chemins de projection. Voies afférentes des nerfs crâniens Schémas de la structure des voies afférentes de projection
Lire aussi
Quelques régularités dans la structure des voies de projection efférentes
1. Le premier neurone de toutes les voies efférentes est localisé dans le cortex cérébral.
2. Les voies de projection efférentes occupent la jambe antérieure, le genou et la partie antérieure de la jambe postérieure de la capsule interne, passent à la base des jambes du cerveau et du pont.
3. Toutes les voies efférentes se terminent dans les noyaux des nerfs crâniens moteurs et dans les cornes antérieures de la moelle épinière, où se trouve le dernier motoneurone.
4. Les voies efférentes forment un croisement complet ou partiel, à la suite duquel les impulsions du cortex cérébral sont transmises aux muscles de la moitié opposée du corps.
Les organes des sens réalisent la perception de divers stimuli agissant sur l'organisme humain et animal, ainsi que l'analyse primaire de ces stimuli. L'académicien IP Pavlov a défini les organes sensoriels comme des zones périphériques des analyseurs. Leurs éléments de perception spécifiques sont les terminaisons nerveuses sensibles - des récepteurs qui convertissent l'énergie d'un stimulus externe en impulsions nerveuses. Ces derniers contiennent sous forme codée des informations sur les objets et les phénomènes. monde extérieur. Ces impulsions sont transmises le long des voies nerveuses afférentes au sous-cortical et centres corticaux, où a lieu l'analyse finale des irritations. Selon la doctrine des analyseurs, les voies afférentes représentent leur section médiane conductrice, et les zones corticales percevantes sont leurs extrémités centrales. L'apparition des sensations est associée aux sections corticales des analyseurs.
Chez les protozoaires, la sensibilité est inhérente à la couche externe du protoplasme de leur cellule unique. Chez les animaux inférieurs, dont le corps est constitué d'endoderme et d'ectoderme, toutes les cellules de ce dernier répondent aux stimuli externes. Parallèlement à la différenciation musculaire et système nerveux dans l'ectoderme sont isolées des cellules perceptives séparées, associées au système nerveux central et représentant les cellules sensorielles primaires: d'abord (dans les cavités intestinales inférieures), elles sont dispersées dans tout le corps, puis elles sont regroupées à certains endroits, notamment autour du bouche. Ces groupes de cellules sensorielles sont les plus simples dans la structure et les fonctions des organes sensoriels. Enfin, des formes plus parfaites sont observées dans les supérieurs, où les organes sensoriels comprennent non seulement des éléments perceptifs, mais également des appareils spéciaux supplémentaires (auxiliaires): d'abord des cellules épithéliales indifférentes (de soutien), puis des tissus conjonctifs et musculaires.
Au cours de l'évolution, des organes sont développés qui sont adaptés pour percevoir une grande variété d'agents. environnement- mécanique, physique, chimique. Par exemple, les termites perçoivent un champ magnétique, les abeilles et les fourmis - rayons ultra-violets, cafards et calamars - rayons infrarouges, les poissons ont un organe de la ligne latérale qui détecte la direction et la vitesse du mouvement de l'eau, tandis que les musaraignes et les chauves-souris capable de recevoir des vibrations ultrasonores. Chez les animaux supérieurs et les humains, les organes sensoriels sont l'organe de l'odorat, l'organe du goût, l'organe de la vision, l'organe vestibulocochléaire et la peau qui, avec ses appendices, forme l'enveloppe générale du corps.
Sur la base des caractéristiques de développement, de structure et de fonction, on distingue 3 types d'organes sensoriels. Le type I comprend les organes de la vision et de l'odorat, qui sont établis dans l'embryon en tant que partie du cerveau. Leur structure est basée sur des cellules sensorielles primaires, ou neurosensorielles. Ces cellules ont des processus périphériques spécialisés qui perçoivent les vibrations des ondes lumineuses ou des molécules de substances volatiles, et des processus centraux par lesquels l'excitation est transmise aux neurones afférents.
Au type II appartiennent les organes du goût, de l'ouïe et de l'équilibre. Ils sont pondus dans la période embryonnaire sous la forme d'un épaississement de l'ectoderme, les placodes. Leur principal élément récepteur est constitué par les cellules épithéliales sensorielles à détection secondaire. Contrairement aux cellules neurosensorielles, elles n'ont pas de processus de type axone. L'excitation qui en résulte sous l'influence de substances aromatisantes, de vibrations de l'air ou d'un milieu liquide est transmise aux terminaisons des nerfs correspondants.
Le troisième type d'organes sensoriels est représenté par des corps et des formations récepteurs encapsulés ou non encapsulés. Ceux-ci incluent des récepteurs dans la peau et le tissu sous-cutané. Ce sont des terminaisons nerveuses entourées de tissu conjonctif ou de cellules gliales. caractéristique commune de toutes les cellules réceptives est la présence de flagelles - kinocils ou microvillosités - stéréocils. Des molécules de protéines photo-, chimio- et mécanoréceptrices spéciales sont intégrées dans la membrane plasmique des flagelles et des microvillosités. Ces molécules perçoivent des impacts d'un seul type spécifique et les codent en informations spécifiques de la cellule, qui sont transmises aux centres nerveux correspondants.
Les organes sensoriels diffèrent également par la complexité de leur structure anatomique. Les organes du goût et de la sensation cutanée sont relativement simples, représentés principalement par des formations épithéliales. Les organes de l'odorat, de la vision, de l'ouïe et de l'équilibre disposent d'appareils auxiliaires qui garantissent que seuls les stimuli leur sont fournis, à la perception desquels ces organes sensoriels sont adaptés. Ainsi, l'appareil auxiliaire de l'organe olfactif est le labyrinthe ethmoïde et les sinus paranasaux, qui dirigent le flux d'air vers les récepteurs olfactifs. L'organe de la vision est équipé d'un appareil optique qui projette une image d'objets extérieurs sur la rétine de l'œil. L'organe de l'audition a un complexe appareil agencé capter et transmettre des sons.
Les appareils auxiliaires des organes sensoriels assurent non seulement l'interaction de stimuli spécifiques avec les récepteurs, mais bloquent également le chemin des stimuli étrangers et inadéquats et protègent également les organes sensoriels des influences mécaniques externes et des dommages.
Caractéristiques générales des voies motrices descendantes :
1. Schéma à 2 neurones de la structure ;
2. les fibres d'un neurone se croisent ;
3. 2 neurone - dans les cornes antérieures de la moelle épinière.
Les voies de projection descendantes (effectrices, efférentes) conduisent les impulsions du cortex, des centres sous-corticaux aux sections sous-jacentes, aux noyaux du tronc cérébral et aux noyaux moteurs des cornes antérieures de la moelle épinière. Ces chemins peuvent être divisés en deux groupes : 1) le moteur principal, ou pyramidale, voie (voies cortico-nucléaires et cortico-spinales)
transporte les impulsions des mouvements volontaires du cortex cérébral aux muscles squelettiques de la tête, du cou, du tronc et des membres via les noyaux moteurs correspondants du cerveau et de la moelle épinière;
2) voies motrices extrapyramidales
transmettent les impulsions des centres sous-corticaux aux noyaux moteurs des nerfs crâniens et spinaux, puis aux muscles.
La voie pyramidale comprend un système de fibres le long desquelles les impulsions motrices du cortex cérébral, du gyrus précentral, des neurones pyramidaux géants (cellules de Betz) sont dirigées vers les noyaux moteurs des nerfs crâniens et les cornes antérieures de la moelle épinière, et d'eux aux muscles squelettiques. Compte tenu de la direction des fibres, ainsi que de l'emplacement des faisceaux dans le tronc cérébral et les cordons de la moelle épinière, le trajet pyramidal est divisé en trois parties : 1) cortico-nucléaire - aux noyaux des nerfs crâniens ; 2) cortico-spinal latéral (pyramidal) ) - aux noyaux des cornes antérieures de la moelle épinière; 3) corticospinal antérieur (pyramidal) - également aux cornes antérieures de la moelle épinière.
Voie cortico-nucléaire est un faisceau de processus de neurones pyramidaux géants, qui du cortex tiers inférieur du gyrus précentral
descendre jusqu'à la capsule interne et traverser son genou. De plus, les fibres de la voie cortico-nucléaire vont à la base du tronc cérébral. Partant du mésencéphale et plus loin, dans le pont et le bulbe rachidien, les fibres de la voie cortico-nucléaire passer du côté opposé aux noyaux moteurs des nerfs crâniens
III et IV - dans le mésencéphale, V, VI, VII - dans le pont, IX, X, XI, XII - dans
moelle allongée. Dans ces noyaux, le chemin cortico-nucléaire (pyramidal) se termine. Ses fibres constitutives forment des synapses avec les cellules motrices de ces noyaux. Les processus des cellules motrices mentionnées quittent le cerveau dans le cadre des nerfs crâniens correspondants et sont envoyés aux muscles squelettiques de la tête et du cou et les innervent.
Voies corticospinales latérales et antérieures (pyramidales) , proviennent également de neurones pyramidaux géants gyrus précentral, ses 2/3 supérieurs . Les axones de ces cellules vont à la capsule interne, traversent la partie antérieure de son pédicule postérieur (juste derrière les fibres du tractus cortico-nucléaire) et descendent jusqu'à la base du pédoncule cérébral. De plus, les fibres cortico-spinales, descendantes, pénètrent dans les faisceaux de fibres du pont allant dans le sens transversal et sortent dans moelle , où sur sa surface avant (inférieure) ils forment des rouleaux saillants - pyramides . Au bas de la moelle allongée une partie des fibres va du côté opposé et continue dans la moelle épinière latérale, se terminant progressivement dans les cornes antérieures de la moelle épinière avec des synapses sur les cellules motrices de ses noyaux.
Cette la partie des voies pyramidales impliquée dans la formation de la décussation pyramidale (décussation motrice) est appelée voie latérale cortico-spinale (pyramidale). Les fibres du tractus cortico-spinal qui ne participent pas à la formation de la décussation pyramidale et ne se croisent pas du côté opposé continuent leur chemin vers le bas dans le cadre du funicule antérieur de la moelle épinière. Ces fibres constituent la voie corticospinale antérieure (pyramidale).
Ensuite, ces fibres passent également du côté opposé, mais à travers la commissure blanche de la moelle épinière et fin sur les cellules motrices de la corne antérieure
côté opposé de la moelle épinière. Il convient de noter que tous les chemins pyramidaux sont croisés, c'est-à-dire leurs fibres sur leur chemin vers le prochain neurone plus tôt ou
tard de l'autre côté.
Le deuxième neurone de la voie motrice volontaire descendante (cortico-spinale) est constitué des cellules des cornes antérieures de la moelle épinière, dont les longs processus quittent la moelle épinière dans le cadre des racines antérieures et sont envoyés dans le cadre des nerfs spinaux pour innerver les muscles squelettiques.
Voies extrapyramidales, réunis en un seul groupe, contrairement aux voies pyramidales, ont des connexions étendues dans le tronc cérébral et avec le cortex cérébral, qui a repris les fonctions de contrôle et de gestion du système extrapyramidal. Le cortex cérébral, qui reçoit des impulsions à la fois le long des voies sensorielles ascendantes directes (direction corticale) et des centres sous-corticaux, contrôle les fonctions motrices du corps par des voies extrapyramidales et pyramidales.
Le cortex cérébral influence les fonctions motrices de la moelle épinière à travers le système de noyaux cervelet-rouge, à travers la formation réticulaire, qui a des connexions avec le thalamus et le striatum, à travers les noyaux vestibulaires.
Ainsi, les centres du système extrapyramidal comprennent noyaux rouges, dont l'une des fonctions est maintenir tonus musculaire
nécessaire pour maintenir le corps dans un état d'équilibre sans effort de volonté. Le noyau rouge, qui appartient également à la formation réticulaire, reçoit des impulsions du cortex cérébral, du cervelet(issues des voies proprioceptives cérébelleuses) et elle-même reliée aux noyaux moteurs des cornes antérieures de la moelle épinière.
Tractus nucléo-spinal rouge
fait partie de l'arc réflexe dont le chaînon porteur est les voies proprioceptives spino-cérébelleuses. Par ici provient du noyau rouge
(paquet de Monakov), passe du côté opposé (traverser
Truite) et descend dans le funicule latéral
moelle épinière, se terminant sur les cellules motrices de la moelle épinière. Les fibres de ce chemin passent dans le dos (pneu) du pont et les sections latérales de la moelle allongée.
Un maillon important dans la coordination des fonctions motrices du corps humain est tractus vestibulocérébral . Il relie les noyaux de l'appareil vestibulaire aux cornes antérieures de la moelle épinière et assure les réactions d'ajustement du corps en cas de déséquilibre. Les axones des cellules participent à la formation du tractus vestibulospinal noyaux vestibulaires nerf vestibulocochléaire. Ces fibres descendent dans cordon antérieur moelle épinière et se terminent à cellules motrices des cornes antérieures moelle épinière.
Les noyaux qui forment le tractus vestibulo-spinal sont situés dans lien direct avec le cervelet, ainsi qu'avec faisceau longitudinal postérieur, qui à son tour est relié aux noyaux des nerfs oculomoteurs. La présence de communication avec les noyaux des nerfs oculomoteurs assure la préservation de la position globe oculaire(direction de l'axe visuel) en tournant la tête et le cou.
Dans la formation du faisceau longitudinal postérieur et des fibres qui atteignent les cornes antérieures de la moelle épinière (tractus réticulo-spinal) les cellules sont impliquées formation réticulaire partie souche du cerveau, principalement le noyau intermédiaire (noyau de Kahal), le noyau de la commissure épithalamique (postérieure) (noyau de Darkshevich), auquel viennent fibres des noyaux basaux des hémisphères grand cerveau.
Contrôle des fonctions du cervelet, qui participe à la coordination des mouvements de la tête, du tronc et des membres et est associé à son tour aux noyaux rouges et Appareil vestibulaire, est effectuée à partir du cortex cérébral à travers le pont le long tractus cortico-pontocérébelleux . Ce chemin de guidage est composé de deux neurones . corps cellulaires premier neurone se trouvent dans le cortex des lobes frontal, temporal, pariétal et occipital. Leurs processus - les fibres du pont cortical sont envoyées à la capsule interne et la traversent. Les fibres du lobe frontal, que l'on peut appeler fibres de pont frontales, traversent la crus antérieure de la capsule interne, les fibres nerveuses des lobes temporal, pariétal et occipital par la crus postérieure. De plus, les fibres des voies du pont cortical traversent la base du tronc cérébral. Dans la partie antérieure (à la base) du pont, les fibres des voies du pont cortical se terminent en synapses sur les cellules des noyaux du pont du même côté du cerveau. Les cellules des noyaux du pont avec leurs processus constituent deuxième neurone tractus cortico-pontocérébelleux. Les axones des cellules des noyaux du pont sont repliés en faisceaux - les fibres transversales du pont, qui passent du côté opposé, traversent en même temps dans le sens transversal les faisceaux descendants de fibres des voies pyramidales et à travers le pédicule cérébelleux moyen est envoyé à l'hémisphère cérébelleux du côté opposé.
Ainsi, les voies du cerveau et de la moelle épinière établissent des connexions entre les centres afférents et efférents (effecteurs), participent à la formation d'arcs réflexes complexes dans le corps humain. Certaines voies conductrices (systèmes de fibres) commencent ou se terminent dans le tronc cérébral et les noyaux, qui assurent des fonctions qui ont une certaine automaticité. Ces fonctions (par exemple, le tonus musculaire, les mouvements réflexes automatiques) sont réalisées sans la participation de la conscience, bien que sous le contrôle du cortex cérébral. D'autres voies transmettent des impulsions vers le cortex cérébral, vers les parties supérieures du système nerveux central, ou du cortex vers les centres sous-corticaux (vers les ganglions de la base, les noyaux du tronc cérébral et la moelle épinière). Les voies conductrices unissent fonctionnellement le corps en un seul tout, assurent la cohérence de ses actions.
question testà la conférence:
1. Caractéristiques générales des voies motrices.
2. Éléments structurels et fonctionnels de la voie cortico-nucléaire.
3. Éléments structurels et fonctionnels de la voie cortico-nucléaire.
4. Éléments structurels et fonctionnels du système extrapyramidal.
5. Le rôle des structures sous-corticales dans la formation du tonus musculaire.
6. Le rôle des éléments structuraux du mésencéphale dans la régulation du tonus musculaire et la régulation du réflexe quadrigéminal.
Chemins de projection sont constitués de neurones et de leurs fibres qui assurent les connexions entre la moelle épinière et le cerveau. Les voies de projection relient également les noyaux du tronc aux noyaux basaux et au cortex cérébral, ainsi que les noyaux du tronc au cortex et aux noyaux du cervelet. Les chemins de projection peuvent être Ascendant et descendant.
Ascendant(sensoriel, sensible ou afférent)chemins de projection conduire les impulsions nerveuses des extero-, proprio- et interorécepteurs (terminaisons nerveuses sensibles de la peau, organes du système musculo-squelettique)
tableau 7, MAIS.
| Voies de projection ascendantes | ||
| Type de voies | Valeur fonctionnelle | Localisation |
| au cortex cérébral | ||
| Toutes les voies sensorielles sont communes et types spéciaux sensibilité | Conduction des informations sensorielles de toutes les modalités vers le cortex cérébral | Substance blanche de la moelle épinière tegmentum de la capsule interne du tronc cérébral fibres thalamo-corticales (radiosité) |
| au cervelet | ||
| Voies sensorielles proprioceptives et voies sensorielles d'autres modalités | Transmission des informations sensorielles au cortex cérébelleux | Substance blanche de la moelle épinière Pédoncules cérébelleux inférieurs Pédoncules cérébelleux supérieurs |
| au toit du mésencéphale (le quadrigemina) | ||
| Tractus spino-tectal | Conduction des informations sensorielles vers les noyaux du quadrigemina | Substance blanche de la moelle épinière du tegmentum du tronc cérébral |
appareil, organes internes), ainsi que des organes des sens dans une direction ascendante vers le cerveau, principalement vers le cortex cérébral, où ils se terminent principalement au niveau de la couche cytoarchitectonique IV (tableau 7, MAIS). Plus de 20 voies sensorielles ascendantes ont été décrites à ce jour ; il y a tout lieu de croire que cette liste se poursuivra. Leur caractéristique distinctive est la transmission séquentielle en plusieurs étapes d'informations sensorielles au cortex cérébral via un certain nombre de centres nerveux intermédiaires.
En plus du cortex cérébral, des informations sensorielles sont envoyées à d'autres parties du système nerveux, à savoir le cervelet, le mésencéphale et la formation réticulaire.
descendant(efférent ou centrifuge)chemins de projection conduire les impulsions nerveuses du cortex cérébral, où elles proviennent des neurones pyramidaux de la cinquième couche cytoarchitectonique, aux noyaux basaux et souches du cerveau et ensuite aux noyaux moteurs de la moelle épinière et du tronc cérébral (tableau 7, B). Ils transmettent des informations liées à la programmation des mouvements du corps dans des situations spécifiques, ce sont donc des voies motrices. caractéristique commune voies motrices descendantes est qu'elles passent nécessairement par capsule interne- une couche de substance blanche dans les hémisphères cérébraux qui sépare le thalamus des ganglions de la base. Dans le tronc cérébral, la plupart des descendants
Tableau 7, B. Caractéristiques générales des chemins conducteurs
| Voies de projection descendantes | ||
| Type de voies | Valeur fonctionnelle | Localisation |
| Connexions de projection du cortex cérébral | ||
| Voies cortico-spinales et cortico-nucléaires (système pyramidal) | Exercer des influences corticales sur les centres moteurs de la moelle épinière et du tronc cérébral | Capsule interne de matière blanche de la base du cordon du tronc cérébral de la moelle épinière |
| Voies rouges nucléo-spinales, réticulo-spinales, vestibulospinales et tectospinales (système extrapyramidal) | Conduire l'influence des structures du système extrapyramidal sur les centres moteurs de la moelle épinière et du tronc cérébral | substance blanche base du tronc cérébral moelle épinière |
| Connexions de projection du cerveau avec le cervelet | ||
| Voies cortico-pontine et pontine-cérébelleuse | Effectuer des influences corticales sur les centres du cervelet | substance blanche capsule interne base du tronc cérébral pédoncules cérébelleux moyens |
les voies menant à la moelle épinière et au cervelet vont à sa base.
Parmi les voies descendantes, les voies du système extrapyramidal, ainsi que les voies réticulo-spinales, ont une importance significative.
En tableau. sept; sept, MAIS et 7, B donné caractéristiques générales les voies de projection les plus importantes ; ils sont discutés plus en détail ci-dessous.
Il convient de noter que la subdivision des voies descendantes impliquées dans la régulation et la coordination des mouvements dans les systèmes pyramidaux et extrapyramidaux est largement conditionnelle et ne reflète pas pleinement la complexité des relations hiérarchiques entre le cortex et de nombreux centres moteurs sous-corticaux, qui, de plus, ont une préhistoire différente du développement onto- et phylogénétique.
ROSZDRAV
Etablissement public d'enseignement supérieur professionnel
UNIVERSITÉ MÉDICALE D'ÉTAT DE L'EXTRÊME-ORIENT
AGENCE FÉDÉRALE DE LA SANTÉ ET DU DÉVELOPPEMENT SOCIAL
DÉPARTEMENT D'ANATOMIE HUMAINE
VOIES DU CERVEAU ET DE LA MOELLE ÉPINIÈRE
Manuel pédagogique et méthodique pour les étudiants en médecine et pédiatrie.
Les facultés
Khabarovsk
UDC 611.81 + 611.82 (075.8) LBC 28.706ya 73 P 782
COMPILATEURS :
Professeur agrégé G.A. Ivanenko, professeur agrégé A.V. Kouznetsov
REVISEURS :
Docteur en Sciences Médicales, Professeur B.Ya. Ryzhavsky Docteur en sciences médicales, professeur A.M. Hélimski
Approuvé lors de la réunion de l'Université CMS 23.01.07.
Avant-propos ……………………………………………………………………4
Classification des parcours …………………………………………5
Voies de projection …………………………………………...6
Voies afférentes ………………………………………...6
Voies proprioceptives (profondes)
sensibilité …………………………………………………………..7
Chemin conducteur de l'analyseur de peau ……………………………….11
Voie conductrice de la sensibilité tactile …………………….14
Voies proprioceptives vers le cervelet …………………………………………...17
Chemin conducteur de l'analyseur intéroceptif ………………….24
Voies efférentes ……………………………………….26
Voies pyramidales ……………………………………………………………………………………………………………………………………… ……………………………………………………………………27
Système nerveux extrapyramidal …………………………………….36
Tractus réticulo-spinal ………………………………………...41
Voie prédverno-rachidienne …………………………………………….41
Voies descendantes du cervelet ……………………………………………………..42
Voies de commissariat ……………………………………….46
Parcours associatif ……………………………………………48
AVANT-PROPOS
La section "Anatomie du système nerveux central" est l'une des plus difficiles du cours d'anatomie humaine étudiée par les étudiants -
médecins. Son importance est déterminée par le rôle qu'il joue dans la formation de la vision du monde matérialiste dialectique des élèves.
vision, compréhension correcte des comportements simples et complexes,
tout le système de conscience et de raison dans le comportement humain, sa pensée, sa mémoire et son travail créatif.
Leçon sur le thème "Voies conductrices du système nerveux central"
est la finale, résumant le travail des étudiants dans l'étude
Institut de recherche d'anatomie du SNC. Dans les diverses activités du système nerveux,
Les voies sont le substrat morphologique qui fournit des connexions entre les différentes structures cérébrales et fonctionnelles.
développement du système nerveux dans son ensemble, ils diffèrent par la complexité de la structure
et une fiabilité opérationnelle élevée.
L'étude de l'anatomie des voies du cerveau et de la moelle épinière est tâche difficile. Les manuels d'anatomie humaine ne présentent que leur description classique sans les aspects cliniques de la structure. Pour les futurs pédiatres, en outre, vous devez également connaître les caractéristiques liées à l'âge des voies, car cela a grande importance dans la pratique clinique.
Le support pédagogique proposé s'adresse aux étudiants
il y a des cours I-II des facultés de médecine et de pédiatrie en complément
matériel pour étudier le sujet "Anatomie des voies du système nerveux central". Dans le manuel, les voies conductrices sont considérées
sont prises d'un point de vue clinique, il sera donc utile pour les étudiants seniors en préparation pour les cours sur les maladies nerveuses.
Miam. Nous pensons que ce guide va médecins serviables- Inter-
nous et les résidents cliniques qui veulent devenir neuropathologistes et neurologues.
CLASSIFICATION DES VOIES
Les voies du système nerveux central sont un système de fibres nerveuses qui relient divers départements le cerveau et la moelle épinière, à la fois entre eux et à l'intérieur soit du cerveau soit de la moelle épinière, fournissant des fonctions bilatérales
communication entre les différentes structures cérébrales. Grace à
voies, l'activité intégrative du système nerveux central est atteinte
système, l'unité de l'organisme et sa connexion avec l'environnement extérieur.
Sections d'arcs réflexes multineuronaux complexes, représentés par
les chaînes de neurones le long desquelles un influx nerveux suit une direction strictement définie sont considérées comme des voies du système nerveux central.
Les voies conductrices sont généralement divisées en trois groupes : la projection, qui
missural et associatif.
Selon la direction de l'impulsion, la projection
les voies sont divisées en afférentes (centripètes) et efférentes
nye (centrifuge). Les voies afférentes conduisent les impulsions nerveuses des récepteurs aux centres du cerveau et de la moelle épinière. Pu- efférent
ti - conduit les impulsions des centres du cerveau et de la moelle épinière au travail
quels organes.
Les voies commissurales relient des parties du cortex droit
ème et hémisphères cérébraux gauche.
Les chemins associatifs peuvent être définis comme des chaînes de neu-
rons (dans un hémisphère du cerveau) reliant divers centres du système nerveux et unissant ainsi
voies afférentes et efférentes dans l'arc réflexe.
Dans l'ontogenèse, les voies de projection se développent initialement, oui
Les prochaines - commissurales et les plus récentes - voies associatives.
Dans cet ordre, nous les considérerons dans ce tutoriel.
VOIES DE PROJECTION
Ces voies assurent une connexion bidirectionnelle du cortex cérébral avec les noyaux du tronc cérébral et les noyaux de la moelle épinière.
Les voies de projection sont divisées en afférentes (sensorielles) et
efférent (moteur). Fonctionnellement, ils pré-
constituent un tout unique, car ils sont les maillons d'un réseau complexe.
arc de conférence. Mais en raison de la complexité de la structure, ces liens sont considérés
séparément en voies afférentes et efférentes.
VOIES AFFERENTES
Ce sont des voies sensorielles par lesquelles s'effectue la projection de la surface du corps, les organes internes, muscles, articulations dans les sens
centres stimulants et moteurs du cortex.
De par la nature des impulsions conduites, les voies afférentes
sont divisés en trois groupes.
JE. Voies extéroceptives - transportent les impulsions (douleur, température-
nye, tactile, pression), résultant de l'impact
environnement externe sur la peau.
II. Voies proprioceptives - conduisent les impulsions des organes du mouvement
nia (muscles, tendons, capsules articulaires, ligaments), véhiculent des informations
théorie sur la position des parties du corps dans l'espace.
III. Voies intéroceptives - conduisent les impulsions des organes internes
les nouveaux vaisseaux, où les chimio-, baro- et mécanorécepteurs perçoivent l'état de l'environnement interne du corps, l'intensité du métabolisme, la chimie du sang et de la lymphe et la pression dans les vaisseaux.
Quelques régularités dans la structure des voies afférentes de projection.
Le début de chaque voie est représenté par des récepteurs situés dans la peau, les tissus sous-cutanés ou les parties profondes du corps.
Le premier neurone de toutes les voies afférentes est en dehors du centre
système nerveux central, dans les ganglions spinaux.
Le deuxième neurone est localisé dans les noyaux de la partie spinale ou souche du cerveau.
Toutes les voies ascendantes passent par le tegmentum du tronc cérébral.
Le troisième neurone dans les voies menant au cortex des hémisphères de la douleur
cerveau, est situé dans les noyaux du thalamus et dans le cervelet
tei - dans le cortex cérébelleux.
Les chemins qui amènent les impulsions au cortex cérébral ont une décussation faite par des processus 2e neurone ; grâce à ça-
mu chaque moitié du corps est projetée sur l'opposé
pou du cerveau.
Les voies cérébelleuses soit n'ont pas une seule décussation, soit
sont baptisés deux fois, de sorte que chaque moitié du corps est projetée sur le cortex de la même moitié du cervelet.
Voies reliant le cervelet au cortex cérébral
sont franchis.
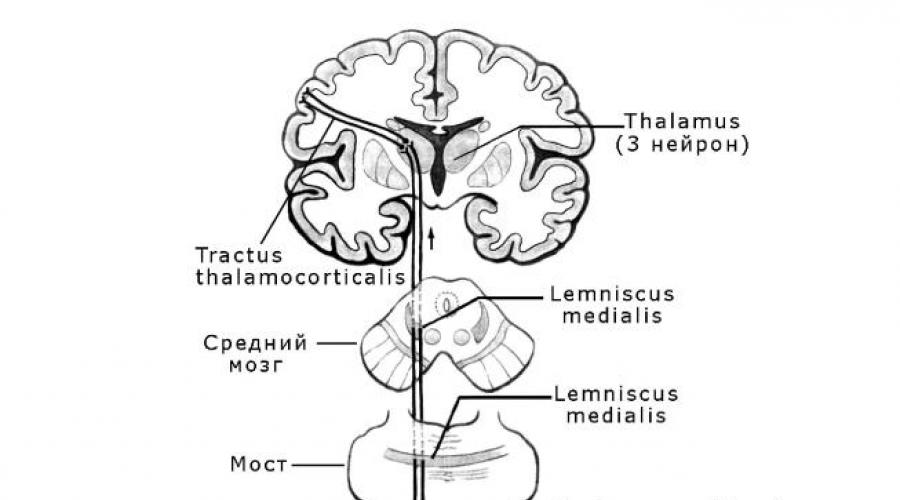
La voie conductrice de la sensibilité proprioceptive (profonde)
ness (Tractus ganglio-bulbo-thalamo-corticalis)
Cette voie conduit la sensation musculo-articulaire consciente à partir des propriorécepteurs de l'appareil de mouvement. Phylogénétiquement, c'est le plus jeune. Avec perte de sensation articulaire-musculaire -
le patient perd une idée de la position des parties du corps dans l'espace
ve, ne peut pas déterminer la direction du mouvement des membres ; dans la douleur-
e il y a violation de la coordination des mouvements : disproportionné, disproportionné
mouvements coordonnés, démarche instable.
Le chemin des impulsions proprioceptives conscientes est à trois neurones.
ny (Fig. 1). Il est représenté par trois faisceaux consécutifs : tractus gangliobulbaris, tractus bulbothalamicus, tractus thalamocorticalis.

Riz. 1. Voies de la sensibilité proprioceptive
direction corticale.
Les premiers neurones sont représentés par des cellules pseudo-unipolaires,
dont les corps sont situés dans les nœuds rachidiens. Les dendrites cellulaires sont envoyées à la périphérie dans le cadre des nerfs rachidiens et se terminent
propriorécepteurs dans les os, le périoste, les ligaments et la capsule des articulations, des tendons et des muscles. Axones - les processus centraux du pseudo-
les cellules pré-unipolaires, dans le cadre des racines postérieures, pénètrent dans la moelle épinière segment par segment, sans pénétrer dans la matière grise, montent - à
la composition des cordons postérieurs de la moelle épinière, formant des faisceaux des cordons postérieurs
kov : faisceau mince médialement situé de Gaulle (fasciculus gracilis) et
latéralement - faisceau en forme de coin de Burdakh (fasciculus cuneatus). Le faisceau de Gaulle conduit des impulsions proprioceptives conscientes de membres inférieurs et moitié inférieure torse partie concernée, à
il convient aux fibres de 19 nœuds rachidiens : 1 coccygien, 5
sacrée, 5 lombaire, 8 thoracique inférieure. Le faisceau de Burdakh conduit une profonde sensation articulaire-musculaire de la partie supérieure du torse, du cou,
membres supérieurs. Il comprend les fibres de 12 ganglions rachidiens : 4 thoraciques supérieurs et 8 cervicaux.
Les fibres des cordons postérieurs sont disposées en couches. Le plus moi-
dialy (plus près du sillon médian postérieur) sont des fibres adjacentes provenant des nœuds rachidiens sacrés. Les faisceaux de Gaulle et de Burdach atteignent le bulbe rachidien sans s'interrompre dans la moelle épinière.
Dans le bulbe rachidien, les fibres des cordons postérieurs se rapprochent du poison
cadres: nucleus gracilis et nucleus cuneatus, situés dans les tubercules du même nom, et ici ils passent aux deuxièmes neurones. Les premiers neurones constituent le chemin - tractus gangliobulbaris.
Les axones des seconds neurones, dont les corps sont situés dans le noyau gracilis et le noyau cunéiforme du bulbe rachidien, passent au contre-
le côté opposé, formant une décussation dorsale sensible ou décussation de la boucle médiale - decussatio lemniscorum. Ces fibres sont
pleurs partie intégrante boucle médiale - lemniscus medialis, passe dans le bulbe rachidien dorsal aux pyramides, puis à travers la partie dorsale du pont et le tegmentum du mésencéphale. Fibres des seconds neurones, dos-
thalamus, approchez-vous des noyaux ventrolatéraux du bu-
gra, où ils passent aux corps des troisièmes neurones. Dans la région du pont, une boucle vertébrale relie ces fibres (chemins des sens de la peau)
membres, tronc et cou), ainsi que l'anse trijumeau
(sensibilité cutanée et proprioceptive du visage, qui conduit
fibre nerf trijumeau). Les deuxièmes neurones constituent le chemin
tractus bulbothalamicus.
Une partie des axones des seconds neurones (cellules des noyaux minces et en forme de coin) à travers les jambes inférieures du cervelet atteignent le cortex de ses hémisphères sur leurs propres côtés et opposés. Ainsi, le cervelet reçoit des impulsions proprioceptives, grâce auxquelles il participe à la coordination des mouvements.
Les fibres des troisièmes neurones, dont les corps sont situés dans le ventro-
les noyaux latéraux du thalamus traversent la partie médiane de la jambe postérieure de la capsule interne, puis, dans le cadre de la couronne radiante, ils s'approchent du gyrus précentral de l'hémisphère cérébral, où ils se terminent
sont situés sur les cellules de la quatrième couche du cortex. Les troisièmes neurones constituent le trajet tractus thalamocorticalis. Dans le tiers supérieur du gyrus précentral de la
c'est des impulsions proprioceptives conscientes du membre inférieur et de la même moitié du corps ; dans le tiers médian du gyrus - du membre supérieur; dans le tiers inférieur - de la tête. Étant donné que les fibres sont
e tract dans la région de la moelle allongée faire une croix (re-
croix de l'anse médiale), puis dans le gyrus précentral droit
la lusaria du gros cerveau reçoit des impulsions de la moitié gauche du corps ;
de la moitié droite du corps - au gyrus précentral de l'hémisphère gauche -
Des muscles de la tête, de la capsule et des ligaments de l'articulation temporo-mandibulaire
les impulsions proprioceptives conscientes de tava sont effectuées le long des fibres
nous les nerfs trijumeau et glossopharyngien. Ces voies sont à trois neurones.
Les corps des premiers neurones sont situés dans les nœuds sensitifs indiqués
nerfs crâniens. Les corps des seconds neurones sont des cellules sensorielles
noyaux des nerfs crâniens dans la région du tronc cérébral. Les axones sont auto-
ry neurones passent du côté opposé et, dans le cadre du tractus nucleothalamicus, atteignent les noyaux ventrolatéraux du thalamus, où se trouvent les corps des troisièmes neurones. À partir des corps des troisièmes neurones du thalamo-
le tractus cortical (tractus thalamocorticalis) est dirigé par le milieu
La plupart des fibres de l'écorce du ver sont croisées.
Riz. 5. Voie spino-cérébelleuse postérieure (Flexiga).
Les fibres du tractus cérébelleux spinal postérieur ne se croisent pas dans la région de la moelle épinière et du bulbe rachidien, c'est pourquoi cette voie est appelée voie cérébelleuse directe ou non croisée.
Différences structurelles entre les deux voies cérébelleuses spinales
partagent leurs différences fonctionnelles. On pense que le long de la voie postérieure, le cervelet reçoit des informations des récepteurs musculaires et tendineux.
le fossé de chaque groupe musculaire isolé. Les signaux, provenant apparemment de grands groupes musculaires, pénètrent dans le cervelet par la voie antérieure.
Au deuxième type de connexions " système musculo-squelettique- moelle épinière - cervelet "la commutation sur les noyaux du tronc cérébral comprend
fibres arquées externes.
Dans le cadre des jambes inférieures au cortex des hémisphères cérébelleux passent
fibres ticulo-cérébelleuses qui ferment la moelle épinière
connexions réticulo-cérébelleuses. On suppose que le cervelet reçoit des informations proprioceptives supplémentaires sur le mouvement à travers eux.
yah engagé les muscles squelettiques membres. De plus, reti-
les fibres culo-cérébelleuses transmettent les signaux de la zone motrice du cortex des hémisphères cérébraux, c'est-à-dire qu'elles constituent le dernier segment des connexions cortico-réticulaires-cérébelleuses.
La prochaine "entrée" des signaux afférents au cervelet est l'olivo-
la voie cérébelleuse, qui pénètre par les pédoncules cérébelleux inférieurs,
décussant et se terminant sur les cellules de Purkinje du cortex cérébelleux. Le noyau d'olive est considéré comme l'un des plus importants "pré-
noyaux cérébelleux. On pense qu'à travers le noyau de l'olive et l'olive-
voie cérébelleuse Le cervelet reçoit des signaux du cortex cérébral, ext-
système rapyamidal et informations proprioceptives du segment
appareil de tare de la moelle épinière.
Le long des voies provenant des noyaux vestibulaires (le noyau vestibulaire
mais-fibres cérébelleuses) à travers les jambes inférieures du cervelet jusqu'à sa partie la plus ancienne, le "système floculo-nodulaire", le cervelet reçoit des informations des récepteurs du labyrinthe, c'est-à-dire des informations sur les forces du
racines, sur la position de la tête dans l'espace.
Les principales connexions du cortex cérébral avec le cervelet sont
traversent leurs propres noyaux de pont. Pour eux de l'écorce de toutes les actions de la semi-
les charias sont dirigées par des chemins de pont corticaux. De plus, depuis les pyramides
chemins qui courent à la base du pont entre ses propres poisons
ramie, les collatéraux se ramifient vers les cellules des noyaux. Les axones des cellules possèdent
les noyaux veineux du pont sont croisés et forment un cérébellopontin
conduit, qui est anatomiquement représenté par le cervelet moyen
jambes, ses fibres se terminent sur les cellules du cortex des hémisphères
cervelet.
Ayant reçu par ses voies afférentes des informations sur l'état et le tonus des muscles, sur la position des parties du corps, sur l'action d'un certain nombre de facteurs environnementaux, ainsi que sur l'acte moteur proposé, le cerveau
le starter le traite et corrige en conséquence le mouvement émergent afin qu'un mouvement optimal et coordonné au maximum soit formé.
acte moteur dynated. Une telle coordination est obtenue grâce aux signaux provenant du cervelet le long de ses connexions efférentes.
zyam aux centres moteurs du cerveau et de la moelle épinière.
Les principales structures intégratives du cervelet sont
neurones shevid (cellules de Purkinje), qui donnent naissance à des efférents
fibres du cortex cérébelleux et sont classiquement pris comme le 3ème neurone des voies cérébelleuses. Les axones des neurones en forme de poire vont aux noyaux du cervelet, où se trouve le 4ème neurone des voies cérébelleuses. À partir du noyau de la tente, commence le chemin cérébelleux-nucléaire efférent de projection, les fibres de ce chemin traversent le pédoncule cérébelleux inférieur et
chivayutsya dans les noyaux moteurs des nerfs crâniens et les noyaux de la formation réticulaire.
Les voies efférentes les plus importantes du cervelet proviennent du noyau denté. Voie cérébelleuse-tegmentale (tractus cerebellotegmentalis) passe dans le pédoncule cérébelleux supérieur et, traversant,
se termine par le noyau rouge, d'où le noyau rouge
tractus rachidien, qui, avec le réticulaire-
voie spinale, principale voie descendante du système extrapyramidal (le trajet de ces voies est décrit dans la section sur l'extrapyramidal
à mi-chemin). Ces chemins se terminent sur les cellules motrices du spin-
cerveau de la jambe. Ainsi, les voies cérébelleuses afférentes et efférentes assurent la régulation des mouvements au niveau du tronc cérébral.
cerveau rusé.
Voies de l'analyseur intéroceptif
Le terme « intérocepteurs » a été proposé pour la première fois en 1906 par C. Sher-
sonnerie pour désigner les récepteurs tube digestif, sp-
cialisé sur la perception des stimuli chimiques.
Tous les organes internes (relatifs à l'appareil digestif, respiratoire
corps, systèmes génito-urinaires), ainsi que les parois des vaisseaux sanguins contiennent une variété de terminaisons nerveuses sensorielles appelées interorécepteurs. Certains interorécepteurs ne provoquent normalement pas de sensations. D'autres interorécepteurs permettent de ressentir
soif, faim, envie d'uriner, etc. Les récepteurs sont très divers et portent une charge fonctionnelle différente. À
les dépendances à ce sujet sont distinguées :
chémorécepteurs - percevant les irritations chimiques;
mécanorécepteurs - percevant le degré d'étirement mécanique des parois des vaisseaux sanguins, des organes internes;
thermorécepteurs.
À partir des récepteurs des organes internes, le flux d'impulsions est dirigé vers le cortex cérébral. Chemins conducteurs de l'analyseur d'organes internes
nouvelle passe dans le cadre des nerfs crâniens V, VII, IX, X ou dans le dos
les racines de la moelle épinière.
Des organes internes de la tête, du cou, de la cavité thoracique et en partie des organes abdominaux, les voies afférentes passent dans le cadre de l'indication
nerfs crâniens. Les corps des premiers neurones (cellules pseudo-unipolaires) sont situés dans les nœuds sensitifs des nerfs crâniens. Péri-
des processus sphériques dans la composition des nerfs correspondants se rapprochent des organes internes. Les processus centraux faisant partie des racines des nerfs crâniens V, VII, IX, X pénètrent dans le tronc cérébral. De plus, les fibres de cette voie se rapprochent des neurones des noyaux suivants du tronc cérébral, qui sont les corps des seconds neurones :
1) noyau spinal du nerf trijumeau (noyau spinal);
2) le noyau d'un chemin solitaire (noyau solitarius) - VII, IX, X paires de nerfs crâniens.
Les axones des seconds neurones passent du côté opposé et, dans le cadre de la boucle médiale, atteignent le thalamus. Les corps du troisième neuro-
nouveau sont représentés par les cellules des noyaux du thalamus. Leurs axones forment
elles forment un faisceau de fibres qui traverse le tiers postérieur de la patte postérieure de la capsule interne et se termine sur les neurones du noyau cortical de l'analytique.
congestion des organes internes - dans la partie inférieure de la post-centrale et pré-
gyrus central.
Une partie des voies afférentes des organes abdominaux et afférentes
les voies de déchirure des organes pelviens font partie des grands et petits nerfs splanchniques (nervi splanchnici major et minor) et des nerfs pelviens internes (nervi splanchnici pelvini).
Les corps des premiers neurones sont situés dans les nœuds rachidiens
(cellules pseudo-unipolaires). Leurs axones sont inclus dans les racines postérieures
courir dans la moelle épinière et se terminer sur les corps des seconds neurones - dans
zone interstitielle de matière grise de la moelle épinière.
Les axones des corps des seconds neurones traversent les cordons postérieur et latéral.
kah ; ils rejoignent principalement le tractus spinothalamicus et atteignent
yut noyaux du thalamus, où se trouvent les corps des troisièmes neurones. Leurs neurites se rapprochent du noyau cortical de l'analyseur d'organes internes.
L'innervation afférente des organes internes n'a pas de segment
caractère conteneur ; il a de nombreuses voies de sens interne-
vations. Les principales voies d'innervation proviennent des segments vertébraux les plus proches.
cerveau de la jambe. Autres moyens d'innervation des organes internes
proviennent de segments éloignés de la moelle épinière. De cette façon,
les informations des organes internes ne parviennent pas seulement au plus proche,
mais aussi dans des segments éloignés de la moelle épinière. Dans les lésions de la moelle épinière, les voies accessoires sont les seules voies afférentes des organes internes.
VOIES EFFÉRENTES
Voies de projection descendantes (effecteur, effet-
enzymatique) conduisent les impulsions du cortex, des centres sous-corticaux vers le bas
départements couchés, aux noyaux du tronc cérébral et aux noyaux moteurs de la moelle épinière.
Les voies efférentes sont divisées en deux groupes. Le premier groupe de
Ils forment des voies pyramidales qui commencent par des cellules pyramidales dans le cortex cérébral. Le deuxième groupe comprend extrapyramid-
voies nye (extrapyramidales) qui conduisent les impulsions des ganglions de la base.
Quelques régularités dans la structure des voies de projection efférentes.
Le premier neurone de toutes les voies efférentes est localisé dans le cortex cérébral (voies pyramidales), le cortex cérébelleux, les noyaux sous-corticaux (système extrapyramidal).
Les voies de projection efférentes occupent la jambe antérieure, qui
leno et la partie antérieure de la jambe postérieure de la capsule interne, passant
dyat à la base des jambes du cerveau et du pont.
Toutes les voies efférentes se terminent dans les noyaux moteurs de l'humain
nerfs spinaux et dans les cornes antérieures de la moelle épinière, où
etsya dernier, neurone moteur.
Axones des motoneurones des cornes antérieures de la moelle épinière
ils partent de la moelle épinière dans le cadre de ses racines antérieures, puis dans le cadre des nerfs spinaux et de leurs branches, dans le cadre de leurs branches, ils atteignent les muscles squelettiques.
Les chemins efférents forment une décussation complète ou partielle,
à la suite de quoi les impulsions du cortex cérébral ne sont pas
sont transmis aux muscles de la moitié opposée du corps.
À Les fibres des voies efférentes suivantes correspondent aux motoneurones des cornes antérieures de la moelle épinière :
1) voie cortico-spinale antérieure (pyramidale) ;
2) voie latérale cortico-spinale (pyramidale) ;
3) spinale nucléaire rouge façons;
4) tectospinal façons;
5) olivo-spinal façon.
Au cours du processus de phylogenèse, des voies efférentes se sont d'abord formées pour conduire les impulsions du cervelet. Ce sont les plus anciens
leur. Plus tard, des voies provenant du tegmentum du mésencéphale se sont développées. Les plus jeunes sont les allées pyramidales, qui forment
sont réconciliés dans le cadre du développement du néocortex des hémisphères cérébraux.
VOIES PYRAMIDIQUES
Les voies pyramidales exercent un contrôle conscient du squelette
muscles de noé ; servent à effectuer des mouvements très différenciés.
Les voies pyramidales ont atteint leur plus haut développement chez l'homme. Ce sont les voies efférentes les plus jeunes.
Les fibres des voies pyramidales sont dirigées des cellules pyramidales du cortex cérébral vers les neurones des noyaux moteurs du crâne
nerfs ou cornes antérieures de la moelle épinière. En fonction de cela-
Les voies pyramidales sont divisées en voies cortico-spinales (antérieure et latérale) et en voie cortico-nucléaire. Ces voies et d'autres commencent par de grandes cellules pyramidales, y compris les cellules pyramidales géantes de Betz, dont les corps sont situés dans la cinquième couche du cortex du précentral
gyrus des hémisphères cérébraux. Selon certaines études
Les voies pyramidales commencent également dans le cortex du tiers postérieur des gyri frontal supérieur et moyen, du tiers antérieur du lobule pariétal supérieur et du gyrus supramarginal de l'hémisphère cérébral.
Les voies pyramidales sont caractérisées par quelques caractéristiques communes, un
1) les premiers neurones des voies sont de grosses cellules pyramidales
ki, (y compris les cellules pyramidales géantes de Betz), dont les corps
certains sont situés dans la cinquième couche du cortex cérébral ;
2) dans la région des hémisphères cérébraux, les trajets pyramidaux se succèdent dans le cadre de la couronne rayonnante puis de la capsule interne, occupant le genou et les deux tiers antérieurs de la jambe postérieure de cette dernière ;
3) dans le tronc cérébral, les voies pyramidales traversent séquentiellement
couper les jambes du cerveau, pont, bulbe rachidien, veineux situé
4) à la frontière du bulbe rachidien et de la moelle épinière, 80% des fibres corticales
dans le tractus rachidien passe du côté opposé
bien, formant une croix de pyramides;
5) dans la moelle épinière, les faisceaux pyramidaux se succèdent dans les cordons antérieurs
(tractus cortico-spinal antérieur) et dans les cordons latéraux de la moelle épinière (tractus cortico-spinal latéral);
6) les seconds neurones des voies pyramidales sont représentés par des cellules du
noyaux ventraux des nerfs crâniens (pour la voie cortico-nucléaire)
ou cellules des noyaux moteurs des cornes antérieures de la substance grise
va de la moelle épinière (pour le trajet cortico-spinal);
7) axones des corps des seconds neurones dans le crâne ou la colonne vertébrale
les nerfs latéraux atteignent les muscles squelettiques, où ils se terminent
sont des effecteurs.
Tractus cortico-spinal
(Tractus corticospinalis)
Le tractus cortico-spinal (Fig. 6) conduit l'influx nerveux des cellules pyramidales de la couche V du cortex des hémisphères cérébraux vers le neuro-
nous les noyaux moteurs des cornes antérieures de la matière grise de la moelle épinière.

Riz. 6. Voies pyramidales (voies cortico-spinales).
Les axones des premiers neurones de la voie pyramidale font partie des rayons
couronne, puis passer entre le thalamus et le lenticulaire
noyau à la capsule interne, où ils passent de manière compacte, occupant
les deux tiers médians de la patte postérieure de la capsule interne. Dans le capuchon intérieur-
sule plus près de son genou se trouvent des fibres qui transmettent les impulsions aux muscles du membre supérieur, derrière elles se trouvent des fibres pour les muscles du tronc, puis - des fibres pour les muscles du membre inférieur. Puis dans-
les lobes du trajet suivent le milieu de la base des jambes du cerveau dans la partie ventrale
nouvelle partie du pont. Dans la région du pont, les fibres de la moelle cortico-spinale
ils perdent leur compacité. Un grand nombre de petits faisceaux de cortex
dans le tractus rachidien sont séparés les uns des autres par leurs propres noyaux-
mi du pont et des fibres du tractus cérébellopontin.
Dans le bulbe rachidien, des faisceaux disparates se rejoignent et forment des pyramides (pyramis), qui font saillie dans les veines.
tral surface du cerveau sous la forme de crêtes longitudinales. Donc, pyramide-
dys, clairement visible sur la surface ventrale de la moelle allongée
ha, sont des amas de fibres de la cortico-moelle épinière-
A la frontière avec la moelle épinière, la plupart (80%) des pi-
chemin ramide passe du côté opposé, formant une pyramide
croix médiane (decussatio pyramidum). Ceci est "moteur" ou inférieur
ny décussation, contrairement au "sensible" - supérieur, formé par les fibres des boucles médiales. fibres du tractus cortico-spinal
qui ont traversé du côté opposé, passent du côté
natiques de la substance blanche de la moelle épinière et forment une cor-
tractus covo-spinal (tractus corticospinalis lateralis). Petite partie
(20%) des fibres restent couchées, descendent dans les cordons antérieurs
kah matière blanche de la moelle épinière, formant la corticale antérieure
moelle épinière (tractus corticospinalis antérieur).
Le tractus cortico-spinal latéral occupe la partie interne
partie postérieure du funicule latéral et descend dans la moelle épinière jusqu'à son département sacré. Cette voie se sépare de la surface de la moelle épinière
Xia avec une couche de fibres du tractus cérébelleux spinal postérieur. Comme suit-
dans la direction caudale, les fibres partent de ce tractus, qui
seigle fin sur les cellules des noyaux moteurs des cornes antérieures de chaque
segment long. Un nombre significativement plus grand de fibres est séparé du tractus dans la zone d'épaississement de la moelle épinière, où dans les segments de cette dernière il y a un plus grand nombre de neurones effecteurs, nécessaires
mine pour l'innervation de la masse musculaire des membres supérieurs et inférieurs. En bo-