contractile citoscheletice (vimentină, desmină, actină și miozină).
Unitatea structurală și funcțională a placentei formate este cotiledonul, format din vilozitatea tulpină („ancoră”) și
ramuri secundare și terțiare (finale). Numărul total de cotiledoane din placentă ajunge la 200.
Partea mamă placenta este reprezentată de o placă bazală și septuri de țesut conjunctiv care separă cotiledoanele unul de celălalt, precum și goluri umplute cu sânge matern. Celulele trofoblaste (trofoblastul periferic) se găsesc și în punctele de contact dintre vilozitățile stem și înveliș.
În stadiile incipiente ale sarcinii, vilozitățile coriale distrug straturile principale ale membranei uterine cele mai apropiate de făt, iar în locul lor se formează lacune pline cu sânge matern, în care vilozitățile coriale atârnă liber.
Părțile adânci nedistruse ale membranei care se desprind, împreună cu trofoblastul, formează placa bazală.
Stratul bazal al endometrului (lamina bazala)- tesutul conjunctiv al mucoasei uterine deciduală celule. Aceste celule mari de țesut conjunctiv, bogate în glicogen, sunt situate în straturile profunde ale mucoasei uterine. Au limite clare, nuclei rotunjiți și citoplasmă oxifilă. În a 2-a lună de sarcină, celulele deciduale sunt semnificativ mărite. În citoplasma lor, pe lângă glicogen, sunt detectate lipide, glucoză, vitamina C, fier, esteraze nespecifice, dehidrogenaza acizilor succinic și lactic. În placa bazală, mai des la locul de atașare a vilozităților de partea maternă a placentei, se găsesc grupuri de celule citotrofoblaste periferice. Se aseamănă cu celulele deciduale, dar diferă printr-o bazofilie mai intensă a citoplasmei. O substanță amorfă (fibrinoidul lui Rohr) este situată pe suprafața plăcii bazale cu fața către vilozitățile coriale. Fibrinoidul joacă un rol esențial în asigurarea homeostaziei imunologice în sistemul mamă-făt.
O parte a cochiliei principale care cade, situată la marginea corionului ramificat și neted, adică de-a lungul marginii discului placentar, nu este distrusă în timpul dezvoltării placentei. Creștend strâns până la corion, se formează placă de capăt,împiedicând scurgerea sângelui din lacunele placentei.
Sângele din lacune circulă continuu. Provine din arterele uterine, care intră aici din membrana musculară a uterului. Aceste artere trec de-a lungul septurilor placentare și se deschid în lacune. Sângele matern curge din placentă prin vene care provin din lacunele cu găuri mari.
Formarea placentei se încheie la sfârșitul lunii a 3-a de sarcină. Placenta asigură nutriția, respirația tisulară, creșterea, reglarea rudimentelor organelor fetale formate în acest moment, precum și protecția acesteia.
Funcțiile placentei. Principalele funcții ale placentei: 1) respirator; 2) transportul nutrienților; apă; electroliți și imunoglobuline; 3) excretor; 4) endocrin; 5) participarea la reglarea contracției miometrului.
Suflare fătul este furnizat de oxigenul atașat de hemoglobina maternă, care difuzează prin placentă în sângele fetal, unde se combină cu hemoglobina fetală.
(HbF). CO 2 asociat cu hemoglobina fetală din sângele fătului difuzează și prin placentă, intră în sângele mamei, unde se combină cu hemoglobina maternă.
Transport dintre toți nutrienții necesari dezvoltării fătului (glucoză, aminoacizi, acizi grași, nucleotide, vitamine, minerale) provine din sângele mamei prin placentă în sângele fetal și, dimpotrivă, produsele metabolice excretate din sângele mamei. pătrunde în sângele mamei din corpul său (funcția excretorie). Electroliții și apa trec prin placentă prin difuzie și prin pinocitoză.
Veziculele pinocitare ale simplastotrofoblastului sunt implicate în transportul imunoglobulinelor. Imunoglobulina care intră în sângele fătului îl imunizează pasiv împotriva posibilei acțiuni a antigenelor bacteriene care pot intra în timpul bolilor materne. După naștere, imunoglobulina maternă este distrusă și înlocuită cu nou sintetizată în organismul copilului sub acțiunea antigenelor bacteriene asupra acesteia. Prin placentă, IgG, IgA pătrund în lichidul amniotic.
functia endocrina este una dintre cele mai importante, intrucat placenta are capacitatea de a sintetiza si secreta o serie de hormoni care asigura interactiunea dintre embrion si corpul mamei pe tot parcursul sarcinii. Locul de producere a hormonului placentar este citotrofoblastul și în special simplastotrofoblastul, precum și celulele deciduale.
Placenta este una dintre primele care sintetizează gonadotropină corionică, a cărui concentrație crește rapid în săptămâna 2-3 de sarcină, atingând un maxim în săptămâna 8-10, iar în sângele fetal este de 10-20 de ori mai mare decât în sângele mamei. Hormonul stimulează producția de hormon adrenocorticotrop (ACTH) de către glanda pituitară, îmbunătățește secreția de corticosteroizi.
joacă un rol important în dezvoltarea sarcinii lactogen placentar, care are activitatea prolactinei si hormonului luteotrop hipofizar. Sprijină steroidogeneza în corpul galben al ovarului în primele 3 luni de sarcină și, de asemenea, participă la metabolismul carbohidraților și proteinelor. Concentratia lui in sangele mamei creste progresiv in luna a 3-4 de sarcina si apoi continua sa creasca, ajungand la maxim pana in luna a 9-a. Acest hormon, împreună cu prolactina hipofizară maternă și fetală, joacă un rol în producerea de surfactant pulmonar și în osmoreglarea fetoplacentară. Concentrația sa mare se găsește în lichidul amniotic (de 10-100 de ori mai mult decât în sângele mamei).
În corion, precum și în decidua, se sintetizează progesteronul și pregnandiolul.
Progesteronul (produs mai întâi de corpul galben în ovar, iar din săptămâna 5-6 în placentă) inhibă contracțiile uterine, stimulează creșterea acestuia, are efect imunosupresor, suprimând reacția de respingere fetală. Aproximativ 3/4 din progesteronul din corpul mamei este metabolizat și transformat în estrogen, iar o parte este excretat în urină.
Estrogenii (estradiol, estronă, estriol) sunt produși în simmplasto-trofoblastul vilozităților placentare (coriale) la mijlocul sarcinii și până la sfârșitul sarcinii.
Sarcina activitatea lor crește de 10 ori. Ele provoacă hiperplazie și hipertrofie a uterului.
În plus, în placentă se sintetizează hormoni melanocitari și adrenocorticotropi, somatostatina etc.
Placenta conține poliamine (spermină, spermidină), care afectează îmbunătățirea sintezei ARN în celulele musculare netede ale miometrului, precum și oxidaze care le distrug. Un rol important îl au aminoxidazele (histamiza, monoaminoxidaza), care distrug aminele biogene - histamina, serotonina, tiramina. În timpul sarcinii, activitatea lor crește, ceea ce contribuie la distrugerea aminelor biogene și la scăderea concentrației acestora din urmă în placentă, miometru și sângele matern.
În timpul nașterii, histamina și serotonina, împreună cu catecolaminele (norepinefrină, adrenalină), sunt stimulente ale activității contractile celulelor musculare netede (SMC) ale uterului, iar până la sfârșitul sarcinii, concentrația lor crește semnificativ datorită unei scăderi accentuate ( de 2 ori) în activitatea aminooxidazelor (histaminaza etc.).
Cu activitate de muncă slabă, există o creștere a activității aminooxidazelor, de exemplu, histaminaza (de 5 ori).
Placenta normală nu este o barieră absolută pentru proteine. În special, la sfârșitul lunii a 3-a de sarcină, fetoproteina pătrunde într-o cantitate mică (aproximativ 10%) din făt în sângele mamei, dar organismul matern nu respinge acest antigen, deoarece citotoxicitatea limfocitelor materne scade în timpul sarcina.
Placenta împiedică trecerea unui număr de celule materne și a anticorpilor citotoxici către făt. Rolul principal în aceasta este jucat de fibrinoid, care acoperă trofoblastul atunci când este parțial deteriorat. Acest lucru previne intrarea antigenelor placentare și fetale în spațiul intervilos și, de asemenea, slăbește „atacul” umoral și celular al mamei împotriva fătului.
În concluzie, remarcăm principalele trăsături ale stadiilor incipiente de dezvoltare a embrionului uman: 1) tip asincron de zdrobire completă și formarea de blastomere „luminoase” și „întunecate”; 2) izolarea precoce și formarea organelor extraembrionare; 3) formarea precoce a veziculei amniotice și absența pliurilor amniotice; 4) prezența a două mecanisme în stadiul de gastrulație - delaminarea și imigrația, timp în care are loc și dezvoltarea organelor provizorii; 5) tip interstițial de implantare; 6) dezvoltarea puternică a amniosului, corionului, placentei și dezvoltarea slabă a sacului vitelin și alantoidei.
21.5. SISTEM MAMA-FET
Sistemul mamă-făt apare în timpul sarcinii și include două subsisteme - corpul mamei și corpul fătului, precum și placenta, care este legătura dintre ele.
Interacțiunea dintre corpul mamei și cel al fătului este asigurată în primul rând de mecanisme neuroumorale. În același timp, în ambele subsisteme se disting următoarele mecanisme: receptor, perceperea informației, reglator, procesarea acesteia și executiv.
Mecanismele receptorilor din corpul mamei sunt localizate în uter sub formă de terminații nervoase sensibile, care sunt primele care percep informații despre starea fătului în curs de dezvoltare. În endometru există chimio-, mecano- și termoreceptori, iar în vasele de sânge - baroreceptori. Terminațiile nervoase receptoare de tip liber sunt în special numeroase în pereții venei uterine și în decidua în zona de atașare a placentei. Iritarea receptorilor uterini determină modificări ale intensității respirației, tensiunii arteriale în corpul mamei, ceea ce asigură conditii normale pentru fătul în curs de dezvoltare.
Mecanismele de reglare ale corpului mamei includ părți ale sistemului nervos central (lobul temporal al creierului, hipotalamus, formațiune reticulară mezencefalică), precum și sistemul hipotalamo-endocrin. important functie de reglementare efectuează hormoni: hormoni sexuali, tiroxina, corticosteroizi, insulină, etc. Astfel, în timpul sarcinii, se produce o creștere a activității cortexului suprarenal al mamei și o creștere a producției de corticosteroizi, care sunt implicați în reglarea metabolismului fetal. Placenta produce gonadotropină corionică, care stimulează formarea ACTH hipofizar, care activează activitatea cortexului suprarenal și intensifică secreția de corticosteroizi.
Aparatul neuroendocrin reglator al mamei asigură păstrarea sarcinii, nivelul necesar de funcționare a inimii, vaselor de sânge, organelor hematopoietice, ficatului și nivelul optim al metabolismului, gazelor, în funcție de nevoile fătului.
Mecanismele receptorilor corpului fetal percep semnale despre modificările din corpul mamei sau despre propria homeostazie. Se găsesc în pereții arterelor și venelor ombilicale, în gura venelor hepatice, în pielea și intestinele fătului. Iritarea acestor receptori duce la o modificare a ritmului cardiac al fătului, a vitezei fluxului sanguin în vasele acestuia, afectează conținutul de zahăr din sânge etc.
Mecanismele de reglare neuroumorale ale corpului fetal se formează în procesul de dezvoltare. Primele reacții motorii la făt apar în luna a 2-3-a de dezvoltare, ceea ce indică maturizarea centrilor nervoși. Mecanismele de reglare a homeostaziei gazelor se formează la sfârșitul celui de-al doilea trimestru de embriogeneză. Începutul funcționării glandei endocrine centrale - glanda pituitară - se notează în luna a 3-a de dezvoltare. Sinteza corticosteroizilor în glandele suprarenale ale fătului începe în a doua jumătate a sarcinii și crește odată cu creșterea acesteia. Fătul are sinteza crescută de insulină, ceea ce este necesar pentru a-i asigura creșterea asociată cu metabolismul carbohidraților și energetic.
Acțiunea sistemelor de reglare neuroumorală ale fătului este îndreptată către mecanismele executive - organele fătului, care asigură o modificare a intensității respirației, a activității cardiovasculare, a activității musculare etc., și asupra mecanismelor care determină modificarea. la nivelul schimbului de gaze, metabolism, termoreglare și alte funcții.
În asigurarea conexiunilor în sistemul mamă-făt, un rol deosebit de important îl joacă placenta, care este capabil nu numai să acumuleze, ci și să sintetizeze substanțele necesare dezvoltării fătului. Placenta indeplineste functii endocrine, producand o serie de hormoni: progesteron, estrogen, gonadotropina corionica umana (CG), lactogen placentar etc. Prin placenta se realizeaza conexiuni umorale si neuronale intre mama si fat.
Există, de asemenea, conexiuni umorale extraplacentare prin membranele fetale și lichidul amniotic.
Canalul de comunicare umoral este cel mai extins și mai informativ. Prin ea intră oxigenul și dioxid de carbon, proteine, carbohidrați, vitamine, electroliți, hormoni, anticorpi etc. (Fig. 21.20). In mod normal, substantele straine nu patrund in corpul mamei prin placenta. Ele pot începe să pătrundă numai în condiții de patologie, atunci când funcția de barieră a placentei este afectată. O componentă importantă a conexiunilor umorale sunt conexiunile imunologice care asigură menținerea homeostaziei imune în sistemul mamă-făt.
În ciuda faptului că organismele mamei și ale fătului sunt străine genetic în compoziția proteinelor, conflictul imunologic nu apare de obicei. Acest lucru este asigurat de o serie de mecanisme, printre care sunt esențiale următoarele: 1) proteine sintetizate de simplastotrofoblast, care inhibă răspunsul imun al organismului mamei; 2) gonadotropina corionica si lactogenul placentar, care sunt in concentratie mare pe suprafata simplastotrofoblastului; 3) efectul imunomasking al glicoproteinelor fibrinoidului pericelular al placentei, încărcate la fel ca limfocitele din sângele de spălat, este negativ; 4) proprietățile proteolitice ale trofoblastului contribuie și ele la inactivarea proteinelor străine.
Apele amniotice, care conțin anticorpi care blochează antigenele A și B, caracteristice sângelui unei femei însărcinate, participă și ele la apărarea imunitară și nu le permit să intre în sângele fătului.
Organismele materne și fetale sunt un sistem dinamic de organe omoloage. Înfrângerea oricărui organ al mamei duce la o încălcare a dezvoltării organului cu același nume al fătului. Deci, dacă o femeie însărcinată suferă de diabet, în care producția de insulină este redusă, atunci fătul are o creștere a greutății corporale și o creștere a producției de insulină în insulele pancreatice.
Într-un experiment pe animale, s-a stabilit că serul de sânge al unui animal din care a fost îndepărtată o parte a unui organ stimulează proliferarea în organul cu același nume. Cu toate acestea, mecanismele acestui fenomen nu sunt bine înțelese.
Conexiunile nervoase includ canalele placentare și extraplacentare: placentare - iritația baro- și chemoreceptorilor în vasele placentei și cordonului ombilical și extraplacentare - intrarea în sistemul nervos central al mamei a iritațiilor asociate creșterii fetale etc.
Prezența conexiunilor neuronale în sistemul mamă-făt este confirmată de datele privind inervația placentei, un conținut ridicat de acetilcolină în ea,

Orez. 21.20. Transportul substanțelor prin bariera placentară
dezvoltarea fetală în cornul uterin denervat al animalelor de experiment etc.
În procesul de formare a sistemului mamă-făt, există o serie de perioade critice, cele mai importante pentru stabilirea interacțiunii între cele două sisteme, menite să creeze condiții optime pentru dezvoltarea fătului.
21.6. PERIOADE CRITICE DE DEZVOLTARE
În timpul ontogenezei, în special embriogenezei, există perioade de sensibilitate mai mare a celulelor germinale în curs de dezvoltare (în timpul progenezei) și a embrionului (în timpul embriogenezei). Acest lucru a fost observat pentru prima dată de medicul australian Norman Gregg (1944). Embriologul rus P. G. Svetlov (1960) a formulat teoria perioadelor critice de dezvoltare și a testat-o experimental. Esența acestei teorii
este de a aproba pozitia generala că fiecare etapă a dezvoltării embrionului în ansamblu și a organelor sale individuale începe cu o perioadă relativ scurtă de restructurare calitativ nouă, însoțită de determinarea, proliferarea și diferențierea celulelor. În acest moment, embrionul este cel mai susceptibil la efecte dăunătoare de diferite naturi (expunerea la raze X, medicamente etc.). Astfel de perioade în progeneză sunt spermiogeneza și ovogeneza (meioza), iar în embriogeneză - fertilizarea, implantarea (în timpul căreia are loc gastrulația), diferențierea straturilor germinale și depunerea organelor, perioada de placentare (maturarea finală și formarea placentei), formarea multor sisteme funcționale, nașterea.
Printre organele și sistemele umane în curs de dezvoltare, un loc special aparține creierului, care în stadiile incipiente acționează ca organizator principal al diferențierii țesutului înconjurător și primordiilor de organe (în special organele senzoriale), iar mai târziu este caracterizat de celule intensive. reproducere (aproximativ 20.000 pe minut), care necesită condiții trofice optime.
În perioadele critice, factorii exogeni dăunători pot fi substanțe chimice, inclusiv multe medicamente, radiații ionizante (de exemplu, raze X în doze de diagnosticare), hipoxie, înfometare, medicamente, nicotină, viruși etc.
Substanțele chimice și medicamentele care traversează bariera placentară sunt deosebit de periculoase pentru făt în primele 3 luni de sarcină, deoarece nu sunt metabolizate și se acumulează în concentrații mari în țesuturile și organele acestuia. Drogurile interferează cu dezvoltarea creierului. Inaniția, virusurile provoacă malformații și chiar moarte intrauterină (Tabelul 21.2).
Deci, în ontogeneza umană, se disting mai multe perioade critice de dezvoltare: în progeneză, embriogeneză și viața postnatală. Acestea includ: 1) dezvoltarea celulelor germinale - ovogeneza si spermatogeneza; 2) fertilizare; 3) implantare (7-8 zile de embriogeneză); 4) dezvoltarea rudimentelor axiale ale organelor și formarea placentei (3–8 săptămâni de dezvoltare); 5) stadiul creșterii creierului îmbunătățit (15-20 săptămâni); 6) formarea principalelor sisteme funcționale ale corpului și diferențierea aparatului reproducător (20-24 săptămâni); 7) naștere; 8) perioada neonatală (până la 1 an); 9) pubertate (11-16 ani).
Metode și măsuri de diagnosticare pentru prevenirea anomaliilor dezvoltării umane. Pentru a identifica anomaliile în dezvoltarea umană, medicina modernă are o serie de metode (neinvazive și invazive). Deci, toate femeile însărcinate de două ori (la 16-24 și 32-36 săptămâni) sunt ultrasonografie, care permite depistarea unui număr de anomalii în dezvoltarea fătului și a organelor acestuia. În săptămâna 16-18 de sarcină folosind metoda de determinare a conținutului alfa-fetoproteinaîn serul sanguin al mamei pot fi detectate malformații ale sistemului nervos central (în cazul creșterii nivelului său de mai mult de 2 ori) sau anomalii cromozomiale, de exemplu, sindromul Down - trisomia cromozomului 21 sau
Tabelul 21.2. Momentul apariției unor anomalii în dezvoltarea embrionilor și a fetușilor umani

altă trisomie (aceasta este evidențiată printr-o scădere a nivelului substanței de testat de mai mult de 2 ori).
Amniocenteza- o metoda de cercetare invaziva prin care lichidul amniotic este prelevat prin peretele abdominal al mamei (de obicei in saptamana a 16-a de sarcina). În viitor, se efectuează o analiză cromozomială a celulelor lichidului amniotic și alte studii.
Monitorizarea vizuală a dezvoltării fetale este, de asemenea, utilizată folosind laparoscop, introdus prin peretele abdominal al mamei în cavitatea uterină (fetoscopie).
Există și alte modalități de a diagnostica anomaliile fetale. Cu toate acestea, sarcina principală a embriologiei medicale este de a preveni dezvoltarea lor. În acest scop, se dezvoltă metode de consiliere genetică și selecție a cuplurilor căsătorite.
Metode de inseminare artificiala celulele germinale de la donatori evident sănătoși fac posibilă evitarea moștenirii unui număr de trăsături nefavorabile. Dezvoltarea ingineriei genetice face posibilă corectarea leziunilor locale ale aparatului genetic al celulei. Deci, există o metodă, a cărei esență este obținerea unei biopsii testiculare din
bărbații cu o boală determinată genetic. Introducerea ADN-ului normal în spermatogonie și apoi transplantul spermatogoniei în testiculul iradiat anterior (pentru a distruge celulele germinale defecte genetic), reproducerea ulterioară a spermatogoniei transplantate duce la faptul că spermatozoizii nou formați sunt eliberați de defect determinat genetic. Prin urmare, astfel de celule pot produce descendenți normali atunci când o celulă reproductivă feminină este fertilizată.
Metoda de crioconservare a spermei vă permite să mențineți capacitatea de fertilizare a spermatozoizilor pentru o perioadă lungă de timp. Acesta este folosit pentru a păstra celulele germinale ale bărbaților asociate cu pericolul de expunere, rănire etc.
Metoda de inseminare artificială și transfer de embrioni(fertilizarea in vitro) este utilizată pentru a trata atât bărbații, cât și femeile infertilitate feminină. Laparoscopia este folosită pentru a obține celule germinale feminine. Un ac special este folosit pentru a perfora membrana ovarului în zona foliculului vezicular, pentru a aspira ovocitul, care este ulterior fertilizat de spermatozoizi. Cultivarea ulterioară, de regulă, până la stadiul de 2-4-8 blastomere și transferul embrionului în uter sau trompe uterine asigură dezvoltarea acestuia în condițiile organismului matern. În acest caz, este posibil să se transplanteze embrionul în uterul unei mame „surogat”.
Îmbunătățirea metodelor de tratament al infertilității și prevenirea anomaliilor dezvoltării umane sunt strâns legate de problemele morale, etice, juridice, sociale, a căror soluție depinde în mare măsură de tradițiile stabilite ale unui anumit popor. Acesta este subiectul unui studiu și discuții speciale în literatura de specialitate. În același timp, progresele în embriologia clinică și reproducerea nu pot afecta semnificativ creșterea populației din cauza costului ridicat al tratamentului și a dificultăților metodologice în lucrul cu celulele germinale. De aceea, la baza activităților care vizează îmbunătățirea sănătății și creșterea numerică a populației stă munca preventivă a unui medic, bazată pe cunoașterea proceselor de embriogeneză. Pentru nașterea de urmași sănătoși, este important să duceți un stil de viață sănătos și să abandonați obiceiuri proaste, precum și să desfășoare un ansamblu al acelor activități care sunt de competența instituțiilor medicale, publice și de învățământ.
Astfel, în urma studierii embriogenezei umane și a altor vertebrate, s-au stabilit principalele mecanisme de formare a celulelor germinale și fuziunea acestora cu apariția unui stadiu unicelular de dezvoltare, zigotul. Dezvoltarea ulterioară a embrionului, implantarea, formarea straturilor germinale și rudimentele embrionare ale țesuturilor, organelor extraembrionare arată o relație evolutivă strânsă și continuitate în dezvoltarea reprezentanților diferitelor clase ale lumii animale. Este important de știut că există perioade critice în dezvoltarea embrionului, când riscul de deces intrauterin sau de dezvoltare în funcție de condițiile patologice crește brusc.
cale. Cunoașterea proceselor obișnuite de bază ale embriogenezei face posibilă rezolvarea unui număr de probleme în embriologia medicală (prevenirea anomaliilor dezvoltării fetale, tratamentul infertilității), pentru a implementa un set de măsuri care previn moartea fetușilor și a nou-născuților.
Întrebări de control
1. Compoziția tisulară a copilului și a părților materne ale placentei.
2. Perioade critice ale dezvoltării umane.
3. Asemănări și diferențe în embriogeneza vertebratelor și a oamenilor.
4. Surse de dezvoltare tisulară a organelor provizorii.
Histologie, embriologie, citologie: manual / Yu. I. Afanasiev, N. A. Yurina, E. F. Kotovsky și alții - ed. a 6-a, revizuită. si suplimentare - 2012. - 800 p. : bolnav.
Selectie din baza de date: Doctrina bolii, etiologie generala, patogeneza generala Metodologica m, Vopr ex. Partea generala.docx, 1 Introducere Nosologie generala.doc, 2017-18 Entomologie generala.doc, teste de examen pentru cursul histologie, citologie, embriologi, Job general VMT.doc, Manual IGIENĂ GENERALĂ ŞI MILITARĂ.doc, energie generală practică.doc, 2 curs Etiologie generală şi patogeneză B E.ppt, Song why-why general.docx.Tipuri de blastule
celoblastula format cu strivire uniformă completă. Are un singur strat de blastoderm și o cavitate (blastocel) situată în centru. Lancelet.
Amfiblastula format în timpul zdrobirii complete neuniforme. Are un blastoderm multistratificat și un blastocel situat excentric. Amfibieni.
Discoblastula Se formează în timpul zdrobirii parțiale meroblastice și discul germinativ este turtit pe gălbenuș. Este format din blastomeri și corespunde acoperișului și zonei marginale. Gălbenușul corespunde fundului blastulei, iar decalajul îngust dintre ele este blastocelul. Păsări, reptile.
gastrulatie - perioada de embriogeneză, care este însoțită de formarea straturilor germinale: ectoderm, endoderm, mezoderm, iar embrionul însuși capătă o structură cu trei straturi.
Straturile germinale sunt dispuse in straturi: a) ectoderm - frunza exterioara; b) endoderm - frunza interioara; c) între ele se află notocordul și mezodermul;
Tipuri de gastrulatie
Procesul de gastrulare are loc în patru moduri principale:
Imigrarea - deplasarea unei părți din blastomerele peretelui în mijlocul embrionului și formarea stratului interior - endodermul;
Invaginare - procesul de indentare (invaginare) a unei părți a peretelui (inferioară) blastulei în mijloc;
Epiboly - procesul de murdărire prin divizarea rapidă a celulelor dintr-o secțiune a peretelui blastulei celulelor unei alte secțiuni, a căror diviziune are loc mai lent. Acest lucru se întâmplă atunci când blastomerele polului vegetativ au o cantitate mare de gălbenuș și se divid încet (amfibieni).
Delaminarea este un proces care este însoțit de o diviziune tangențială a peretelui blastulei și duce la formarea a două straturi: ectoderm primar- exterior; endoderm primar - in intern. caracteristic păsărilor și mamiferelor.
Gastrulația la om are loc în perioada de la 7 la 17 zile de ontogeneză prenatală și constă din 2 faze consecutive:
I faza decurge de la 7 la 14 zile si consta in formarea straturilor germinale externe (ectoderm) si interne (endoderm). Ca urmare delaminare un strat de celule este desprins de nodulul germinal, îndreptat spre cavitatea blastocistului - endodermul primar(hipoblast). În același timp, printre celulele nodulului germinal de sub hipoblast, un proces cavitație- din cauza acumulării de lichid în centrul nodulului, apare o cavitate, iar celulele din jurul acestuia capătă o formă epitelială (se formează o veziculă amniotică). Marginile opuse ale endodermului primar se pliază în jos și, crescând împreună, se formează veziculă de gălbenuș. Părțile ambelor vezicule adiacente una cu cealaltă (fundul amniotic și acoperișul gălbenușului) formează buricul embrionar sau discul embrionar (din această formațiune se formează corpul embrionului).
În paralel cu formarea acestor vezicule din nodulul germinativ, începând din ziua a 8-a, celulele sunt evacuate. mezodermul extraembrionar, care formează corionul și tulpina amniotică, care stă la baza viitorului cordon ombilical.
faza a II-a apare de la 15 la 17 zile, și constă în formarea mezodermului germinal. Se realizează prin imigrarea celulelor ectodermului primar în spațiul dintre două straturi germinale. Formarea sa are loc prin formarea unei îngroșări medii a benzii anterioare și a nodulului primar. Spațiul dintre straturile germinale este umplut cu țesut conjunctiv embrionar - mezenchim.
Sursa formării mezenchimatoase este mezodermul și, într-o măsură mai mare, ecto- și endodermul. Prin urmare, ei disting între endomezenchimul, care se dezvoltă din endomezoderm, și ectomezenchimul, care este de origine ectodermică.
Nu există diferențe morfologice între ele, dar dau naștere la structuri diferite:
a) entomezenchim - tesuturi ale mediului intern;
b) ectomezenchim - osiculele auditive, tesuturile conjunctive ale capului;
Evacuarea celulelor din zona nodulului primar duce la formarea unui șir axial al embrionului - acorduri. Pe peretele sacului vitelin la sfârșitul celei de-a doua săptămâni se formează insule de sânge și începuturile vaselor de sânge primare. Un proces asemănător unui deget crește în piciorul amniotic al părții posterioare a endodermului intestinal - alantois. Vasele sacului vitelin cresc în peretele alantoidei și vilozităților coriale, care sunt spălate de sângele matern. Formată ca urmare a acestor procese alantocorion asigură nutriția și respirația fătului în acest stadiu.
Histo-organogeneza
Histo-organogeneza- procesul de depunere și formare a țesuturilor organelor și sistemelor de organe în perioada embrionară ca urmare a unui număr de etape succesive: inducție, determinare, reproducere, migrare, creștere celulară, interacțiuni intercelulare și moarte celulară.
Inducţie - influența factorilor organizatori ai unor părți ale embrionului asupra altora, drept urmare a dezvoltare ulterioară organe și țesuturi.
Factorul de organizare (inductor) este un loc (punct) specific al embrionului, care afectează alte părți ale embrionului și determină direcția ulterioară a dezvoltării acestuia. Astfel de inductori pot fi proteine, nucleoproteine, steroizi. Factorii de organizare pot fi de ordinul I și II.
De exemplu: organizatorul, care este prezent în buza dorsală a blastoporului, induce o secțiune a ectodermului și provoacă diferențierea acestuia în placa neură. Acesta este organizatorul primei comenzi. La rândul său, în placa neurală apare un organizator de ordinul doi, care contribuie la transformarea unei secțiuni a tubului neural într-un ocular.
Determinare - determinarea căii ulterioare de dezvoltare a celulelor pe bază genetică datorită blocării componentelor individuale ale genomului.
Determinarea stă la baza proceselor de diferențiere și există 4 tipuri principale de diferențiere: 1) ootipică; 2) blastomeric; 3) rudimentar; 4) histogenetic;
Ootipic - când materialul sursă este reprezentat de zone prezumtive ale citoplasmei zigotului.
Blastomeric -în perioada blastulei.
Rudimentar- caracterizată prin apariția unor secțiuni separate ale straturilor germinale (stadiul gastrulației timpurii).
Histogenetic- caracterizat prin apariția unor rudimente ale diferitelor țesuturi în limitele unui singur strat germinativ.
reproducere celule. Reproducerea celulară se bazează pe diferite programe ale aparatului metabolic al celulei - activitate autosintetică și heterosintetică. La autosintetice metabolismul celular de activitate are ca scop îmbunătățirea proceselor de reproducere - o creștere a numărului de celule. heterosintetic- are ca scop formarea unor structuri specifice, sau sinteza si izolarea unor produse specifice.
Procesul de refacere a structurii unui obiect biologic după distrugerea acestuia se numește regenerare . În funcţie de nivelul de organizare, regenerarea poate fi: 1) celulară; 2) țesut; 3) organ.
În funcție de starea țesuturilor și organelor, regenerarea este împărțită în:
fiziologic, care apare constant într-un organism sănătos;
reparatorie, care apare după traumatizare.
Înălţime - procesul de formare, dezvoltare și organizare a celulelor animale sau umane ca urmare a unui număr de transformări complexe care au loc din momentul divizării la următoarea diviziune.
Interacțiunea celulară - procesele de interacţiune dintre celulele diferitelor primordii şi celulele lor de acelaşi tip ale aceluiaşi primordiu au un rol principal în diferenţierea progresivă a celulelor rudimentelor embrionare.
S-a stabilit că pentru dezvoltarea și existența normală a epiteliului pielii și a altor epitelii este necesar contactul constant sau temporar cu țesutul conjunctiv în curs de dezvoltare.
moartea celulelor - acesta este procesul de oprire ireversibilă a tuturor funcțiilor celulei și a conexiunii acesteia cu mediul.
Conceptul de organe provizorii. Structura.
Organele provizorii - organe temporare care se dezvoltă în procesul de embriogeneză, în afara corpului embrionului, și îndeplinesc funcții care asigură creșterea și dezvoltarea embrionului însuși.
Acestea includ: 1) sacul vitelin; 2) amnios; 3) membrana seroasa; 4) alantoida; 5) corion; 6) placenta.
Sacul vitelin- este o veziculă asociată tubului intestinal, al cărei perete este acoperit cu epiteliu în interior, iar în exterior este format din țesut conjunctiv.
Formarea ei are loc în stadiul gastrulei precoce, când pe stratul germinal intern se pot distinge endodermul germinal (intestinal) și ectodermul gălbenuș extraembrionar situat de-a lungul periferiei discului. După formarea pliului trunchiului, sacul vitelin rămâne legat de tubul intestinal prin tulpina vitelină.
¨Funcţii: a) trofice; b) hematopoietică (7-8 săptămâni);
Amnion- o înveliș continuă care este situată în jurul fătului și participă la producerea lichidului amniotic și constă din două părți amniotic cu fața la embrion și extern - seros.
Formarea amnionului are loc datorită ectodermului și stratului mezoderm parietal, care formează mai întâi pliuri amniotice care cresc spre suprafața dorsală a embrionului. Pliurile care cresc pe embrion sunt conectate, iar ambele foi - ectodermul și mezodermul parietal adiacente acestuia, cresc împreună cu foile cu același nume de pe partea opusă. Din două foi de pliuri se formează două membrane - amniotică (apă) și seroasă - externă.
Lichidul produs de celulele ectodermului membranei amniotice contine proteine, carbohidrati si asigura dezvoltarea libera a embrionului, deprecierea acestuia de eventuale deteriorari.
¨Funcții: asigurarea dezvoltării libere a embrionului, protecția împotriva eventualelor deteriorări mecanice și a influenței gravitației.
Membrana seroasă Se formează simultan cu amniotic, participă la furnizarea de oxigen a embrionului, ceea ce ne permite să-l considerăm ca un organ respirator provizoriu.
Allantoisîși începe dezvoltarea în partea caudală a embrionului însuși sub forma unei excrescențe a peretelui ventral al intestinului posterior, format din endoderm și foaia viscerală a mezodermului. Partea proximală a alantoidei este situată de-a lungul tulpinii gălbenușului, iar partea distală, în creștere, crește în golul dintre amnios și membrana seroasă. Acest organ de schimb și excreție gazoasă: prin vasele formate în mezodermul alantoidei se livrează oxigen; produsele metabolice sunt eliberate în alantoisă. Recent, s-a stabilit că în stadiile incipiente ale ontogenezei umane, alantois îndeplinește funcția de analog al pungii lui Fabricius, adică. îndeplinește funcția de organ central al limfocitopoiezei b. După 2 luni de embriogeneză, alantoida este redusă.
Chorion- membrana viloasa, se dezvolta din trofoblast si mezodermul extraembrionar. Inițial, trofoblastul este reprezentat de o membrană cu vilozități primare, prin care, după implantarea embrionului, se stabilește o legătură cu corpul mamei. La 2-3 săptămâni de dezvoltare apare un mezoderm extraembrionar care germinează la trofoblast și împreună cu acesta formează vilozități epiteliomezenchimatoase secundare. După aceea, trofoblastul se transformă într-un corion sau membrană viloasă.
Pătrunzând în membrana mucoasă a uterului, corionul se formează odată cu acesta placenta.
placenta - Acesta este un organ care asigură o legătură permanentă între făt și corpul mamei.
¨Dezvoltarea placentei începe în a 3-a săptămână, când vasele încep să crească în vilozitățile epiteliomezenchimale secundare și se formează vilozități terțiare. În viitor (săptămânile 6-8), macrofagele, fibroblastele și fibrele de colagen se diferențiază în jurul vaselor. Formarea fibrelor de colagen în vilozități coincide cu o creștere a activității proteolitice a epiteliului trofoblastic (citotrofoblast) și a derivatului său (sincitiotrofoblast).
Odată cu dezvoltarea placentei, se produce distrugerea mucoasei uterine și schimbarea nutriției histiotrofice în hepatotrofă. Aceasta înseamnă că vilozitățile corionului sunt spălate de sângele mamei, care s-a revărsat din vasele distruse ale endometrului în lacune.
Placenta este formată din 2 părți: 1) maternă; 2) fetală (embrionară).
Partea maternă a placentei format de membrana mucoasă a uterului în zona în care vilozitățile corionului fetal cresc în ea, aceasta este așa-numita carcasă principală de drop-off(deciduală). În plus față de această membrană, în compoziția endometrului uterului femeilor însărcinate, ele disting vilozități coriale fără creștere internă - carcasă de cădere lângă perete, și carcasă de picătură de sac, care separă embrionul de cavitatea uterină.
Partea fetală a placentei format din corionul vilos.
Există două tipuri de corion: a) corion ramificat; b) corion neted;
corion ramificat, ale căror vilozități cresc în endometru la locul deciduei.
Corion neted- locul de contact al trofoblastului cu teaca bursală.
Procesul de formare a placentei umane are loc cel mai intens între 3 și 6 săptămâni de embriogeneză.
În funcție de structura placentei mamiferelor, se disting 4 tipuri:
1) epiteliocorial; 2) desmocorial; 3) endoteliocorial; 4) hemocorial;
Placenta umană aparține placentei viloase hemocoriale discoide.
Unitatea structurală a placentei este cotiledon- aceasta corespunde ramificării unei vilozități tulpinii. Tulpina sau vilozitatea ancoră este o excrescere mare a plăcii coriale, strâns îmbinată cu decidua, de pe a cărei suprafață laterală se extind numeroase ramuri de plăci coriale mici.
Placenta umană are aproximativ 200 de cotiledoane, care sunt delimitate unul de celălalt prin septuri de țesut conjunctiv - septe prin care trec vasele arteriale, care transportă sângele îmbogățit cu oxigen și substanțe nutritive către lacunele placentei. Venele lacunare se deschid în lacune cu deschideri largi, prin care sângele matern curge din placentă.
Peretele lacunei este format din țesutul conjunctiv al endometrului, care este acoperit cu straturi dintr-o substanță amorfă - Fobrinoidul lui Rohr.
Acea parte a deciduei, care este situată între corionul ramificat și neted, care crește strâns împreună cu acesta, formează așa-numitul placă de capăt care împiedică curgerea sângelui în cavitatea uterină.
Bariera hemoplacentară este o barieră care separă sângele mamei de sângele fătului. Este format din epiteliocite și membrana bazală a hemocapilarelor vilozităților coriale, îmbogățită cu macrofage și fibroblaste, țesutul conjunctiv care înconjoară microcirculația, membrana bazală a vilozităților coriale, stratul sincitiotrofoblast și, de asemenea, situat pe suprafața ultimul fibrinoid Langhans.
Funcțiile placentei: 1. trofic; 2. excretor; 3. depunerea; 4. endocrin; 5. respirator; 6. protectoare.
Cordon ombilical- un cordon format din tesut conjunctiv, in care trec principalele vase - doua artere si o vena, asigurand circulatia sangelui intre fat si placenta. Cordonul ombilical include și resturile tulpinii de gălbenuș, alantoida.
Cordonul ombilical se bazează pe țesut conjunctiv mucos - așa-numitul B jeleu de arton, care conține o cantitate mare de acid hialuronic. Acesta din urmă asigură turgența cordonului ombilical. Dintre elementele celulare din compoziția cordonului ombilical, s-au găsit bazofile tisulare și celule Kashchenko-Hofbauer, care protejează fătul de infecția intrauterină.
PERIOADE CRITICE DE DEZVOLTARE
Perioade critice de dezvoltare- sunt perioadele de cea mai mare sensibilitate a celulelor germinale in curs de dezvoltare (progeneza) si a embrionului (embriogeneza) la actiunea factorilor nefavorabili.
Embriolog domestic Svetlov P.G. în 1960 a fost formulată şi demonstrată experimental teoria perioadelor critice de dezvoltare. Esența acestei teorii constă în faptul că fiecare etapă a dezvoltării embrionului în ansamblu și a organelor sale individuale începe cu o perioadă relativ scurtă de restructurare calitativ nouă, însoțită de determinarea, proliferarea și diferențierea celulelor. În această perioadă, embrionul este cel mai sensibil la acțiunea agenților dăunători.
Aceste perioade sunt: progeneza- spermatogeneza si ovogeneza; embriogeneza- fertilizarea, implantarea (6-8 zile), placentarea și dezvoltarea primordiilor axiale (săptămâna a 3-a, a 8-a), perioada de dezvoltare sporită a creierului (15-20 săptămâni), perioada de formare a principalelor sisteme funcționale ale organism (20-24 săptămâni), procesul de naștere. ontogeneza postnatală- perioada nou-născuților (până la 1 an), perioada pubertății (11-16 ani).
ȚESUTURI scheletice.
Țesuturile scheletice(textus sceletales) sunt țesuturi cartilaginoase și osoase care participă la metabolismul apă-sare și îndeplinesc funcții de susținere, de protecție și mecanice.
țesuturile cartilajului
țesuturile cartilajului(teхtus cartilaginei) reprezintă singurul tip de țesut în care nu există vase și fac parte dintr-un număr de organe ale sistemului respirator, articulații, discuri intervertebrale. Ele constau din celule (condrocite și condroblaste) și o cantitate mare de substanță intercelulară.
Cartilajul conține aproximativ 75% apă, 10 - 15% materie organicăşi 4-7% săruri anorganice. Substanța uscată conține 50 până la 70% colagen.
Clasificarea țesuturilor cartilajului
În funcție de structura și caracteristicile structurale și funcționale ale substanței intercelulare, se disting 3 tipuri de țesut cartilaginos:
1. Cartilajul hialin.
2. Cartilaj elastic.
3. Cartilaj fibros.
¨Funcții. Țesutul cartilajului din organism îndeplinește funcții de susținere și modelare.
Histogenia țesutului cartilajului
Dezvoltarea țesutului cartilajului se realizează în perioada embrionară și în perioada postembrionară în timpul regenerării.
Histogeneza embrionară
Sursa dezvoltării țesutului cartilaginos în ontogenie este mezenchimul - țesutul conjunctiv embrionar.
Histogenia cartilajului are loc în 3 etape:
¨ Primul stagiu -formarea unui rudiment condrogen sau a insulei condrogene.În acele locuri în care se formează cartilaj la început, are loc o compactare a mezenchimului, celulele acestuia își pierd procesele, reproducerea lor se intensifică și sunt strâns adiacente între ele, formează o anumită tensiune - turgor. Se numesc astfel de zone rudimente condrogenice sau insulițe condrogenice. Celulele mezenchimale, care fac parte din insula condrogenă, se diferențiază în condroblaste, celule care dau naștere la formarea țesutului cartilajului. În citoplasma unor astfel de celule, numărul de ribozomi liberi crește, apar zone ale reticulului endoplasmatic granular.
¨ A doua faza- formarea cartilajului primar. Are loc o rotunjire a celulelor zonei centrale, o creștere a dimensiunii lor, în citoplasmă se dezvoltă un reticul endoplasmatic granular, cu ajutorul căruia sunt sintetizate și secretate proteine fibrilare (colagen de tip III). Ca urmare a acestor modificări, condroblastele se transformă în condrocite primare. Substanța intercelulară astfel formată se distinge prin oxifilie.
¨ A treia etapă- diferențierea cartilajului. În această etapă, condrocitele primare se transformă în condrocite secundare și dobândesc capacitatea de a sintetiza, pe lângă substanțele enumerate mai sus, glicozaminoglicani sulfatați (sulfați de condroitin) asociați cu proteine de colagen (proteoglicani).
La periferia anlagei cartilaginoase, la limita cu mezenchimul, a pericondriu- o teaca care acopera exteriorul cartilajului in curs de dezvoltare, formata din exterior fibrosȘi condrogen intern straturi (cambiale).
Creșterea cartilajului poate avea loc în două moduri:
¨ Prin suprapunere (creștere apozițională), în timp ce în zona condrogenă, celulele se divid intens, se diferențiază condroblasteîn condrocite, care produc substanță intercelulară, există o stratificare de celule nou formate pe cartilajul deja existent la periferie. În procesul de secreție a substanței intercelulare, aceste celule se imunează în ea.
¨ Prin intern (interstitial) creştere, celulele cartilajului situate în centrul unui cartilaj tânăr în curs de dezvoltare își păstrează capacitatea de a se diviza mitotic pentru o perioadă de timp, rămânând într-un singur gol (grupuri izogenice de celule). Prin creșterea numărului acestor celule, are loc o creștere a masei cartilajului din interior. O astfel de creștere se observă în embriogeneză, precum și în timpul regenerării țesutului cartilajului.
Regenerare fiziologică - apare datorita activitatii condrocitelor, producerii lor de substanta condromucoid, colagen si elastina, care contribuie la neoplasmul fibrelor condrinice.
Pe măsură ce cartilajul crește și se dezvoltă, secțiunile sale centrale sunt din ce în ce mai separate de vase și întâmpină dificultăți în alimentație, efectuate difuz din pericondriu. Prin urmare, condrocitele își pierd capacitatea de a se reproduce, unele sunt distruse, iar proteoglicanii se transformă într-o proteină mai simplă - albumoid.
Celulele cartilajului
Condroblaste - celule tinere nediferențiate capabile de proliferare și sinteza de substanță intercelulară.
Formă- neregulat, alungit, turtit.
Dezvoltare- din semi-celule stem (perechondroblast), care provin din celule stem. Celulele stem, semi-celulele stem, condroblastele și condrocitele formează o serie histogenetică.
Citoplasma- contine un reticul endoplasmatic (granular si agranular) bine dezvoltat si elemente ale complexului Golgi, mult ARN. Colorat bazofil.
În timpul dezvoltării cartilajului, condroblastele se transformă în condrocite. Condroblastele realizează creșterea cartilajului periferic (apozițional).
Condrocite - principalele celule ale țesutului cartilajului.
Formă- oval, rotund sau poligonal.
Localizare- localizate în cavităţi speciale ale substanţei intercelulare (lacune). Aceste grupuri de celule se numesc (izogenice).
petrecându-se din cauza diviziunii unei celule. Există trei tipuri de condrocite în grupul izogen:
eu tipul de celule predomină în cartilajele tinere în curs de dezvoltare; diviziunea este adesea observată în aceste celule, ceea ce ne permite să le considerăm ca o sursă de reproducere a grupurilor izogenice.
caracteristică pentru aceste celule este prezența unui indice nuclear – citoplasmatic ridicat.
Citoplasma- are elemente vacuolare bine dezvoltate, complex lamelar, mitocondrii si ribozomi liberi.
II tipul celular - caracterizat prin scăderea indicelui nuclear - citoplasmatic, slăbirea sintezei ADN-ului, dar sinteza ARN este crescută, reticulul endoplasmatic granular, complexul Golgi, care asigură formarea și secreția de glicozaminoglicani și proteoglicani în substanța intercelulară, este dezvoltat intens. Citolema și caryolema sunt de obicei sinuoase.
III tip de condrocite. Aceste celule se caracterizează printr-un indice nuclear - citoplasmatic scăzut, o dezvoltare puternică și o aranjare ordonată a reticulului endoplasmatic granular. Acest tip de celule își păstrează capacitatea de a forma și secreta proteine, reducând în același timp sinteza glicozaminoglicanilor.
substanta intercelulara a cartilajului reprezentată de o componentă organică - proteine, lipide, glicozaminoglicani și proteoglicani. Concentrația de proteoglicani în acest țesut este cea mai mare. Proteinele fibrilare, în principal colagenul de tip II, sunt conținute în cantități mari.
Orientarea fibrelor determinată de direcția liniilor de forță.
Stratul de substanță intercelulară adiacent cavității celulare și care formează peretele acesteia este caracterizat de o refracție ridicată a luminii și conține o rețea de fibrile care formează pâslă. Uneori este numită capsula celulară a cartilajului.
Cartilajul hialin.
Localizare- în peretele traheei, bronhiilor, la joncțiunea coastelor și sternului, suprafețe articulare și în plăcile metaepifizare.
Structura. Cartilajul hialin este acoperit la exterior pericondriu(pericondru).
Pericondrul este format din două straturi: 1) exterior; 2) intern;
* Extern - format din tesut conjunctiv fibros cu vase de sange.
* Intern - format în principal din celule precondroblaste și condroblaste.
Sub pericondriu, în stratul de suprafață al cartilajului însuși, există tineri condrociteîn formă de fus, a cărui axă lungă este îndreptată de-a lungul suprafeței cartilajului.
În straturile mai adânci, condrocitele capătă o formă ovală și rotunjită, dispuse în mai multe grupe, formând grupări izogenice. Condrocitele tinere și grupurile izogenice sunt înconjurate de fibre condromucoide și de colagen (colagen tip II).
· Cu toate acestea, nu toate cartilajele hialine au aceeași structură.
Cartilajul hialin al suprafeței articulare nu are pericondriu pe suprafața orientată spre interiorul articulației. Cartilajul articular este format din trei zone care nu sunt clar definite: a) extern; b) medie; in adancul;
În zona exterioară există celule mici nespecializate turtite.
În mijloc- celulele sunt mai mari, ovale, rotunjite, dispuse in coloane perpendiculare pe suprafata.
zona profundă compus din cartilaj calcificat; numai în această zonă s-au găsit vase de sânge.
Țesut elastic de cartilaj
Acest țesut este localizat în acele structuri care sunt supuse îndoirii - acestea sunt: auriculul, roșcovul și cartilajele sfenoide ale laringelui.
*Structura.- Conform planului general al structurii, cartilajul elastic este asemanator hialinului. În exterior, este acoperit cu un pericondriu. Celulele cartilajului sunt situate în capsule individual sau formează grupuri izogenice.
Spre deosebire de cartilajul hialin, substanta intercelulara contine, alaturi de fibrele de colagen, fibre elastice formate din proteina elastina. Ele pătrund în substanța intercelulară în toate direcțiile. În straturile adiacente pericondrului, fibrele elastice trec fără întrerupere în fibrele elastice ale pericondrului.
Există mai puține lipide, glicogen și sulfați de condroetin în cartilajul elastic decât în hialin. În plus, calcificarea nu apare niciodată în cartilajul elastic.
Țesut cartilaj fibros
Localizareîntre vertebrele discurilor, articulații semi-mobile, în locurile în care țesutul conjunctiv fibros (tendoane, ligamente) trece în cartilajul hialin și unde restricția mișcării este însoțită de o tensiune puternică.
Structura- substanta intercelulara contine fascicule de colagen indreptate paralel, care treptat se desfac si trec in cartilajul hialin. Condrocitele din fibrocartilaj sunt aranjate sub formă de rânduri deosebite - coloane.
Citoplasma celulele sunt adesea vacuolate. În direcția de la cartilajul hialin la tendon, fibrocartilajul devine mai mult ca un tendon. Pe marginea cartilajului și a tendonului, în loc de coloane de celule tendinoase, între fasciculele de colagen lipite în substanța fundamentală, se află coloane de celule de cartilaj comprimat, care, fără nicio margine, trec în adevărate celule de tendon situate în țesutul conjunctiv dens.
Țesut osos, caracteristici generale.
Os(textus osseus) este un tip specializat de țesut conjunctiv care are un grad ridicat de mineralizare a substanței intercelulare.
Țesutul osos este format din elemente celulare (osteoblaste, osteocite și osteoclaste) și substanță intercelulară (oseină și osteomucoid).
Substanța intercelulară conține aproximativ 70% compuși anorganici, în principal fosfați de calciu. Compușii organici sunt reprezentați în principal de proteine și lipide care alcătuiesc matricea. Compușii organici și anorganici în combinație dau un țesut de susținere foarte puternic.
Funcții
musculo-scheletice- datorita rezistentei semnificative a tesutului osos, asigura miscarea corpului in spatiu si sustinerea acestuia.
3. depozit calciu și fosfor în organism;
Clasificarea țesutului osos
În funcţie de structură şi proprietăți fizice Există două tipuri de țesut osos:
1. Reticulofibros (fibros grosier)
2. Farfurie
Reticular - țesut osos fibros- prezinta o dispunere multidirectionala de fascicule de fibre de osseina (colagen tip I), inconjurate de osteomucoid calcificat. Osteocitele se află între fasciculele de fibre de osseină din lacunele osteomucoidului. Acest țesut este caracteristic scheletului fetal; la adulți, se găsește numai în zonele suturilor craniului și în locurile în care tendoanele sunt atașate de oase.
țesut osos lamelar- caracteristică este o aranjare strict paralelă a mănunchiurilor de fibre de colagen și formarea plăcilor osoase.
În funcție de orientarea acestor plăci în spațiu, acest țesut, la rândul său, se împarte în: 1) compact; 2) spongios;
compact- caracterizat prin absenţa cavităţilor. Din ea sunt construite diafizele oaselor tubulare.
Spongios- caracterizata prin faptul ca placile osoase formeaza trabecule situate in unghi una fata de alta. Ca rezultat, se formează o structură spongioasă. Țesutul osos spongios formează oase plate epifize ale oaselor tubulare.
Histogeneza osoasa
Sursa dezvoltării țesutului osos este mezenchimul. Odată cu dezvoltarea țesutului osos, se formează două diferențe de celule (serie histogenetică).
¨ Primul rand- celule stem osteogene, celule semi-stem stromale, osteoblaste, osteocite.
¨ Al doilea rând- origine hematogena - celula stem hematopoietica, celula semistem hematopoietica (predecesorul celulelor mieloide si macrofagelor), celula monocitara formatoare de colonii unipotente (monoblast), promonocit, monocit, osteoclaste (macrofage).
Distingeți între dezvoltarea embrionară și postembrionară a țesutului osos.
Embrionară Dezvoltarea osoasă poate avea loc în două moduri:
1. Direct din mezenchim – osteohistogeneză directă.
2. Din mezenchim în locul modelului de os cartilaginos dezvoltat anterior, osteohistogeneză indirectă.
Dezvoltare postembrionară osul se efectuează în timpul regenerării și osteogenezei ectopice.
Gastrulația este o perioadă de dezvoltare embrionară în care are loc reproducerea, creșterea și mișcarea celulelor individuale și a straturilor celulare extinse. Principala diferență între gastrulație și perioadele anterioare de embriogeneză este dobândirea de către celule a capacității de mișcări morfogenetice direcționate, care duc la o restructurare profundă a embrionului. Dacă rezultatul zdrobirii a fost formarea multicelularității, atunci gastrulația duce la formarea unui embrion multistrat.
Mișcările morfogenetice ale celulelor variază foarte mult în diferite clase de animale. Luați în considerare principalele tipuri de gastrulație:
1) Invaginarea este invaginarea unui perete al blastulei în blastocel.Această metodă de gastrulare este caracteristică lanceletei.
2) Epiboly - murdărire cu celule mici care se divizează rapid de celule mari, cu diviziune lentă, supraîncărcate cu gălbenuș și, prin urmare, nu arată capacitatea de mișcare. Această metodă de gastrulare este observată la amfibieni.
3) Delaminare - scindarea blastodermei în două straturi. Cu această metodă de gastrulare, deplasările celulare sunt aproape absente. Elementele de delaminare se găsesc în timpul gastrulației peștilor, păsărilor și mamiferelor.
4) Imigrarea - evacuarea activă a unei părți a celulelor peretelui blastulei în blastocel. Când celulele sunt evacuate dintr-un singur pol al blastulei, se vorbește de imigrare unipolară, din doi poli - bipolar, iar când celulele sunt evacuate de pe întreaga suprafață a embrionului - multipolar. Este, de asemenea, un mod destul de comun de gastrulație și se găsește la multe vertebrate (pești, păsări, mamifere).
Există tipuri mixte de gastrulație. În general, tipurile considerate de gastrulație sunt într-o anumită măsură condiționate și, în majoritatea cazurilor, este mai corect să spunem că unul dintre tipuri este dominant în natura mișcărilor morfogenetice, iar mai multe dintre aceste tipuri pot apărea simultan în proces. de gastrulaţie la multe animale.
Mecanisme de gastrulatie.
Oricât de diverse sunt tipurile de gastrulație, există modificări comune la nivel celular care duc la mișcări morfogenetice. Majoritatea proceselor de modelare se bazează pe diviziunile celulare, apariția unor solicitări mecanice în rezervor și apoi alternanța polarizării (întinderea) și contracția celulară.
Polarizarea celulară este redistribuirea organelelor cu activarea rapidă a sintezei actinei, asamblarea fasciculelor de microtubuli și alungirea celulei în direcția mișcării viitoare. În acest caz, se formează așa-numita celulă în formă de balon. Este caracteristic că o astfel de polarizare afectează nu o celulă, ci întregul strat de celule: i.e. polarizarea unei celule induce următoarea transformare. Acest proces se realizează numai în prezența contactelor celulare și, prin urmare, se numește polarizare a celulei de contact.
Celulele nu pot rămâne într-o stare de polarizare la infinit: după un anumit timp, are loc contracția - o astfel de deformare a unei celule polarizate care reduce raportul dintre suprafața acesteia și volum. Acest proces este realizat de aparatul contractil al celulei - microfilamente. În cursul contracțiilor succesive, stratul se îndoaie și are loc o mișcare morfogenetică elementară.
Astfel, apariția într-o anumită regiune din cauza condițiilor externe speciale și a efectelor diviziunii celulare creează solicitări mecanice în rezervor. Aceste tensiuni duc la apariția polarizării celulare, care poartă „informații” despre direcția mișcării viitoare, iar implementarea ei (adică mișcarea reală) are loc în timpul contracției.
Procesele celulare care stau la baza proceselor formative ale dezvoltării timpurii, coordonarea lor în timp și spațiu și posibile alte cauze ale mecanismelor surprinzător de complexe și ordonate ale gastrulației sunt încă neclare.
Pentru comoditatea luării în considerare a cursului gastrulației și a rezultatelor sale la vertebrate, întregul proces este împărțit condiționat în două etape: gastrulație timpurie și târzie.
În timpul gastrulației timpurii, inițial un singur strat de celule blastule, reorganizându-se în oricare dintre modurile de mai sus, formează două straturi. Stratul exterior de celule se numește ectoderm, iar stratul interior se numește endoderm. La vertebratele inferioare, se formează o nouă cavitate - gastrocoelul. Deschiderea care duce spre exterior din gastrocoel se numește blastopor (gura primară), iar marginile sale sunt numite buze.
Materialul buzei dorsale a blastoporului la diferite specii de animale este transformat ulterior într-o notocordă, iar materialul lateral - în al treilea strat germinativ - mezodermul. Prin urmare, pentru a înțelege mișcările morfogenetice ale blastoporului, este un punct de referință destul de important. Soarta blastoporului variază de la animal la animal. La unele (primare-stome), blastoporul, dezvoltându-se și diferențiându-se în consecință, se transformă într-o gură definitivă, în altele (secundar-stome), blastoporul se transformă în anus. La vertebratele superioare (păsări, mamifere), nu formează un blastopor în timpul imigrării. Pentru orientarea în viitoarele moduri de dezvoltare a embrionului, nu putem vorbi decât de un analog al blastoporului.
Astfel, ca urmare a gastrulației timpurii, se formează un embrion cu două straturi și un blastopor, iar la mamifere, în plus, se formează și unele organe extraembrionare.
Odată cu gastrulația târzie, se formează un al treilea strat germinal - mezodermul, un complex de organe osoase și organe extraembrionare.
Embriologia clasică descrie două moduri de formare a mezodermului: enteroceloasă și teloblastică. Cu metoda enterocelului, mezodermul se formează ca o colecție de celule care s-au separat de intestinul primar, iar cu metoda teloblastului, mezodermul se formează din celule situate la capătul posterior al viitorului embrion la limita ecto-ului. și endodermul.
Organele axiale sunt notocorda, tubul neural și intestinul primar. Primul material al buzei dorsale a blastoporului este notocorda, un fir de celule dens situat de-a lungul liniei mediane a embrionului între ecto- și endoderm. Sub influența sa, tubul neural începe să se formeze în stratul germinal exterior. Și în sfârșit, endodermul formează intestinul primar.
Formarea tubului neural este direct legată de neurulație - așezarea sistemului nervos central. Neurulația este o perioadă foarte importantă și interesantă în dezvoltarea embrionului, nu numai pentru că este pus un sistem complex,
dar și pentru că în timpul formării tubului neural,
cea mai apropiată interacțiune între structurile adiacente: ectoderm, coardă și mezoderm. Trebuie subliniat că una dintre principalele consecințe ale mișcărilor morfogenetice este că grupurile de celule care anterior puteau fi semnificativ îndepărtate unele de altele sunt atât de apropiate încât interacțiunile dintre ele devin posibile, care se numesc inducție. Neurulația, în special formarea tubului neural, este rezultatul unor astfel de interacțiuni inductive.
Embriologie generală și comparată
Plan
1. Caracteristicile morfofuncționale ale celulelor germinale masculine.
2. Tipuri de ouă în funcție de numărul și plasarea gălbenușului. Structura și funcția oului.
3. Fertilizarea, conceptul fazelor sale îndepărtate și de contact.
4. Definiția zdrobirii și tipurile sale.
5. Gastrulare, metode de gastrulatie precoce si tardiva.
6. Organe extraembrionare ale vertebratelor (amnios, sacul vitelin, corion, alantois, cordon ombilical, placenta).
7. Placenta, tipuri de placente după structura, forma și metoda de hrănire a fătului.
8. .Conceptul de fertilizare in vitro și semnificația acestuia.
9. Placenta umană, caracteristicile și semnificațiile sale morfologice.
10. Structura placentei.
11. Componentele structurale ale barierei hemocoriale (placentare).
12. Sistemul mamă-făt.
13. Conceptul de perioade critice de dezvoltare.
În complexul științelor medicale, embriologia ocupă unul dintre locurile proeminente. Cunoașterea embriologiei este necesară pentru a înțelege principalele modele de dezvoltare intrauterină și caracteristicile sale specifice la diferiți reprezentanți ai regnului animal în legătură cu conditii diferite vieţile lor şi originile specifice. Cunoașterea elementelor de bază ale embriologiei comparate ajută la înțelegerea modelelor biologice generale ale evoluției vertebratelor, a condiționalității filogenetice a proceselor de formare a corpului uman și, de asemenea, la înțelegerea elementelor de bază ale ingineriei genetice. În același timp, este important despre înțelegerea consecințelor influența diverșilor factori de mediu nefavorabili asupra embriogenezei reprezentanților diferitelor specii.
Cunoașterea embriologiei este necesară pentru viitorul medic pentru prevenirea rațională a anomaliilor și malformațiilor, precum și pentru prevenirea efectelor adverse ale factorilor nocivi de mediu și de zi cu zi asupra cursului sarcinii. Studiul embriologiei umane este rațiunea științifică pentru astfel de discipline precum obstetrică, ginecologie și pediatrie. Cunoașterea etapelor incipiente ale embriogenezei umane face posibilă corectarea proceselor de formare și dezvoltare a celulelor germinale primare, determinarea cauzelor gametopatiilor, prevenirea infertilității și, de asemenea, determinarea etapelor clivajului embrionului, cauzele gemenilor identici, determinarea momentul și etapele de implantare, care sunt necesare în cazul dezvoltării extracorporale a embrionului.
Embriologie- știința formării și dezvoltării embrionului.
Embriologie generală - studiază cele mai generale modele de formare și dezvoltare a embrionului.
Embriologie specială - studiază trăsăturile dezvoltării individuale a reprezentanților anumitor grupuri sau specii.
Embriologie , știința care studiază dezvoltarea unui organism în stadiile sale incipiente, înainte de metamorfoză, ecloziune sau naștere. Fuziunea gameților - un ou și un spermatozoid - cu formarea unui zigot dă naștere unui nou individ, dar înainte de a deveni aceeași creatură cu părinții săi, acesta trebuie să treacă prin anumite etape de dezvoltare: diviziunea celulară, formarea straturile și cavitățile germinale primare, apariția axelor embrionare și a axelor de simetrie, dezvoltarea cavităților celomice și a derivaților acestora, formarea membranelor extraembrionare și, în final, apariția unor sisteme de organe care sunt integrate funcțional și formează unul sau altul recunoscut. organism. Toate acestea fac obiectul studiului embriologiei.
Procese și etape embriogeneza
1. Fertilizarea
2. Zdrobire
3. Gastrulare
4. Neurulație
5. Histogenie
6. Organogeneza
7. Sistemogeneza
Dezvoltarea este precedată de gametogeneză, adică. formarea si maturarea spermatozoizilor si ovulelor. Procesul de dezvoltare a tuturor ouălor unei specii date decurge în general în același mod.
Gametogeneza. Spermatozoizii maturi și ouăle diferă în structura lor, doar nucleii lor sunt similari; cu toate acestea, ambii gameți sunt formați din celule germinale primordiale cu aspect identic. În toate organismele cu reproducere sexuală, aceste celule germinale primare se separă de alte celule în stadiile incipiente de dezvoltare și se dezvoltă într-un mod special, pregătindu-se să-și îndeplinească funcția - producția de celule sexuale sau germinale. Prin urmare, ele sunt numite plasmă germinativă - spre deosebire de toate celelalte celule care alcătuiesc somatoplasma. Este destul de evident, însă, că atât germoplasma, cât și somatoplasma provin dintr-un ou fecundat - un zigot care a dat naștere unui nou organism. Deci practic sunt la fel. Factorii care determină care celule vor deveni sexuale și care vor deveni somatice nu au fost încă stabiliți. Cu toate acestea, în cele din urmă, celulele germinale capătă diferențe destul de clare. Aceste diferențe apar în procesul de gametogeneză.
Celulele germinale primare, aflându-se în gonade, se divid odată cu formarea de celule mici - spermatogonie în testicule și oogonie în ovare. Spermatogonia și oogonia continuă să se dividă de multe ori, formând celule de aceeași dimensiune, ceea ce indică creșterea compensatorie atât a citoplasmei, cât și a nucleului. Spermatogonia și oogonia se divid mitotic și, prin urmare, își păstrează numărul diploid inițial de cromozomi.
După ceva timp, aceste celule încetează să se divizeze și intră într-o perioadă de creștere, în care apar modificări foarte importante în nucleele lor. Cromozomii primiți inițial de la doi părinți sunt împerecheați (conjugați), intrând în contact foarte strâns. Acest lucru face posibilă încrucișarea ulterioară (încrucișarea), în timpul căreia cromozomii omologi sunt rupți și conectați într-o nouă ordine, schimbând secțiuni echivalente; ca urmare a încrucișării, apar noi combinații de gene în cromozomii oogoniei și spermatogoniei.
Când nucleul a fost reconstruit și s-a acumulat o cantitate suficientă de citoplasmă în celulă, procesul de diviziune se reia; intreaga celula si nucleul sufera doua tipuri diferite de diviziuni, care determina procesul propriu-zis de maturare a celulelor germinale. Una dintre ele - mitoza - duce la formarea de celule similare cu originalul; ca urmare a celuilalt - meioza sau diviziunea de reducere, în timpul căreia celulele se divid de două ori, se formează celule, fiecare dintre ele conține doar jumătate (haploid) din numărul de cromozomi față de originalul, și anume, unul din fiecare pereche. La unele specii, aceste diviziuni celulare au loc în ordine inversă. După creșterea și reorganizarea nucleilor în oogonii și spermatogonii și imediat înainte de prima diviziune a meiozei, aceste celule se numesc ovocite și spermatocite de ordinul întâi, iar după prima diviziune a meiozei, ovocite și spermatocite de ordinul doi. În cele din urmă, după a doua diviziune a meiozei, celulele din ovar sunt numite ouă (ouă), iar cele din testicul sunt numite spermatide. Acum, ovulul s-a maturizat în sfârșit, iar spermida nu a trecut încă prin metamorfoză și nu se transformă într-un spermatozoid.
Rolul biologic al spermatozoizilor în procesul de fertilizare
1. Oferă o întâlnire cu ovocitul.
2. Oferă 23 de cromozomi parentali.
3. Determină sexul copilului.
4. Introduce un centrol în ovocit.
5. Oferă ADN mitocondrial.
6. Provoacă terminarea meiozei de către ovul.
7. Introduce o proteină semnal de clivaj.
O diferență importantă între oogeneză și spermatogeneză trebuie subliniată aici. Dintr-un ovocit de ordinul întâi, ca urmare a maturării, se obține un singur ou matur; restul de trei nuclei și o cantitate mică de citoplasmă se transformă în corpuri polari care nu funcționează ca celule germinale și ulterior degenerează. Toată citoplasma și gălbenușul, care ar putea fi distribuite pe patru celule, sunt concentrate într-una singură - într-un ou matur. În schimb, un spermatocit de ordinul întâi dă naștere la patru spermatide și același număr de spermatozoizi maturi, fără a pierde un singur nucleu. În timpul fertilizării, numărul diploid, sau normal, de cromozomi este restabilit.
Ou. Ovulul este inert și de obicei mai mare decât celulele somatice ale organismului. Oul de șoarece are aproximativ 0,06 mm în diametru, în timp ce diametrul oului de struț este mai mare de 15 cm. Ouăle sunt de obicei de formă sferică sau ovală, dar pot fi și alungite. Mărimea și alte caracteristici ale oului depind de cantitatea și distribuția gălbenușului nutritiv din acesta, care se acumulează sub formă de granule sau, mai rar, sub forma unei mase continue. Prin urmare, ouăle sunt împărțite în diferite tipuri, în funcție de conținutul de gălbenuș din ele. În ovocitele homolecitale, numite și isolecital sau oligolecital, există foarte puțin gălbenuș și este distribuit uniform în citoplasmă.
Sperma. Spre deosebire de un ou mare și inert, spermatozoizii sunt mici, de la 0,02 la 2,0 mm lungime, sunt activi și capabili să parcurgă o distanță lungă pentru a ajunge la ovul. Există puțină citoplasmă în ele și nu există deloc gălbenuș.
Forma spermatozoizilor este diversă, dar printre ele se pot distinge două tipuri principale - flagelate și neflagelate. Formele flagelate sunt relativ rare. La majoritatea animalelor, un rol activ în fertilizare îi revine spermatozoidului.
Fertilizare- fuziunea celulelor sexuale. Semnificație biologică: reluarea diploȘi un set de cromozomi; determinarea sexului copilului; iniţierea zdrobirii. Faze: d instantna (capacitate si i, taxiuri); contact (acrozomal eu reacție, denudare si eu, penetrac si i, reacție corticală)
Fertilizare. Fertilizarea este un proces complex în timpul căruia un spermatozoid intră într-un ovul și nucleii lor fuzionează. Ca urmare a fuziunii gameților, se formează un zigot - în esență unul nou, capabil să se dezvolte dacă sunt prezente condițiile necesare pentru aceasta. Fertilizarea determină activarea oului, stimulându-l la modificări succesive, ducând la dezvoltarea unui organism format.
Când un spermatozoid intră în contact cu suprafața oului, membrana gălbenușă a oului se schimbă, transformându-se într-o membrană de fertilizare. Această modificare este considerată o dovadă că a avut loc activarea ouălor. În același timp, pe suprafața ouălor care conțin puțin sau deloc gălbenuș, un așa-zis. o reacție corticală care împiedică alți spermatozoizi să intre în ovul. La ouăle care conțin mult gălbenuș, reacția corticală are loc mai târziu, astfel încât mai mulți spermatozoizi pătrund de obicei în ele. Dar chiar și în astfel de cazuri, un singur spermatozoid, primul care ajunge la nucleul ovulului, este fertilizat.
În unele ouă, la locul de contact al spermatozoizilor cu membrana plasmatică a oului, se formează o proeminență a membranei - așa-numita. tuberculul de fertilizare; facilitează pătrunderea spermatozoizilor. De obicei, capul spermatozoidului și centriolii aflați în partea mijlocie a acestuia pătrund în ovul, în timp ce coada rămâne în exterior. Centriolii contribuie la formarea fusului în timpul primei diviziuni a unui ovul fertilizat. Procesul de fertilizare poate fi considerat finalizat atunci când cei doi nuclei haploizi - ovulul și spermatozoidul - se contopesc și cromozomii lor se conjugă, pregătindu-se pentru prima zdrobire a ovulului fecundat.
Despărțirea- formarea unei blastule embrionare multicelulares.Caracteristici: a) complet, parţial; b) uniformă, neuniformă; c) sincron, asincron.
Despărțirea. Dacă aspectul membranei de fertilizare este considerat un indicator al activării oului, atunci diviziunea (zdrobirea) este primul semn al activității reale a oului fertilizat. Natura zdrobirii depinde de cantitatea și distribuția gălbenușului în ou, precum și de proprietățile ereditare ale nucleului zigot și de caracteristicile citoplasmei oului (cele din urmă sunt în întregime determinate de genotipul organismului mamei). ). Există trei tipuri de clivaj a unui ovul fertilizat.
reguli de zdrobire. S-a stabilit că fragmentarea respectă anumite reguli, numite după cercetătorii care le-au formulat primii. Regula lui Pfluger: axul trage întotdeauna în direcția cu cea mai mică rezistență. Regula lui Balfour: rata clivajului holoblastic este invers proporțională cu cantitatea de gălbenuș (gălbenușul face dificilă divizarea atât a nucleului, cât și a citoplasmei). Regula lui Sacks: celulele sunt de obicei împărțite în părți egale, iar planul fiecărei noi diviziuni intersectează planul diviziunii anterioare în unghi drept. Regula lui Hertwig: nucleul și fusul sunt de obicei situate în centrul protoplasmei active. Axa fiecărui ax de diviziune este situată de-a lungul axei lungi a masei protoplasmei. Planurile de diviziune intersectează de obicei masa protoplasmei în unghi drept cu axele sale.
Ca urmare a zdrobirii celulelor fertilizate, se formează blastomeri. Când există o mulțime de blastomeri (la amfibieni, de exemplu, de la 16 la 64 de celule), ele formează o structură care seamănă cu o zmeură și se numește morula.
Blastula. Pe măsură ce zdrobirea continuă, blastomerele devin mai mici și mai strânse între ele, dobândind o formă hexagonală. Această formă crește rigiditatea structurală a celulelor și densitatea stratului. Continuând să se dividă, celulele se împing între ele și, ca urmare, când numărul lor ajunge la câteva sute sau mii, formează o cavitate închisă - blastocelul, în care intră lichidul din celulele din jur. În general, această formațiune se numește blastula. Formarea sa (la care mișcările celulare nu participă) încheie perioada de zdrobire a ouălor.
În ouăle homolecitale, blastocelul poate fi situat central, dar în ouăle telolecitale, este de obicei deplasat de gălbenuș și este situat excentric, mai aproape de polul animal și chiar sub blastodisc. Deci, blastula este de obicei o bilă goală, a cărei cavitate (blastocoel) este umplută cu lichid, dar în ovocitele telolecitale cu fragmentare discoidală, blastula este reprezentată de o structură aplatizată.
La holoblastic clivaj, stadiul de blastula este considerat complet atunci când, ca urmare a diviziunii celulare, raportul dintre volumele citoplasmei și nucleului lor devine același ca în celulele somatice. Într-un ou fecundat, volumul gălbenușului și al citoplasmei nu corespunde deloc mărimii nucleului. Cu toate acestea, în procesul de zdrobire, cantitatea de material nuclear crește oarecum, în timp ce citoplasma și gălbenușul doar se divid. În unele ouă, raportul dintre volumul nucleului și volumul citoplasmei în momentul fertilizării este de aproximativ 1:400, iar până la sfârșitul etapei de blastula este de aproximativ 1:7. Acesta din urmă este aproape de raportul caracteristic atât celulelor reproductive primare, cât și celulelor somatice.
gastrulatie
1. Formarea unui nucleu multistrat.
2. Următoarea etapă după zdrobire e mbr si geneza a .
3. Tipul de gastrulatieaideterminată de tipul de ou şi de tipul de zdrobire a zigoţilors.
4. Gastrulatia precoce si am intarziat.
În timpul gastrulației ai procesele au loc:
Ovoplasmatic da segregare
prezumtiv s complot și
Proliferare
Diferenţiere
Inducţie
Comitet hohotitor
Expresia genelor
Reprimarea genelor
Rolul biologic – educație e cotoderm sși endodermul s
|
Tipul de gastrulatie ai |
reprezentanți |
Tip ouă |
Despărțirea |
Tip gastruli si |
|
Invaginatie |
Lancelet |
Oligolecitalși solecital I |
Sincron complet uniform |
celoblastula |
|
e pibolia |
Amfibieni |
Moderat polilecital |
Asincron complet neuniform |
Amfiblastula |
|
delaminare |
Insecte |
Polilecital |
superficial |
Periblastula |
|
Migrația |
Păsări |
Polilecital |
Meroblastic |
|
Gastrulare tardivăși eu |
Din timp |
Sursa dezvoltării mezodermuluis |
Mecanism |
|
Electroceln s th |
Invaginatie |
Endoderm |
flambaj |
|
Teloblastic esk uy |
e pibolia |
Teloblast sbuzele laterale ale blastoporului |
in miscare |
|
Migrație cu formare primitivă de dungi |
Migrația și diviziuneași nat și cu mine |
E cotoderma |
in miscare |
Organele provizorii
1. Amnion
2. Sacul vitelin
3. Al antois
4. Corion
5. Placenta
6. Membrană seroasă
tipuri de alimente
1. Vitelotrofe f - 30 ore, includerea gălbenușului a ovocitului.
2. Histiotrofic - a 2-a zi - a 3-a luna, țesuturile din jur.
3. Hematotrof e - luna a 3-a - până la naștere, placentă.
Gastrula. Gastrula este etapa de dezvoltare embrionară în care embrionul este format din două straturi: exteriorul - ectoderm și interiorul - endodermul. La diferite animale se ajunge la acest stadiu dublu strat căi diferite deoarece ouăle de diferite specii conţin sumă diferită gălbenuş. Cu toate acestea, în orice caz, rolul principal în acest lucru este jucat de mișcările celulare, și nu de diviziunile celulare.
Invaginatie. În ouăle homolecitale, care sunt de obicei holoblastic zdrobirea, gastrulația are loc de obicei prin invaginare ( invaginare) a celulelor polului vegetativ, ceea ce duce la formarea unui embrion cu două straturi, având forma unui bol. Blastocelul original se contractă, dar se formează o nouă cavitate, gastrocelul. Deschiderea care duce în acest nou gastrocel se numește blastopor (un nume nefericit deoarece se deschide nu în blastocel, ci în gastrocel). Blastoporul este situat în regiunea viitorului anus, la capătul posterior al embrionului, iar în această regiune se dezvoltă cea mai mare parte a mezodermului - al treilea, sau mijlociu, strat germinal. Gastrocoelul este numit și archenteron sau intestinul primar și servește ca rudiment al sistemului digestiv.
Involuţie. La reptile și păsări, ale căror ouă telolecitale conțin o cantitate mare de gălbenuș și sunt zdrobite meroblastic, celulele blastulelor dintr-o zonă foarte mică se ridică deasupra gălbenușului și apoi încep să se înșurubească spre interior, sub celulele stratului superior, formând al doilea strat (inferior). Acest proces de înșurubare a foii celulare se numește involuție. Stratul superior al celulelor devine stratul germinal exterior sau ectodermul, iar stratul inferior devine interiorul sau endodermul. Aceste straturi se contopesc unul în celălalt, iar locul în care are loc tranziția este cunoscut sub numele de buza blastopor. Acoperișul intestinului primar în embrionii acestor animale este format din celule endodermice complet formate și fundul gălbenușului; fundul celulelor se formează mai târziu.
delaminare . La mamiferele superioare, inclusiv la om, gastrulația are loc oarecum diferit, și anume prin delaminare, dar duce la același rezultat - formarea unui embrion cu două straturi. Delaminarea este o stratificare a stratului exterior original de celule, care duce la apariția unui strat interior de celule, adică. endoderm.
rezultatele gastrulației. Rezultatul final al gastrulației este formarea unui embrion cu două straturi. Stratul exterior al embrionului (ectodermul) este format din celule mici, adesea pigmentate, care nu conțin gălbenuș; din ectoderm se dezvoltă în continuare țesuturi precum cele nervoase și straturile superioare ale pielii. Stratul interior (endodermul) este format din celule aproape nepigmentate care rețin ceva gălbenuș; ele dau naştere în principal ţesuturilor care căptuşesc tubul digestiv şi derivaţilor acestuia.
GASTRULAREA FETALULUI UM
Gastrulare precoce Și eu - 7a-14 s zi.
Delaminarea embr și a zonei pe ep și blast și g și poblast (primar uh cotoderma si primara endoderm).
E piblast - amn si otich esk o bule.
Hipoblast -g brazi fac bule.
Trofoblast - citotrofoblast și sincitși otrofoblast.
Discul germinal = fundus amn si otich esk wow + wow brad bule.
De fapt material germinativ - partea de jos a amn si otich esk wow bubble.
Gastrulare tardivă Și eu 14a-17 s zi ki .
Migrație cu formarea striei primare.
Germen exterior de mai sus wah eu mezodermul migrează din discul germinal A .
Toate cele 3 straturi ale embrionului sunt formate din e cotoderm s.
Caracteristicile gastrulațieiaifat uman:
Subecuația completă zdrobirea asincronă a zigoțilors.
Dezvoltare avansată germenul exterior de mai sus V da organe.
Implantarea embrionului în endometru și placentăși eu.
Toate cele trei straturi germinale sunt formate din e cotoderm s.
Frunze germinale. Ectodermul, endodermul și mezodermul se disting pe baza a două criterii. În primul rând, prin localizarea lor în embrion în fazele incipiente ale dezvoltării sale: în această perioadă, ectodermul este întotdeauna situat în exterior, endodermul este în interior, iar mezodermul, care apare ultimul, se află între ele. În al doilea rând, în funcție de rolul lor viitor: fiecare dintre aceste foi dă naștere anumitor organe și țesuturi și sunt adesea identificate prin soarta lor ulterioară în procesul de dezvoltare. Totuși, reamintim că în perioada în care au apărut aceste pliante, nu au existat diferențe fundamentale între ele. În experimentele privind transplantul de straturi germinale, s-a demonstrat că inițial fiecare dintre ele are potența oricăreia dintre celelalte două. Astfel, distincția lor este artificială, dar este foarte convenabil să o folosești în studiul dezvoltării embrionare.
Mezodermul, adică stratul germinal mijlociu se formează în mai multe moduri. Poate apărea direct din endoderm prin formarea de saci celomici, ca în lancetă; simultan cu endodermul, ca la broasca; sau prin delaminare, din ectoderm, ca la unele mamifere. În orice caz, la început mezodermul este un strat de celule care se află în spațiul care a fost ocupat inițial de blastocel, adică. intre ectoderm la exterior si endoderm la interior.
Mezodermul se împarte curând în două straturi celulare, între care se formează o cavitate numită celom. Din această cavitate s-au format ulterior cavitatea pericardică care înconjoară inima, cavitatea pleurală care înconjoară plămânii și cavitatea abdominală, în care se află organele digestive. Stratul exterior al mezodermului - mezodermul somatic - formează, împreună cu ectodermul, așa-numitul. somatopleura. Din mezodermul exterior se dezvoltă mușchii striați ai trunchiului și membrelor, țesutul conjunctiv și elementele vasculare ale pielii. Stratul interior de celule mezodermice se numeste mezoderm splanhnic si impreuna cu endodermul formeaza splanhnopleura. Din acest strat de mezoderm se dezvoltă mușchii netezi și elementele vasculare ale tractului digestiv și derivații acestuia. În embrionul în curs de dezvoltare, există o mulțime de mezenchim liber (mezoderm embrionar) care umple spațiul dintre ectoderm și endoderm.
Derivați ai straturilor germinale. Soarta ulterioară a celor trei straturi germinale este diferită. Din ectoderm se dezvoltă: tot țesutul nervos; straturile exterioare ale pielii și derivații acesteia (păr, unghii, smalț al dinților) și parțial mucoase cavitatea bucală, cavități nazale și anus.
Endodermul dă naștere căptușelii întregului tub digestiv - de la cavitatea bucală până la anus - și tuturor derivaților săi, adică. timus, tiroida, glande paratiroide, trahee, plamani, ficat si pancreas.
Din mezoderm se formează: toate tipurile de țesut conjunctiv, țesut osos și cartilaj, sânge și sistemul vascular; toate tipurile de țesut muscular; sistemele excretor și reproducător, stratul dermic al pielii.
Un animal adult are foarte puține astfel de organe. endodermic origine, care nu ar conține celule nervoase provenite din ectoderm. Fiecare organ important conține, de asemenea, derivați ai mezodermului - vase de sânge, sânge și adesea mușchi, astfel încât izolarea structurală a straturilor germinale să fie păstrată numai în stadiul formării lor. Deja la începutul dezvoltării lor, toate organele capătă o structură complexă și includ derivați ai tuturor straturilor germinale.
Membrane extraembrionare. La animalele care depun ouă pe uscat sau vivipare, embrionul are nevoie de membrane suplimentare care să-l protejeze de deshidratare (dacă ouăle sunt depuse pe uscat) și să ofere nutriție, îndepărtarea produselor finite ai metabolismului și schimbul de gaze.
Aceste funcții sunt îndeplinite de membranele extraembrionare - amnios, corion, sacul vitelin și alantois, care se formează în timpul dezvoltării la toate reptilele, păsările și mamiferele. Corionul și amnionul sunt strâns legate ca origine; se dezvoltă din mezodermul şi ectodermul somatic. Chorion - coaja cea mai exterioară care înconjoară embrionul și alte trei scoici; acest înveliș este permeabil la gaze și prin ea are loc schimbul de gaze.
Amnionul protejează celulele fătului de uscare datorită lichidului amniotic secretat de celulele sale. Sacul vitelin umplut cu gălbenuș, împreună cu tulpina de gălbenuș, furnizează embrionului nutrienții digerați; acest înveliș conține o rețea densă de vase de sânge și celule care produc enzime digestive. Sacul vitelin, ca și alantoida, se formează din mezodermul și endodermul splanhnic: endodermul și mezodermul se răspândesc pe toată suprafața gălbenușului, crescându-l în exces, astfel încât în final întregul gălbenuș se află în sacul vitelin. La mamifere, aceste funcții importante sunt îndeplinite de placentă, un organ complex format din vilozități coriale, care, crescând, pătrund în adânciturile (criptele) mucoasei uterine, unde vin în contact strâns cu vasele de sânge și glandele acesteia.
La om, placenta asigură pe deplin respirația embrionului, nutriția și eliberarea de produse metabolice în fluxul sanguin al mamei.
PĂRȚI ALE COCHIIEI
A. decidua basalis - partea maternă a placentei
B. Decidua capsularis - acoperă embrionul (fătul) - deșeuri de pungă
C. decidua parietalis - parietal
Placenta este discoida, grosime 3 cm, diametru 15-25 cm, greutate 500-600 g.
HEMOCORIALS Y BARIERĂ
1. Endoteliul capilar.
2. Membrană bazală.
3. Țesutul conjunctiv al vilozităților cu celule Kashchenko Hofbau e ra.
4. Membrana bazală a citotrofoblastului.
5. Citotrofoblast
6. Sincitiotrofoblast
7. De la 4 luni. f i brino i d Langhans îl înlocuiește pe 5.
Placenta umană: tip II A, discoidal, hemocor Șial.
placenta MFI - cotiledon (15-20)
A. Plodova parte a placentei - cor vilos si el.
B. Partea mamă - bazalăotpadn iar eu sunt endometrul.
Membranele extraembrionare nu se păstrează în perioada postembrionară. La reptile și păsări, atunci când eclozează, cojile uscate rămân în coaja de ou. La mamifere, placenta și alte membrane extraembrionare sunt eliminate din uter (respinse) după nașterea fătului. Aceste cochilii au oferit vertebratelor superioare independență față de mediu acvaticși, fără îndoială, a jucat un rol important în evoluția vertebratelor, în special în apariția mamiferelor.
Perioada critica - o scurtă perioadă de sensibilitate crescută a embrionului, când apar modificări calitative importante în acesta.
Progeneza
Fertilizare
Implantare - 7-8 zile
Placentația – a 3-a și a-a-8-a săptămână
Dezvoltarea creierului - 15și eu-24 și eu săptămâni și
Dezvoltarea inimii
Naștere
perioada neonatală
Anii adolescenței
Ciclurile menstruale la femei
Menopauza
fluctuații sezoniere
fertilizare in vitro
1976 Luisa Brown (GB) Edvards și Stantow
1. Chirurgie
2. Fertilizarea „in vitro”
3. Incubare 3-4 zile (zdrobire)
4. Blastocist (18-32 blastomere) - „blastocist liber” este plasat în uter
5. Implantarea începe în a 6-a-7 zi (15% succes)
Eextracorporale Oe fertilizareaepermite
1. Alegeți sexul copilului
2. Îmbogățiți (îmbunătățiți) spermatozoizii
3. Asistați spermatozoizii în mișcarea și dizolvarea membranelor ovocitelor
4. Tratați unele tipuri de infertilitate feminină
5. Excludeți sarcina ectopică
Surse de informare:
A)principal
1. Materiale pentru pregătirea unei lecții practice pe tema„Fundamentele embriologiei vertebratelor. Dezvoltarea embrionară a omului. celulele sexuale. Fertilizare, zdrobire.” din tdmu . edu. ua.
2. Prezentarea prelegerii „Embriologie generală și comparată” din tdmu . edu. ua.
4. Histologie, citologie și embriologie / [Afanasiev Yu. I., Yurinși N.A. , Kotovsky E. F. și alții.] ; ed. Yu.I. Afanasiev, N.A. Yurina. – [Ed. a 5-a, revizuită. si suplimentare] . –M. : Medicament. - 2002. - De când. 93 –107 .
5. Histologie: [manual] / ed. DE EXEMPLU. Ulumbekov a, Yu.A. Chelsheva. –[ Ed. a II-a, revizuită. si suplimentare] . – M. : GEOTAR-M ED, 2001. - S. 104-107.
6. Danilov R.K. Histologie. Embriologie. Citologie. : [manual pentru studenții la medicină]/ R. K. Danilov - M .: SRL „Agenția de informații medicale”, 2006. - S. 73–83.
b) suplimentare
1. Atelier de histologie, citologie și embriologie. Editat de N.A. Yurina, A.I. Radostina. G., 1989.- S.40-46.
2. Histologia oamenilor / [Lutsik O. D., Ivanova A. I., Kabak K. S., Chaikovsky Yu. B.]. - Kiev: Book plus, 2003. - S. 72-109.
3. Volkov K.S. Ultrastructura principalelor componente ale sistemelor de organe ale corpului:n avchalny ajutor-atlas/ K.S. Volkov, N.V. Pasechk O . – Ternopil : Ukrmedkniga, 1997. - S.95-99.