Creatorul teoriei cromozomiale a eredității este un om de știință. Teoria cromozomală a eredității
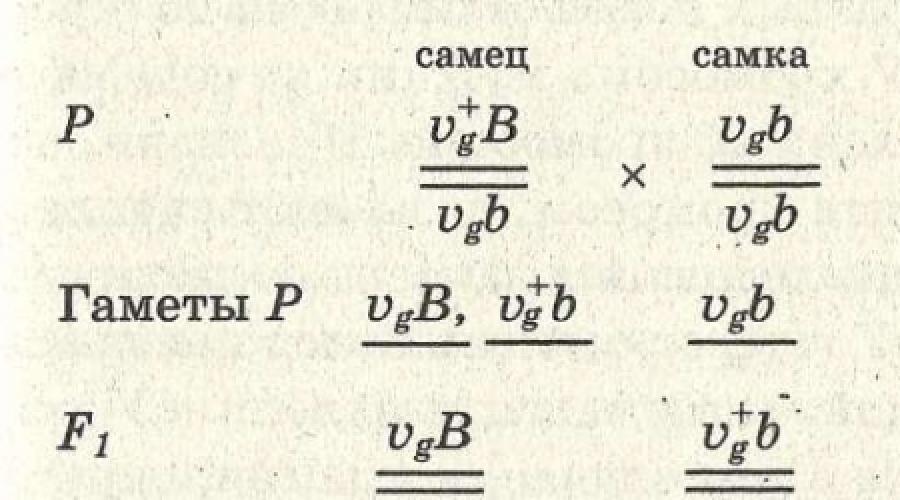
Citeste si
Celulele închise în nucleu sunt purtătoare de gene și reprezintă baza materială a eredității, adică. continuitatea proprietăților organismelor într-un număr de generații este determinată de continuitatea cromozomilor acestora. H. t. n. a apărut la începutul secolului al XX-lea. pe baza teoriei celulare și a utilizării analizei hibridologice pentru a studia proprietățile ereditare ale organismelor.
În 1902, W. Setton în SUA, care a atras atenția asupra paralelismului în comportamentul cromozomilor și așa-numitul mendelian. „factori ereditari”, iar T. Boveri din Germania a prezentat ipoteza cromozomială a eredității, conform căreia factorii ereditari mendeliani (numiți mai târziu gene) sunt localizați în cromozomi. Prima confirmare a acestei ipoteze a fost obținută în studiul mecanismului genetic de determinare a sexului la animale, când s-a constatat că acest mecanism se bazează pe distribuția cromozomilor sexuali în rândul descendenților. Fundamentarea suplimentară a H. t. aparține geneticianului american T. H. Morgan, care a observat că transferul anumitor gene (de exemplu, gena care provoacă Drosophila femela cu ochi albi atunci când este încrucișată cu masculi cu ochi roșii) este asociat cu transferul cromozomului X sexual, adică că trăsături legate de sex (o persoană are câteva zeci de astfel de semne, inclusiv unele defecte ereditare - daltonism, hemofilie etc.).
Dovada lui H. t. a fost obținut în 1913 de geneticianul american C. Bridges, care a descoperit nondisjuncția cromozomală în timpul meiozei la femelele Drosophila și a observat că o perturbare în distribuția cromozomilor sexuali este însoțită de modificări în moștenirea trăsăturilor legate de sex.
Odată cu dezvoltarea lui H. t. s-a constatat că genele situate pe același cromozom constituie un grup de legătură și trebuie moștenite împreună; numărul de grupuri de legătură este egal cu numărul de perechi de cromozomi, constant pentru fiecare tip de organism ; trăsăturile care depind de genele legate sunt, de asemenea, moștenite împreună. Drept urmare, legea combinației independente de caracteristici ar trebui să aibă o aplicare limitată; trăsăturile ale căror gene sunt localizate pe cromozomi diferiți (neomologi) trebuie moștenite în mod independent. Fenomenul legăturii incomplete a genelor (când, împreună cu combinațiile parentale de trăsături, se găsesc combinații noi, recombinante ale acestora în urmașii încrucișărilor) a fost studiat în detaliu de Morgan și colegii săi (A. G. Sturtevant și alții) și a servit ca un justificare pentru aranjarea liniară a genelor în cromozomi. Morgan a sugerat că genele legate ale cromozomilor omologi, care sunt în combinații și în părinți, pot schimba locul în timpul meiozei în forma heterozigotă ®, ca urmare a căreia, împreună cu gameții AB și ab, se formează gameții Ab și aB. Asemenea recombinări apar din cauza rupturilor cromozomilor omologi în zona dintre gene și a conexiunii ulterioare a capetelor rupte într-o nouă combinație: realitatea acestui proces, numit încrucișarea cromozomilor, sau crossing over, a fost dovedită în 1933 de omul de știință K. Stern în experimente cu Drosophila și oamenii de știință americani X. Creightonomi B. McClintock - cu porumb. Cu cât genele legate sunt mai îndepărtate, cu atât este mai probabil ca acestea să se încrucișeze. Dependența frecvenței de încrucișare de distanțele dintre genele legate a fost folosită pentru a construi hărți genetice ale cromozomilor. În anii 30. Secolului 20 F. Dobzhansky a arătat că ordinea de plasare a genelor pe hărțile genetice și citologice ale cromozomilor coincide.
Conform ideilor școlii Morgan, genele sunt purtători discreti și indivizibili de informații ereditare. Cu toate acestea, descoperirea în 1925 de către oamenii de știință sovietici G. A. Nadson și G. S. Filippov, și în 1927 de către omul de știință american G. Möller, a influenței razelor X asupra apariției modificărilor ereditare (mutații) la Drosophila, precum și utilizarea razelor X pentru a accelera procesul de mutație la Drosophila În 1928–30, oamenii de știință sovietici A. S. Serebrovsky, N. P. Dubinin și alții au făcut posibilă formularea de idei despre divizibilitatea unei gene în unități mai mici dispuse într-o secvență liniară și capabile. a modificărilor mutaționale. În 1957, aceste idei au fost dovedite prin munca savantului american S. Benzer cu bacteriofagul T4. Utilizarea razelor X pentru a stimula rearanjamentele cromozomiale a făcut posibil ca N. P. Dubinin și B. N. Sidorov să descopere în 1934 efectul poziției unei gene (descoperit în 1925 de Sturtevant), adică dependența manifestării unei gene de localizarea acestuia pe cromozom. A existat o idee a unității discretității și continuității în structura cromozomului.
H. t. n. se dezvoltă în direcția aprofundării cunoștințelor despre purtătorii universali de informații ereditare - molecule de acid dezoxiribonucleic (ADN). S-a stabilit că o secvență continuă de baze purinice și pirimidinice de-a lungul lanțului ADN formează gene, intervale intergene, semne ale începutului și sfârșitului citirii informațiilor în cadrul unei gene; determină natura ereditară a sintezei proteinelor celulare specifice și, în consecință, natura ereditară a metabolismului. ADN-ul formează baza materială a grupului de legătură în bacterii și mulți viruși (în unele viruși, acidul ribonucleic este purtătorul de informații ereditare) ; Moleculele de ADN care alcătuiesc mitocondriile, plastidele și alte organele celulare servesc ca purtători materiale ai eredității citoplasmatice.
H. t. n., explicând modelele de moștenire a trăsăturilor la animale și organismele vegetale, joacă un rol important în pagina - x. știință și practică. Ea echipează crescătorii cu metode de reproducere a raselor de animale și a soiurilor de plante cu proprietățile dorite. Unele prevederi ale H. t. permite să conducă mai rațional pagina - x. producție. Deci, fenomenul moștenirii unui număr de semne legate de un etaj la pagina - x. animale permise înainte de inventarea metodelor de reglare artificială a sexului în viermi de mătase sacrificați coconii de sexul mai puțin productiv, înainte de a dezvolta o metodă de separare a găinilor după sex prin examinarea cloacii - sacrificarea cocoșilor etc. Valoarea majoră pentru creșterea productivității multor pagini - x. culturile are utilizarea poliploidiei. Studiul bolilor ereditare umane se bazează pe cunoașterea tiparelor de rearanjamente cromozomiale.
modele, descoperite de școala Morgan și apoi confirmate și aprofundate pe numeroase obiecte, sunt cunoscute sub denumirea generală de teoria cromozomală a eredității.
Principalele sale prevederi sunt următoarele:
1. Genele sunt pe cromozomi; fiecare cromozom este un grup de gene de legătură; numărul de grupuri de legături din fiecare specie este egal cu numărul de perechi de cromozomi.
2. Fiecare genă din cromozom ocupă un anumit loc (locus); genele sunt dispuse liniar pe cromozomi.
3. Genele alelice sunt schimbate între cromozomi omologi.
4. Distanța dintre gene (loci) din cromozom este proporțională cu numărul de încrucișări între ele.
Fondatorul teoriei, Thomas Gent Morgan, un genetician american, laureat al Premiului Nobel, a prezentat o ipoteză despre limitarea legilor lui Mendel.
În experimentele sale, a folosit musca de fructe Drosophila, care are calități importante pentru experimentele genetice: lipsă de pretenții, fertilitate, un număr mic de cromozomi (patru perechi) și multe trăsături alternative distincte.
Morgan și studenții săi au stabilit următoarele:
- Genele situate pe același cromozom sunt moștenite împreună sau legate.
- Grupurile de gene situate pe același cromozom formează grupuri de legătură. Numărul de grupuri de legătură este egal cu setul haploid de cromozomi la indivizii homogametici și n + 1 la indivizii heterogametici.
- Între cromozomii omologi poate avea loc un schimb de situsuri (crossing over); ca urmare a încrucișării, apar gameți, ai căror cromozomi conțin noi combinații de gene.
- Frecvența de încrucișare între cromozomi omologi depinde de distanța dintre genele situate pe același cromozom. Cu cât această distanță este mai mare, cu atât frecvența de încrucișare este mai mare. Pentru o unitate de distanță dintre gene, se ia 1 morganid (1% din crossing over) sau procentul de apariție a indivizilor crossover. Cu o valoare a acestei valori de 10 morganide, se poate susține că frecvența încrucișării cromozomilor în punctele de localizare a acestor gene este de 10% și că noi combinații genetice vor fi dezvăluite la 10% din descendenți.
- Pentru a determina natura locației genelor în cromozomi și a determina frecvența de încrucișare între ele, se construiesc hărți genetice. Harta reflectă ordinea genelor de pe cromozom și distanța dintre genele de pe același cromozom. Aceste concluzii ale lui Morgan și ale colaboratorilor săi sunt numite teoria cromozomiala a ereditatii. Cele mai importante consecințe ale acestei teorii sunt ideile moderne despre genă ca unitate funcțională a eredității, divizibilitatea ei și capacitatea de a interacționa cu alte gene.
Exemplu de moștenire legată:
- Vg - aripi normale de Drosophila;
- vg - aripi rudimentare;
- BB - colorare gri corp;
- bb - culoarea întunecată a corpului.
Înregistrarea expresiei cromozomiale:
În acest caz, se respectă regula uniformității hibrizilor din prima generație. În conformitate cu a doua și a treia lege a lui Mendel, 25% din fiecare dintre fenotipurile posibile (muște gri, cu aripi lungi, muște gri cu aripi scurte, muște negre cu aripi lungi și muște negre cu aripi scurte) ar fi de așteptat în analiza ulterioară. cruci. Cu toate acestea, experimentele lui Morgan nu au dat astfel de rezultate. Când o femelă VgVgbb, recesivă în ambele trăsături, a fost încrucișată cu un mascul hibrid F1, s-au format 50% muște cenușii cu aripi scurte și 50% muște cu corp negru și aripi lungi:

Dacă o femelă dihibridă este încrucișată cu un mascul homozigot recesiv, se formează urmași: 41,5% - gri cu aripi scurte, 41,5% - negru cu aripi lungi, 8,5% - gri cu aripi lungi, 8,5% - negru cu aripi scurte. 
Aceste rezultate indică prezența legăturii genelor și încrucișarea între ele. Deoarece 17% dintre indivizii recombinanți au fost obținuți din a doua încrucișare, distanța dintre genele Vg și B este de 17%, sau 17 morganide.
moștenirea legată de sex
Seturile de cromozomi de diferite sexe diferă în structura cromozomilor sexuali. Cromozomul Y masculin nu conține multe dintre alelele găsite pe cromozomul X. Semnele determinate de genele cromozomilor sexuali se numesc legate de sex. Natura moștenirii depinde de distribuția cromozomilor în meioză. La sexele heterogametice, trăsăturile legate de cromozomul X și care nu au o alelă pe cromozomul Y apar chiar și atunci când gena care determină dezvoltarea acestor trăsături este recesivă. La om, cromozomul Y este transmis de la tată la fii, iar cromozomul X este transmis fiicelor. Copiii primesc al doilea cromozom de la mama lor. Este întotdeauna cromozomul X. Dacă mama poartă o genă recesivă patologică pe unul dintre cromozomii X (de exemplu, gena pentru daltonism sau hemofilie), dar ea însăși nu este bolnavă, atunci este purtătoare. Dacă această genă este transmisă fiilor, aceștia se pot naște cu această boală, deoarece nu există nicio alele pe cromozomul Y care suprimă gena patologică. Sexul organismului este determinat în momentul fertilizării și depinde de setul de cromozomi al zigotului rezultat. La păsări, femelele sunt heterogametice, iar masculii sunt homogametici. Albinele nu au cromozomi sexuali. Masculii sunt haploizi. Albinele sunt diploide.

Principalele prevederi ale teoriei cromozomiale a eredității:
- fiecare genă are un loc (loc) specific în cromozom;
- genele din cromozom sunt localizate într-o anumită secvență;
- genele unui cromozom sunt legate, prin urmare sunt moștenite în principal împreună;
- frecvența de încrucișare între gene este egală cu distanța dintre ele;
- setul de cromozomi din celulele unui anumit tip (cariotip) este o trăsătură caracteristică speciei.
Articol pentru concurs "bio/mol/text": 2015 marchează 100 de ani teoria cromozomiala a ereditatii. Principalele sale prevederi au fost formulate de T. Morgan, A. Sturtevant, G. Möller și K. Bridges în cartea „The Mechanism of Mendelian Inheritance”, publicată la New York în 1915. Iar mai târziu, Thomas Morgan a primit primul premiu Nobel „genetic” – pentru descoperirea rolului cromozomilor în ereditate. Conferința internațională „Chromosome 2015”, desfășurată în august 2015 în Novosibirsk Academgorodok, a fost dedicată aniversării teoriei cromozomilor. Următorul text este comentariile autorului poster despre istoria cercetării cromozomilor, prezentată la conferință, iar acum la „Biomolecule” - în cea mai „animată” nominalizare competitivă „ Vizual despre persoana iubită».
Notă!
Informații mai complete pot fi găsite în cartea - Koryakov D.E., Zhimulev I.F. . Novosibirsk: Editura Filialei Siberiei a Academiei Ruse de Științe, 2009 - 258 p., ISBN 978-5-7692-1045-7
Sponsorul nominalizării „Cel mai bun articol despre mecanismele îmbătrânirii și longevității” este Fundația Science for Life Extension. Premiul pentru alegerea publicului a fost sponsorizat de Helicon.
Sponsorii concursului: Laboratorul de soluții de imprimare 3D pentru cercetare în biotehnologie și Studioul de știință vizuală pentru grafică științifică, animație și modelare.
Textul de mai jos este un scurt comentariu asupra afișului, iar informații mai complete pot fi găsite în carte: Koryakov D.E., Zhimulev I.F. Cromozomii. Structură și funcții. Novosibirsk: Editura Filialei Siberiei a Academiei Ruse de Științe, 2009 - 258 p., ISBN 978-5-7692-1045-7.
Faceți clic pe imagine pentru a o mări (se deschide într-o fereastră separată).
Rolul genetic al cromozomilor
Fiecare organism reproduce numai felul său și chiar și în cele mai mici trăsături ale aspectului și comportamentului copiilor, se poate observa o asemănare cu părinții lor. Primul pas spre înțelegerea de ce se întâmplă acest lucru a fost făcut de un călugăr din orașul austriac Brunn (acum este cehul Brno) G. Mendel ( G. Mendel). În 1865, la o reuniune a Societății Brünn a Naturaliștilor, el a făcut un raport intitulat „ Experimente pe hibrizi de plante» ( Versuche über Pflanzen-Hybriden), iar în 1866 l-a publicat în colecția de lucrări a acestei societăți. Călugărul-naturalistul a descris rezultatele încrucișării diferitelor forme de mazăre și a sugerat prezența unor factori speciali de care depind semnele externe ale plantei. Modelele de moștenire ale acestor factori au fost numite ulterior legile lui Mendel. Cu toate acestea, contemporanii nu au înțeles semnificația acestei descoperiri și au uitat de ea și abia în 1900 G. de Vries ( H. de Vries, Olanda), K. Korrens ( C. Correns, Germania) și E. Chermak ( E. Tschermak, Austria) a redescoperit independent legile lui Mendel.
Cu mult înainte de toate aceste studii, care acum s-ar numi analiză genetică, oamenii de știință implicați în botanică, zoologie, embriologie, histologie și fiziologie au pus bazele citogenetica- știința cromozomilor. În diverse articole și cărți, se acordă prioritate descoperirii cromozomilor oameni diferiti, dar cel mai adesea anul descoperirii lor se numește 1882, iar descoperitorul lor este anatomistul german W. Flemming ( W. Flemming). Cu toate acestea, ar fi mai corect să spunem că nu a descoperit cromozomi, ci doar a colectat și simplificat în cartea sa fundamentală " Substanță celulară, nucleu și diviziune celulară» ( Zellsubstanz, Kern und Zellteilung) tot ce se știa despre ei în acel moment. Însuși termenul „cromozom” a fost introdus în știință de histologul german H. Waldeyer ( H. Waldeyer) în 1888, și tradus literal, termenul înseamnă „corp pictat”.
Acum este greu de spus cine a făcut prima descriere a cromozomilor. În 1842, botanistul elvețian K. Naegeli ( C. Nageli) a publicat o lucrare în care a descris câteva corpuri mici care apar în locul nucleului în timpul diviziunii celulare în timpul formării polenului în crini și tradescantia. Poate că acestea au fost primele desene ale cromozomilor. Prima (1873) descriere detaliată mitozăîntr-un vierme plat Mesostoma ehrenbergii aparține, după cum se spune, zoologului german A. Schneider ( FA. Schneider). El a descris nu doar stadiile individuale ale mitozei, care fuseseră văzute înaintea lui, ci întreaga secvență de modificări complexe ale nucleului: apariția corpurilor filamentoase în locul său, divergența lor în direcții opuse și formarea de noi nuclei în celulele fiice. . Un alt tip de împărțire este meioză- descris pentru prima dată în detaliu de E. van Beneden ( E. van Beneden, Belgia) în 1883, observând formarea gameților la viermi rotunzi. El a descoperit că în meioză numărul de cromozomi este redus la jumătate și restabilit în timpul fertilizării și, în ciuda diferenței de mărime, gameții masculini și feminini aduc un număr egal de cromozomi zigotului.
* - Câteva despre locul și scopul elementelor genetice mobile în genoamele pro și eucariote: " Elemente genetice mobile ale procariotelor: stratificarea „societății” vagabonzilor și a locuințelor», « Genomul uman: o carte utilă sau o revistă lucioasă?», « ADN-ul „junk” conduce evoluția mamiferelor?» - Ed.
O altă opțiune de schimb este schimbul de cromatide surori(SHO). Dacă cromatidele sunt schimbate în timpul încrucișării diferit cromozomii, apoi în cazul CHO, cromatidele sunt schimbate în interior unu cromozomii. Pentru prima dată, geneticianul american D. Taylor a văzut CHO ( J. Taylor) în 1958.
Încrucișarea, deși ambiguă, este asociată cu formarea în profaza de meioză a unei structuri speciale dintr-o pereche de cromozomi omologi - complex sinaptonemal. A fost descoperit în 1956 independent de doi citologi americani: M. Moses ( M. Moise) în raci și D. Focett ( D. Fawcett) într-un șoarece.
Diversitatea cromozomilor
Dacă înțelegem prin cromozomi orice purtător de informații ereditare, atunci aceștia sunt extrem de diversi ca mărime, formă, aspect, compoziție și număr. Cromozomii virusurilor și bacteriilor pot fi circulari sau liniari. Cromozomii cloroplastelor și mitocondriilor au formă circulară. Cromozomii nucleari ai eucariotelor au o formă liniară și sunt sub formă de corpuri X și V. forme diferite de obicei vin în minte când menționăm cromozomi. Ei sunt numiti, cunoscuti mitotică sau metafaza, deoarece au acest aspect în timpul diviziunii - mitoză (iar metafaza este una dintre etapele sale).
În 1912, botanistul și citologul rus S.G. Navashin a arătat că cromozomii de metafază au un set individual de caracteristici, inclusiv dimensiunea, raportul lungimii brațului, prezența sateliților și constricțiile. Folosind poziția centromerului sau raportul dintre lungimile brațelor, S.G. Navashin a propus o clasificare a cromozomilor mitotici, care este folosită și astăzi: metacentrici, submetacentrici, acrocentrici și telocentrici.
Numărul de cromozomi din tipuri diferite organisme pot varia foarte mult: de la două (într-o pereche de specii de plante și una dintre furnicile australiene) la 1440 într-o ferigă Ophioglossum reticulatumşi chiar 1600 în radiolariile marine Aulacantha scolymantha. La om, numărul de cromozomi este de 46 și a fost determinat abia în 1955 și publicat în 1956 de către citogeneticianul de origine chineză D. Chio ( J. Tjio) în colaborare cu liderul său A. Levan ( A.Levan) în Suedia. Câteva luni mai târziu, numărul a fost confirmat de britanicul C. Ford ( C. Ford) și D. Hamerton ( J. Hamerton). Au existat încercări de a determina numărul de cromozomi umani încă de la sfârșitul secolului al XIX-lea. În diferite cazuri, au existat sensuri diferite: 18, 24, 47 sau 48, - și abia în 1955 s-au convins că o persoană are cromozomi 46. În cinstea acestui eveniment, o placă comemorativă cu imaginea aceleiași plăci metafazice, conform căreia au fost numărați cromozomii. Este curios că numărul de cromozomi de cimpanzeu (48) a fost descoperit cu 15 ani mai devreme.
Este în general acceptat că numărul de cromozomi din fiecare tip de organism viu este constant, iar în marea majoritate a cazurilor este. Cu toate acestea, unele animale și plante au așa-numitele supranumerar, sau adiţional, cromozomi. Toți cromozomii setului principal sunt numiți Un cromozomi. Ele sunt mereu prezente, iar pierderea sau adăugarea chiar și a unuia dintre ele duce la consecințe grave. Cromozomii suplimentari se numesc cromozomii B, iar principalele lor caracteristici sunt prezența opțională și volatilitatea numărului. Pentru prima dată au fost găsiți cromozomi supranumerari de către E. Wilson ( E. Wilson, SUA) în 1906 la bug Metapodius terminalis.
Un tip deosebit de cromozom numit cromozomi perie de lampă, poate fi văzut în profaza primei diviziuni a meiozei în timpul formării ovocitelor la păsări, pești, reptile și amfibieni. Ele au fost menționate pentru prima dată în cartea sa fundamentală (1882) de V. Fleming, care a descoperit acești cromozomi la axolotl. Și-au primit numele de la asemănarea lor cu o perie pentru curățarea lămpilor cu kerosen.
Un loc cu totul special printre toate tipurile de cromozomi este ocupat de cromozomi politenici, care arată ca șnururi lungi și groase cu dungi transversale. Au fost descoperite de embriologul francez E. Balbiani ( E. balbiani) în 1881 în nucleii celulari ai glandelor salivare ale larvelor de țânțar Chironomus plumosus. Cromozomii politene au jucat un rol remarcabil în dezvoltarea geneticii, citogeneticii și biologiei moleculare. Cu ajutorul lor s-a demonstrat liniaritatea aranjamentului genelor și s-a dovedit fără echivoc rolul genetic al cromozomilor. Pe cromozomii politenici ai Drosophila a fost descris pentru prima dată polimorfismul cromozomial al populațiilor sălbatice. Pe cromozomii politenilor au fost descoperite genele pentru proteinele de șoc termic, componente ale unui sistem care protejează celulele tuturor organismelor de influențele stresante. Cromozomii politenilor au jucat un rol cheie în studiul sistemului de compensare a dozei la Drosophila.
Evoluția cromozomilor și a genomilor
În studiile citogenetice moderne, un rol important îl joacă colorație diferențială. Pentru prima dată, capacitatea cromozomilor de a se colora diferențial (adică inegală ca lungime) a fost demonstrată de britanicul S. Darlington ( C. Darlington) și L. La Cour ( L. La Cour) în 1938. O altă metodă importantă de cercetare este hibridizare in situ, care vă permite să determinați poziția oricărui fragment de ADN pe cromozom. Metoda se bazează pe abilitate acizi nucleici formează molecule dublu catenare, atât ADN-ADN, cât și ARN-ADN. Această metodă a fost inventată în 1969 de D. Goll ( J. Gall) și M. Pardue ( M. Pardue) din SUA și H. John ( H. Ioan), M. Birnstiel ( M. Birnstiel) și C. Jones ( K. Jones) din Marea Britanie.
Combinația acestor metode face posibilă studierea în detaliu a evoluției cromozomilor și genomilor*, iar însoțitorul constant al procesului evolutiv sunt rearanjamente cromozomiale. Pe măsură ce o specie evoluează, în cromozomii ei apar inevitabil rearanjamente care schimbă ordinea genelor în comparație cu specia ancestrală. Cu cât speciile se îndepărtează unele de altele, cu atât mai multe rearanjamente cromozomiale le deosebesc și cu atât ordinea genelor se schimbă mai mult. cunoscut tipuri diferite rearanjamente: deletii (pierdere), duplicari (dublare) si translocari (miscare) sectiunilor cromozomilor, care au fost descoperite de C. Bridges in 1916, 1919 si respectiv 1923. Un alt tip este inversiunile (rotația unui segment de cromozom cu 180 °), descrise de A. Sturtevant în 1921. În plus, există un tip special de rearanjare numită translocare Robertsoniană (sau fuziune centrică). A fost descris pentru prima dată de americanul W. Robertson ( W. Robertson) în 1916, comparând seturile de cromozomi ale speciilor de lăcuste strâns înrudite. Esența acestei rearanjamente se reduce la fuziunea a doi cromozomi acrocentrici într-unul metacentric sau submetacentric. Există și un proces invers - separarea centrică. În acest caz, cromozomul meta- sau submetacentric este împărțit în doi acrocentrici.
* - Pe biomoleculă, puteți găsi o selecție impresionantă de articole care într-un fel sau altul afectează evoluția genomului și modificările cod genetic: « Genomul viral în sistemul de evoluție», « Sub „armonica genică”», « Alopoliploidia sau modul în care genomi diferiți au învățat să trăiască sub același acoperiș», « Genomul complet al cintezelor din Galapagos a dezvăluit în sfârșit mecanismele evoluției lor», « Cum a fost compus genomul eucariot: endosimbioza VS. transfer orizontal continuu»; « Codul misterios al genomului nostru», « Evoluția codului genetic», « La originile codului genetic: spiritele înrudite», « Sinonime atât de diferite" si etc. - Ed.
Poziția cromozomilor în nucleu
ÎN sfârşitul XIX-lea secolul T. Boveri a propus ideea că cromozomii din nucleul de interfază nu sunt amestecați aleatoriu, ci fiecare dintre ei ocupă propriul spațiu. În 1909, el a inventat termenul „ teritoriul cromozomial". Prima dovadă a existenței teritoriilor cromozomiale a fost obținută abia în 1982 de către cercetătorul german T. Kremer ( T. Cremer) cu coautorii. Mai târziu, au vizualizat aceste teritorii folosind coloranți fluorescenți de diferite culori. S-a dovedit că cromozomii mari sunt mult mai probabil să fie găsiți în partea periferică a nucleului, în timp ce cei mici sunt concentrați în principal în partea centrală. În plus, la periferia nucleului, există regiuni de cromozomi care sunt epuizate în gene. Regiunile îmbogățite în gene, dimpotrivă, sunt situate mai aproape de centrul nucleului.
Compoziția cromozomilor. ADN
Cromozomii sunt structuri formate dintr-un complex complex de ADN, ARN și proteine. Un astfel de complex se numește cromatina.
ADN-ul ca substanță chimică a fost descoperit și izolat în formă pură tânărul cercetător elvețian F. Miescher ( F. Miescher), lucrând în 1868-1869 la Universitatea din orașul german Tübingen. El a studiat compoziție chimică leucocite, a căror sursă a fost puroiul din bandajele de la clinica chirurgicală locală. F. Miescher a dezvoltat o metodă de separare a nucleelor și citoplasmei celulelor și a analizat compoziția nucleelor. Pe lângă proteine și lipide, a descoperit o substanță pe care a numit-o nucleină(din cuvânt nucleu- nucleu), iar acum este cunoscut sub numele de ADN. Faptul că ADN-ul este purtătorul de informații ereditare a fost stabilit pentru prima dată în 1944 de americanii O. Avery ( O. Avery), C. MacLeod ( C. MacLeod) și M. McCarthy ( M. McCarty) în experimente privind infectarea șoarecilor cu pneumococi.
Structura moleculei de ADN sub forma unui dublu helix a fost descifrată în 1953 de F. Crick ( F. Crick), D. Watson ( J. Watson), M. Wilkins ( M. Wilkins) și R. Franklin ( R. Franklin) lucrează în Marea Britanie. Pentru această descoperire, primii trei cercetători au primit Premiul Nobel în 1962 (istoria descoperirii a fost descrisă în mod fascinant în cartea „ dublu helix» James Watson Foarte recomandat - Ed.). Rosalind Franklin nu se numără printre beneficiari, deoarece murise de cancer cu patru ani în urmă. Se știe că molecula de ADN constă dintr-o secvență de patru tipuri de nucleotide: adenină, timină, guanină și citozină*. Pentru dezvoltarea unei metode de determinare a secvenței lor ( secvențiere) în 1980, Premiul Nobel a fost acordat lui P. Berg ( P. Berg, SUA), W. Gilbert ( W. Gilbert, SUA) și F. Sanger ( F. Sanger, Marea Britanie).
* - Pe lângă cele patru nucleotide „clasice” din ADN, se găsesc și variantele lor modificate epigenetic: metilcitozină și metiladenina („ A șasea fundație ADN: de la descoperire la recunoaștere"). Și pentru unii bacteriofagi Bacillus subtilis descrie includerea în ADN a uracilului „ARN-nou” - roșu.
Dacă secvențierea a fost inițial proces laborios, care a făcut posibilă „citirea” doar a unui mic fragment la un moment dat, apoi, pe măsură ce tehnologia s-a dezvoltat, a devenit posibilă determinarea, de exemplu, a secvenței complete ADN mitocondrial bărbat (1981). În 1990, a fost lansat un proiect ambițios cu scopul de secvențiere completă a genomului uman, iar primul rezultat a fost prezentat în 2001 (biomolecula: „ Genomul uman: cum a fost și cum va fi"). În același timp, secvențierea unu genomul a costat o sumă colosală – sute de milioane de dolari. Dar tehnologia nu stă pe loc, iar apariția unor noi metode a redus costurile de mii de ori*. Secvențierea întregului genom este acum obișnuită, iar în 2009 a fost lansat proiectul Genome 10K. Scopul său este secvențierea și „asamblarea” completă a 10.000 de genomi de animale în cromozomi.
* - „Legea” lui Moore este de-a dreptul sortită să atingă punctele finale în diverse științe (oriunde a fost posibil să o tragă). Biologia a depășit chiar și electronica: scăderea treptată a costului de secvențiere în 2007 a intrat într-un vârf abrupt, aducând epoca citirii de rutină a genomului în stațiile rurale feldsher sub politicile MHI. Adevărat, în viitorul apropiat, mai trebuie să plătiți - 1000 de dolari plus costurile de transport: „ Tehnologie: 1.000 USD per genom". Dar chiar și acest lucru nu putea fi visat decât înainte de apariția noilor metode de secvențiere a ADN-ului: „ 454-secvențiere (pirosecvențiere ADN cu randament mare)". Și pentru a înțelege procesele de bază (la nivel celular) de dezvoltare a corpului și de victorie asupra cancerului, mai este ceva de visat: „ Secvențierea unei singure celule (versiunea - Metazoa)» - Ed.
Noile tehnologii au permis dezvoltarea unei astfel de direcții precum studiul ADN-ului antic (biomolecule: „ ADN antic: Bună ziua din trecut"). A devenit posibilă izolarea ADN-ului din oase vechi de zeci de mii de ani, iar în 2008, de exemplu, a fost secvențiat genomul mitocondrial de Neanderthal. Studiul ADN-ului antic și, într-adevăr, a întregii biologie moleculare moderne, nu poate fi imaginat fără utilizarea PCR - reacție în lanț a polimerazei. Pentru descoperirea sa, americanul K. Mullis ( K. Mullis) a primit Premiul Nobel în 1993.
Compoziția cromozomilor. Veverițe
ADN-ul din cromozomi suferă mai multe niveluri succesive de ambalare și chiar la primul nivel dublu helix ADN-ul se înfășoară în jurul unui glob de proteine nucleozom(biomolecula: " Roluri, rostogoliri la histona ADN"). Globul conține patru tipuri de proteine numite histonele. În 1982, biologul molecular englez A. Klug ( A. Klug) a primit Premiul Nobel pentru descifrarea structurii tridimensionale a nucleozomilor. Indirect, nucleozomii au primit un alt Premiu Nobel - în 1910 a fost primit de biochimistul german A. Kossel ( A. Kossel) pentru studiul compoziției chimice a substanțelor care formează nucleul celular, inclusiv descoperirea histonelor.
Părțile C-terminale ale moleculelor de histonă sunt strâns pliate, în timp ce părțile N-terminale nu au o structură definită și diverg liber în lateral. În 1963-1964, s-a descoperit că anumite reziduuri de aminoacizi din histone ar putea fi modificate covalent, adică acetilate sau metilate. Acum, lista modificărilor s-a extins semnificativ, atât grupe relativ simple - metil, acetil, fosfat - cât și molecule complexe mari: biotină, oligopeptide sau lanțuri de ADP-riboză pot fi atașate la resturi de aminoacizi. Modificările apar în principal pe părțile N- și, într-o măsură mult mai mică, pe părțile C-terminale ale moleculelor de histonă.
Conform teoria codului histonelor, modificările care sunt prezente pe nucleozomi într-o anumită regiune a cromatinei nu sunt aleatorii, ci „codifică” un anumit proces. Acest punct de vedere a fost formulat în 2000–2001 de B. Shtral ( B. Strahl, SUA), S. Ellis ( C. Allis, SUA) și T. Yenuwein ( T. Jenuwein, Austria). Schematic, procesul codului histonelor poate fi compus din trei etape. În prima etapă funcționează enzimele care modifică anumite reziduuri din histone. În a doua etapă, proteinele care au domenii speciale în acest scop se leagă de aminoacizii modificați. Fiecare dintre domenii este potrivit doar pentru modificarea „sa”. În ultima etapă, aceste proteine legate atrag alte complexe proteice, demarând astfel un fel de proces.
* - Despre perspective strălucitoare și îndoieli serioase în domeniul de aplicare a IPSC: " Cercetătorii francezi au reușit să întinerească celulele oamenilor centenari», « Bulgăre de zăpadă a problemelor cu pluripotență». - Ed.
Heterocromatina
Unul dintre obiectele de studiu ale diverselor procese epigenetice este heterocromatina. A fost descoperit ca regiuni mai întunecate ale cromozomilor în 1907 de către citologul german S. Guthertz ( S. Gutherz), iar termenii „heterocromatină” și „eucromatină” au fost introduși în 1928 de un alt citolog german E. Heitz ( E. Heitz). Pe scurt, eucromatina este părțile cromozomilor în care se află marea majoritate a genelor, în timp ce heterocromatina este în principal regiuni cu ADN necodificator, constând din secvențe scurte, repetate în mod multiplicat. În plus, eu- și heterocromatina diferă în timpul replicării în timpul fazei S a ciclului celular. Această diferență a fost descrisă pentru prima dată în 1959 de A. Lima de Faria ( A. Lima-de-Faria, SUA), investigând procesul de replicare a ADN-ului în testiculele lăcustelor Melanoplus diferentialis. El a arătat că heterocromatina începe și își termină replicarea ADN-ului mai târziu decât eucromatina.
O proprietate importantă a heterocromatinei este capacitatea de a inactiva genele eucromatinei plasate în ea. Acest fenomen se numește efect de poziție de tip mozaic. A fost descoperit în 1930 de G. Möller în Drosophila. Ca urmare a rearanjarii cromozomiale, gena alb a intrat în heterocromatina. Această genă este responsabilă pentru culoarea roșie a ochilor, iar dacă nu funcționează, atunci ochii devin albi. G. Möller, în schimb, a arătat muște ai căror ochi nu erau nici roșii, nici albi, ci pătați, iar la diferite muște petele erau de diferite forme și dimensiuni. Acest lucru se datorează faptului că gena în sine rămâne intactă, dar este inactivată doar aleatoriu în unele celule ale ochiului și funcționează în altele.
În ciuda multor ani de cercetare, procesul de formare a heterocromatinei este încă în mare parte neclar, în special în prima etapă. Se presupune că un proces similar cu interferența ARN(biomolecula: " Despre toți ARN-urile din lume, mari și mici"). Pentru descoperirea acestui fenomen, doi americani E. Fire ( Un foc) și K. Mello ( C. Mello) a primit Premiul Nobel în 2006. Procesul de interferență este complex și în mai multe etape, dar fără a intra în detalii, introducerea unui ARN dublu catenar omolog unei gene într-o celulă duce la inactivarea acestei gene.
Telomerii
Un studiu intensiv al telomerilor a început după americanii E. Blackburn în 1978 ( E. Blackburn) și D. Goll au secvențiat telomerul în ciliați Tetrahymena thermophila. S-a dovedit că telomerii conțin o secvență de șase nucleotide, repetată de 20 până la 70 de ori. În 1985 K. Greider ( C.greider) și E. Blackburn au descoperit o enzimă numită telomeraza, a cărui sarcină este să finalizeze construcția telomerilor. În 2009 E. Blackburn, K. Greider și D. Shostak ( J. Szostak, SUA) a primit Premiul Nobel pentru studiul telomerilor și descoperirea enzimei telomeraze (biomoleculă: „ Premiul Nobel „Fără vârstă”: în 2009, au fost recunoscute lucrările asupra telomerilor și telomerazei», « Îmbătrânirea - prețul de plătit pentru suprimarea tumorilor canceroase?»).
Compensarea dozei
Un număr imens de specii de organisme vii, inclusiv oamenii, au cromozomi sexuali neomologi, de exemplu, X și Y. Acest lucru necesită un proces numit compensarea dozei. Esența sa este următoarea: deoarece numărul de autozomi este același atât la bărbați, cât și la femei, numărul de gene autozomale și, prin urmare, numărul produselor lor, va fi, de asemenea, același. Dar numărul de produse sintetizate din gene situate pe cromozomul sexual va fi de 2 ori mai mare la un sex decât la celălalt. Se dovedește o disproporție care trebuie reglementată cumva, adică pentru a egaliza „doza de gene”. Sistemul de compensare a dozei este chemat pentru a rezolva această problemă (biomolecula: " , SUA) au prezentat ipoteza că la mamiferele femele unul dintre cei doi cromozomi X este inactivat, iar alegerea sa este aleatorie. În acest fel, sistemul de compensare a dozei la mamifere egalizează numărul de cromozomi X activi la sexe diferite: la masculi, există un singur cromozom X, iar la femele, din doi, doar unul funcționează.
La Drosophila, natura a inventat un alt mecanism, în esență opus celui al mamiferelor: singurul Cromozomul X masculin este hiperactivatși funcționează ca doi cromozomi X feminini. Faptul că activitatea totală a două copii ale oricărei gene din cromozomul X la femele și a unei copii la masculii de Drosophila este aceeași a fost descoperit în zorii dezvoltării geneticii. Acest lucru a fost făcut de K. Stern în 1929 și G. Möller în 1931, astfel încât Drosophila este primul organism în care a fost găsită compensarea dozei.
Și, în sfârșit...
Câteva cuvinte despre descoperire, care nu este direct legată de cromozomi, dar este folosită foarte activ, inclusiv pentru studierea diferitelor aspecte ale vieții cromozomilor. În 2008 O. Shimomura ( O. Shimomura), M. Chalfie ( M. Chalfie) și R. Tsien ( R. Tsien) din SUA a primit Premiul Nobel pentru descoperirea, izolarea și aplicarea proteină verde fluorescentă (GFP) meduze Aequorea victoria. Cu ajutorul manipulărilor moleculare, este posibil să se conecteze gena proteinei GFP cu gena oricărei alte proteine și să se obțină o proteină himerică care își va îndeplini atât funcția inițială, cât și strălucirea. în verde. Acest lucru face posibil să vedem în ce celule funcționează proteina, în nucleu sau citoplasmă, în ce părți ale cromozomilor. În plus față de verde (GFP), roșu (RFP) și galben (YFP) proteine fluorescente* sunt acum cunoscute.
* - Despre varietatea de proteine fluorescente și aplicarea lor în cercetare biologică materialele spun: Premiul Nobel pentru chimie fluorescent», « Proteine fluorescente: mai diverse decât credeți!», « „Hai să desenăm” o celulă vie". Și despre bioluminiscența în organismele terestre și marine și activitatea sistemului luciferină-luciferază - articole: „ Bioluminiscență: renaștere», « Strălucire microscopică la scară cosmică». - Ed.
Fiecare celulă conține un anumit număr de cromozomi. Există o mulțime de gene în ele. O persoană are 23 de perechi (46) de cromozomi, aproximativ 100.000 de gene.Genele sunt localizate pe cromozomi. Multe gene sunt localizate pe un cromozom. Un cromozom cu toate genele din el formează un grup de legătură. Numărul de grupuri de legătură este egal cu setul haploid de cromozomi. O persoană are 23 de grupuri de legătură. Genele situate pe același cromozom nu sunt absolut legate. În timpul meiozei, când cromozomii sunt conjugați, cromozomii omologi schimbă părți. Acest fenomen se numește crossing over, care poate apărea oriunde pe cromozom. Cu cât locii sunt localizați mai departe pe același cromozom unul de celălalt, cu atât mai des poate avea loc un schimb de situsuri între ei (Fig. 76).
La musca Drosophila, genele pentru lungimea aripilor (V - lungă și v - scurtă) și culoarea corpului (B - gri și b - negru) sunt în aceeași pereche de cromozomi omologi, adică. aparțin aceluiași grup de legătură. Dacă încrucișați o muscă cu culoarea corpului gri și aripi lungi cu o muscă neagră cu aripi scurte, atunci în prima generație toate muștele vor avea culoarea corpului gri și aripi lungi (Fig. 77).
Ca urmare a încrucișării unui mascul diheterozigot cu o femelă homozigotă recesivă, muștele vor arăta ca părinții lor. Acest lucru se datorează faptului că genele de pe același cromozom sunt moștenite într-un mod legat. La musca mascul Drosophila, ambreiajul este complet. Dacă încrucișați o femelă diheterozigotă cu un mascul homozigot recesiv, atunci unele dintre muște vor arăta ca părinții lor, iar în
Orez. 76. Trecere peste.
1 - doi cromozomi omologi; 2 - al lor decusație în timpul conjugării; 3 - două noi combinații de cromozomi.
pe de altă parte, va avea loc o recombinare a caracteristicilor. O astfel de moștenire are loc pentru genele aceluiași grup de legătură, între care poate avea loc încrucișarea. Acesta este un exemplu de legare incompletă a genelor.
Principalele prevederi ale teoriei cromozomiale a eredității
. Genele sunt localizate pe cromozomi.
. Genele sunt aranjate liniar pe un cromozom.

Orez. 77.Moștenirea legată a genelor pentru culoarea corpului și starea aripilor la musca de fructe.
Gena gri (B) domină gena de culoare a corpului negru (b), gena aripilor lungi (V) domină gena aripilor scurte (v). B și V sunt pe același cromozom.
a - legarea completă a genelor datorită absenței încrucișării cromozomilor la masculii Drosophila: PP - o femelă cenușie cu aripi lungi (BBVV) este încrucișată cu un mascul negru cu aripi scurte (bbvv); F 1 - mascul cenușiu cu aripi lungi (BbVv) încrucișat cu o femelă neagră cu aripi scurte (bbvv); F 2 - deoarece masculul nu trece peste, vor apărea două tipuri de descendenți: 50% - negru cu aripi scurte și 50% - gri cu aripi normale; b - legarea incompletă (parțială) a trăsăturilor din cauza încrucișării cromozomilor la femelele Drosophila: PP - o femelă cu aripi lungi (BBVV) este încrucișată cu un mascul negru cu aripi scurte (bbvv); F 1 - o femelă cenușie cu aripi lungi (BbVv) este încrucișată cu un mascul negru cu aripi scurte (bbvv). F 2 - deoarece încrucișarea cromozomilor omologi are loc la femelă, se formează patru tipuri de gameți și vor apărea patru tipuri de descendenți: neîncrucișați - gri cu aripi lungi (BbVv) și negru cu aripi scurte (bbvv), încrucișări - negru cu aripi lungi (bbVv), gri cu aripi scurte (Bbvv).
. Fiecare genă ocupă un loc specific - un loc.
. Fiecare cromozom este un grup de legătură. Numărul de grupuri de legătură este egal cu numărul haploid de cromozomi.
Genele alelice sunt schimbate între cromozomi omologi. Distanța dintre gene este proporțională cu procentul de încrucișare între ele.
Întrebări pentru autocontrol
1. Unde sunt localizate genele?
2. Ce este un grup de ambreiaj?
3. Care este numărul de grupuri de legătură?
4. Cum sunt legate genele în cromozomi?
5. Cum se moștenește trăsătura lungimii aripilor și a culorii corpului la muștele Drosophila?
6. Ce urmași va apărea la încrucișarea unei femele homozigote cu aripi lungi și în gri corpuri cu un mascul negru homozigot cu aripi scurte?
7. Ce urmași va apărea când un mascul diheterozigot este încrucișat cu o femelă homozigotă recesivă?
8. Ce fel de legătură genică are masculul Drosophila?
9. Care va fi urmașul când o femelă diheterozigotă este încrucișată cu un mascul homozigot recesiv?
10. Ce fel de legătură genică are femela Drosophila?
11. Care sunt principalele prevederi ale teoriei cromozomiale a eredității?
Cuvinte cheie subiect « Teoria cromozomiala ereditate"
genele
grup de ambreiaj
lungime
celule
conjugare
trecere peste
aripile
zbură locală liniară
ereditate
schimb valutar
colorare
organism de cuplu
recombinare
generaţie
poziţie
urmasi
distanţă
rezultat
părinţi
masculin
Femeie
trecere
corp
teorie
complot
cromozomii
culoare
Parte
Uman
număr
Mecanismul de determinare a sexului cromozomal
Diferențele fenotipice între indivizi de diferite sexe se datorează genotipului. Genele sunt localizate pe cromozomi. Există reguli de individualitate, constanță, împerechere a cromozomilor. Setul diploid de cromozomi se numește cariotip. Există 23 de perechi (46) de cromozomi în cariotipul feminin și masculin (Fig. 78).
22 de perechi de cromozomi sunt la fel. Ei sunt numiti, cunoscuti autozomi. A 23-a pereche de cromozomi - cromozomi sexuali.În cariotipul feminin, unul

Orez. 78.Cariotipuri ale diferitelor organisme.1 - o persoana; 2 - tantari; 3 plante skerda.
cromozomii sexuali XX. În cariotipul masculin, cromozomii sexuali sunt XY. Cromozomul Y este foarte mic și conține puține gene. Combinația de cromozomi sexuali din zigot determină sexul viitorului organism.
În timpul maturării celulelor germinale, ca urmare a meiozei, gameții primesc un set haploid de cromozomi. Fiecare ou conține 22 de autozomi + un cromozom X. Sexul care produce gameți care sunt aceiași pe cromozomul sexual se numește sex homogametic. Jumătate dintre spermatozoizi conțin - 22 autozomi + cromozom X și jumătate 22 autozomi + Y. Sexul care formează gameți care sunt diferiți pe cromozomul sexual se numește heterogametic. Sexul copilului nenăscut este determinat în momentul fertilizării. Dacă un ovul este fertilizat de un spermatozoid al cromozomului X, acesta se dezvoltă corp feminin dacă cromozomul Y este masculin (Fig. 79).

Orez. 79.Mecanismul cromozomal al formării sexului.
Probabilitatea de a avea un băiat sau o fată este de 1:1 sau 50%:50%. Această definiție a sexului este tipică pentru oameni și mamifere. Unele insecte (lăcuste și gândaci) nu au un cromozom Y. Masculii au un cromozom X (X0), iar femelele doi (XX). La albine, femelele au 2n seturi de cromozomi (32 de cromozomi), în timp ce masculii au n (16 cromozomi). Femeile au doi cromozomi X sexuali în celulele lor somatice. Unul dintre ele formează un bulgăre de cromatină, care poate fi văzut în nucleele de interfază atunci când este tratat cu un reactiv. Acest bulgăre este corpul lui Barr. Masculii nu au un corp Barr, deoarece au doar un cromozom X. Dacă în timpul meiozei doi cromozomi XX intră în ovul deodată și un astfel de ovul este fertilizat de un spermatozoid, atunci zigotul va avea un număr mai mare de cromozomi.

De exemplu, un organism cu un set de cromozomi XXX (trisomie pe cromozomul X) fenotipul este o fată. Are gonade subdezvoltate. Există doi corpi Barr în nucleii celulelor somatice.
Un organism cu un set de cromozomi XXY (sindromul Klinefelter) fenotipul este un băiat. Testiculele lui sunt subdezvoltate, se remarcă retard fizic și mental. Există un cadavru Barr.
Cromozomii XO (monozomia pe cromozomul X)- a determina Sindromul Shereshevsky-Turner. Un organism cu un astfel de set este o fată. Are gonade subdezvoltate, statură mică. Nici un corp Barr. Un organism care nu are un cromozom X, dar conține doar un cromozom Y, nu este viabil.
Moștenirea trăsăturilor ale căror gene sunt localizate pe cromozomii X sau Y se numește moștenire legată de sex. Dacă genele se află pe cromozomii sexuali, ele sunt moștenite legate de sex.
O persoană are o genă pe cromozomul X care determină semnul coagulării sângelui. Gena recesivă provoacă dezvoltarea hemofiliei. Cromozomul X are o genă (recesivă) care este responsabilă de manifestarea daltonismului. Femeile au doi cromozomi X. O trăsătură recesivă (hemofilie, daltonism) apare numai dacă genele responsabile de aceasta sunt localizate pe doi cromozomi X: X h X h; X d X d . Dacă un cromozom X are o genă H sau D dominantă, iar celălalt are o genă h sau d recesivă, atunci nu va exista hemofilie sau daltonism. Barbatii au un cromozom X. Dacă are o genă H sau h, atunci aceste gene își vor arăta cu siguranță efectul, deoarece cromozomul Y nu poartă aceste gene.
O femeie poate fi homozigotă sau heterozigotă pentru genele situate pe cromozomul X, dar genele recesive apar doar în starea homozigotă.

Dacă genele se află pe cromozomul Y (moștenirea olandeză), apoi semnele condiţionate de ei se transmit din tată în fiu. De exemplu, părul urechii este moștenit prin cromozomul Y. Barbatii au un cromozom X. Toate genele din el, inclusiv cele recesive, apar în fenotip. La sexul heterogametic (masculin), majoritatea genelor situate pe cromozomul X sunt localizate pe hemizigot stare, adică nu au o pereche alelică.
Cromozomul Y conține unele gene care sunt omoloage cu genele cromozomului X, de exemplu, gene pentru diateza hemoragică, daltonismul general etc. Aceste gene sunt moștenite atât prin cromozomul X, cât și prin cromozomul Y.
Întrebări pentru autocontrol
1. Care sunt regulile cromozomilor?
2. Ce este un cariotip?
3. Câți autozomi are o persoană?
4. Ce cromozomi umani sunt responsabili de dezvoltarea sexului?
5. Care este probabilitatea de a avea un băiat sau o fată?
6. Cum este determinat sexul la lăcuste și gândaci?
7. Cum se determină sexul la albine?
8. Cum se determină sexul la fluturi și păsări?
9. Ce este un corp Barr?
10. Cum poți determina prezența unui corp Barr?
11. Ce poate explica apariția mai multor sau mai puțini cromozomi în cariotip?
12. Ce este moștenirea legată de sex?
13. Ce gene sunt legate de sex la oameni?
14. Cum și de ce genele recesive legate de sex își arată efectul la femei?
15. Cum și de ce genele recesive legate de cromozomul X la bărbați își arată efectul?
Cuvinte cheie ale subiectului „Determinarea cromozomală a sexului”
autozomi
fluturi
probabilitate
părul urechii
gameti
genotip
genele
sex heterogametic
bulgăre de cromatină
sex homogametic
daltonism
fată
acțiune
femeie
zigot
individualitate
cariotip
lăcuste
băiat
meioză
mamifer
moment
monosomie
om
trusa
insecte
moştenire
purtător
tratament cu reactiv de fertilizare
organism
individual
împerechere
cupluri
podea
celulele sexuale
descendenți
reguli
semn
păsări
albinele
dezvoltare
diferențe
naștere
înălţime
sindromul Down de coagulare a testiculelor
Sindromul Klinefelter
Sindromul Shershevsky-Turner
orbire
maturare
stat
combinaţie
spermatozoizi
fiul
gandaci de bucatarie
Corpul lui Barr
trisomie
cromozomul Y
fenotip
cromozom
cromozomul X
Uman
miez
ou
§ 5. T. G. Morgan și teoria sa cromozomială
Thomas Gent Morgan s-a născut în 1866 în Kentucky (SUA). După ce a absolvit universitatea la douăzeci de ani, Morgan a primit titlul de doctor în științe la douăzeci și patru de ani, iar la douăzeci și cinci de ani a devenit profesor.
Din 1890, Morgan s-a angajat în embriologia experimentală. În primul deceniu al secolului al XX-lea, îi plăcea întrebările legate de ereditate.

Sună paradoxal, dar la începutul activității sale, Morgan a fost un oponent înflăcărat al învățăturilor lui Mendel și urma să-și respingă legile privind obiectele animale - iepurii. Cu toate acestea, administratorii Universității Columbia au considerat că experiența este prea costisitoare. Așa că Morgan și-a început cercetările asupra unui obiect mai ieftin - musca de fructe Drosophila, iar apoi nu numai că nu a ajuns la negarea legilor lui Mendel, ci a devenit și un demn succesor al învățăturilor sale.

Un cercetător în experimente cu Drosophila creează teoria cromozomiala a ereditatii- cea mai mare descoperire, ocupând, prin expresie N. K. Koltsova, „același loc în biologie ca teoria moleculară în chimie și teoria structurilor atomice în fizică”.
În 1909-1911. Morgan și studenții săi la fel de iluștri A. Sturtevant, G. Moeller, C. Bridges a arătat că a treia lege a lui Mendel necesită completări semnificative: înclinațiile ereditare nu sunt întotdeauna moștenite independent; uneori se transmit în grupuri întregi – legate între ele. Astfel de grupuri situate pe cromozomul corespunzător se pot muta la un alt cromozom omolog în timpul conjugării cromozomilor în timpul meiozei (profaza I).
A fost formulată teoria cromozomală completă T. G. Morganîn perioada 1911-1926. Cu apariţia sa şi dezvoltare ulterioară această teorie este îndatorată nu numai lui Morgan și școlii sale, ci și lucrării unui număr semnificativ de oameni de știință, atât străini, cât și autohtoni, dintre care, în primul rând, trebuie să menționăm N. K. KoltsovaȘi A. S. Serebrovsky (1872-1940).
Conform teoriei cromozomilor, transmiterea informaţiei ereditare este asociată cu cromozomii, in care liniar, la un anumit loc (din lat. locus- loc), genele mint. Deoarece cromozomii sunt perechi, fiecare genă de pe un cromozom corespunde unei gene pereche de pe celălalt cromozom (omologul) aflată în același locus. Aceste gene pot fi aceleași (la homozigoți) sau diferite (la heterozigoți). Diferite forme se numesc genele care apar prin mutație din original alele, sau alelomorfe(din greaca allo - diferit, morph - forma). Alelele afectează manifestarea unei trăsături în moduri diferite. Dacă o genă există în mai mult de două stări alele, atunci astfel de alele în populatiilor* formează o serie de așa-numite alele multiple. Fiecare individ dintr-o populație poate conține oricare două (dar nu mai multe) alele în genotipul său, iar fiecare gamet poate conține doar o alelă, respectiv. În același timp, indivizii cu orice alele din această serie pot fi în populație. Alelele hemoglobinei sunt un exemplu de alele multiple (vezi Capitolul I, § 5).
* (O populație (din latină popularus - populație) este un grup de indivizi din aceeași specie, uniți prin încrucișări reciproce, izolat într-o oarecare măsură de alte grupuri de indivizi din această specie.)
Gradul de dominanță într-o serie de alele poate crește de la gena extremă recesivă la cea extremă dominantă. poate conduce număr mare exemple de acest tip. Deci, la iepuri, seria de gene recesive alele multiple este gena c care determină dezvoltarea albinismului*. Gena c h de colorație himalayană (hermină) (ochi roz, corp alb, vârfuri închise ale nasului, urechilor, coadă și membrelor) va fi dominantă în raport cu această genă; peste această genă, precum și peste gena c, domină gena de culoare gri deschis (chinchilla) c ch. Un stadiu și mai dominant este gena agouti - c a (domină asupra genelor c, c h și c ch). Cea mai dominantă din întreaga serie, gena de culoare neagră C domină peste toate „treptele inferioare ale alelelor” - genele c, c h, c ch, c a.
* (Lipsa pigmentului (vezi capitolul VII, § 5).)
Dominanța, ca și recesivitatea alelelor, nu este o proprietate absolută, ci relativă. Gradul de dominanță și recesivitate poate fi diferit. Aceeași trăsătură poate fi moștenită într-o manieră dominantă sau recesivă.
Deci, de exemplu, pliul de deasupra colțului interior al ochiului (epicanthus) este moștenit predominant la mongoloizi și recesiv la negroizi (boșmani, hotentoți).
De regulă, alelele nou apărute sunt recesive, dimpotrivă, alelele soiurilor vechi de plante sau rase de animale (și mai multe specii sălbatice) sunt dominante.
Fiecare pereche de cromozomi este caracterizată de un anumit set de gene care alcătuiesc grupul de legătură. De aceea grupurile semne diferite uneori moşteniţi unul cu altul.
Deoarece celulele somatice ale Drosophila conțin patru perechi de cromozomi (2n = 8), iar celulele sexuale conțin jumătate din câte (1n = 4), musca de fructe are patru grupuri ambreiaj; în mod similar, la om, numărul de grupuri de legătură este egal cu numărul de cromozomi din setul haploid (23).
Pentru un număr de organisme (Drosophila, porumb) și unii cromozomi umani*, au fost compilate hărți cromozomiale sau genetice, care reprezintă o aranjare schematică a genelor în cromozomi.
* (Până în prezent, a fost posibil să se stabilească localizarea exactă a genelor umane (dacă luăm în considerare numărul total de gene) doar în câteva și relativ cazuri rare, de exemplu, pentru trăsăturile legate de cromozomii sexuali.)

Ca exemplu, să oferim o hartă cromozomală a unei părți a cromozomului X Drosophila (Fig. 24). Cu o precizie mai mare sau mai mică, această hartă reflectă secvența genelor și distanța dintre ele. A fost posibilă determinarea distanței dintre gene folosind analize genetice și citologice de crossing over, care apare în timpul conjugării cromozomilor omologi în timpul zigonemului profazei I a meiozei (vezi Capitolul II, § 7).

Mișcarea genelor de la un cromozom la altul are loc cu o anumită frecvență, care este invers proporțională cu distanța dintre gene: cu cât distanța este mai mică, cu atât mai mare procentul de încrucișare(unitatea de distanță dintre gene este numită după Morgan morganidași este egală cu distanța minimă în cromozom care poate fi măsurată prin încrucișare). Crossover-ul este prezentat în Fig. 25.
În prezent, legătura strânsă a unor loci genici este cunoscută, iar procentul de încrucișare a fost calculat pentru aceștia. Genele legate determină, de exemplu, expresia Factorul Rhși genele sistemului MN al sângelui (despre moștenirea proprietăților sângelui, vezi Capitolul VII, § 3). În unele familii, a fost posibilă urmărirea legăturii factorului Rh cu ovalocitoză(prezența a aproximativ 80-90% eritrocite de formă ovală - anomalia se desfășoară, de regulă, fără manifestări clinice), care dau aproximativ 3% din crossover. Se observă până la 9% de încrucișare între genele care controlează manifestările grupelor sanguine ABO și factorul Lu. Se știe că gena care afectează anomalia structurii unghiilor și a genunchiului este legată și de locii sistemului ABO; procentul de încrucișare între ele este de aproximativ 10. Grupurile de legătură (și, în consecință, hărțile cromozomiale) ale cromozomilor X și Y umani sunt mult mai bine studiate (vezi Capitolul VII, § 6). Se știe, de exemplu, că genele care determină dezvoltarea daltonism(daltonism) și hemofilie(sângerare); procentul de suprapunere dintre ele este de 10.
Corectitudinea ipotezei lui Morgan a fost confirmată la începutul secolului de Kurt Stern (studii citologice) și colaboratorii lui Morgan Theophilus Painter (citolog) și Calvin Bridges (genetician) pe cromozomii giganți ai glandelor salivare ale larvelor de Drosophila (asemănător cu gigantul). cromozomii altor diptere). Pe fig. 26 prezintă o parte a cromozomului gigant al glandei salivare a larvei chironomusului (vierme de sânge).

Când studiați cromozomii giganți cu un microscop cu lumină convențională, striația transversală este clar vizibilă, formată prin alternarea dungilor luminoase și mai întunecate ale discurilor - cromomeri; sunt formate din zone foarte spiralate, dens adiacente.
Formarea unor astfel de cromozomi giganți se numește politenie, adică reduplicarea cromozomilor fără creșterea numărului acestora. În același timp, cromatidele reduplicate rămân una lângă alta, strâns alăturate.
Dacă un cromozom format dintr-o pereche de cromatide se dublează consecutiv de nouă ori, atunci numărul de catene (cromoneme) într-un astfel de cromozom politen va fi de 1024. Datorită despiralizării parțiale a cromozomului, lungimea unui astfel de cromozom crește în comparație cu cea obișnuită. unul de 150-200 de ori.
În 1925, Sturtevant a arătat prezența crossover inegal:într-unul dintre cromozomii omologi pot exista doi loci identici, în care, de exemplu, se află gene care afectează forma ochiului Drosophila - Bar, iar în celălalt - nici un loc. Așa zboară cu un semn pronunțat de ochi cu dungi înguste (gena ultra bar)(vezi fig. 31).
Pe lângă dovezile citologice ale corectitudinii teoriei cromozomilor, au fost efectuate experimente genetice - încrucișând diferite rase de Drosophila. Deci, printre multele gene legate în mușca fructelor, există două gene recesive: gena pentru culoarea corpului negru ( negru) și gena pentru aripi rudimentare ( vestigială).
Să le numim gene a și b. Ele corespund la două alele dominante: gena pentru corpul gri și aripile dezvoltate în mod normal (A și B). La încrucișarea muștelor de rasă aabb și AABB, întreaga primă generație de hibrizi va avea genotipul AaBb. Teoretic vorbind, următoarele rezultate ar trebui să fie așteptate în a doua generație (F 2).

Cu toate acestea, într-un procent mic, dar constant de cazuri, au fost întâlniți descendenți neobișnuiți din gameți neobișnuiți. Aproximativ 18% dintre astfel de gameți au fost observați în fiecare încrucișare (9% Ab și 9% aB).

Apariția unor astfel de excepții este bine explicată prin procesul de încrucișare. Astfel, studiile genetice au făcut posibilă și stabilirea faptului că tulburarea ambreiajului este încrucișarea, care duce la o creștere a variabilității formei, este constantă statistic.
În concluzie, observăm că o serie de prevederi ale geneticii clasice au suferit o serie de modificări astăzi.
Am folosit în mod repetat termenii de gene (alele) și trăsături „dominante” și „recesive”. Cu toate acestea, studii recente au arătat că așa-numitele gene recesive pot de fapt să nu fie deloc recesive. Este mai corect să spunem că genele recesive dau o manifestare vizibilă sau invizibilă foarte slabă în fenotip. Dar în acest din urmă caz, alelele recesive, invizibile în exterior în fenotip, pot fi detectate folosind tehnici biochimice speciale. În plus, aceeași genă în anumite condiții de mediu se poate comporta ca dominantă, în altele - ca recesiv.
Întrucât dezvoltarea tuturor organismelor are loc în funcție de și sub influența mediului extern, manifestarea genotipului într-un anumit fenotip este influențată și de factorii de mediu (temperatura, hrana, umiditatea și compoziția gazelor din atmosferă, presiunea acestuia, prezența formelor patogene pentru un organism dat, compoziția chimică a apei, a solului etc., dar pentru om și fenomene ordine socială). Fenotipul nu arată niciodată toate posibilitățile genotipice. Prin urmare, în conditii diferite manifestările fenotipice ale genotipurilor strâns înrudite pot fi foarte diferite unele de altele. Astfel, atât genotipul cât și mediul sunt implicate (într-o măsură mai mare sau mai mică) în manifestarea unei trăsături.