Codul genetic este citit continuu. Cod genetic: descriere, caracteristici, istoric de cercetare
Citeste si
Funcțiile genetice ale ADN-ului constă în faptul că asigură stocarea, transmiterea și implementarea informațiilor ereditare, care sunt informații despre structura primară a proteinelor (adică, compoziția lor de aminoacizi). Relația ADN-ului cu sinteza proteinelor a fost prezisă de biochimiștii J. Beadle și E. Tatum încă din 1944, în timp ce studiau mecanismul mutațiilor în ciuperca de mucegai Neurospora. Informațiile sunt înregistrate ca o secvență specifică de baze azotate într-o moleculă de ADN folosind codul genetic. Descifrarea codului genetic este considerată una dintre marile descoperiri ale științelor naturale din secolul XX. și sunt echivalate ca importanță cu descoperirea energiei nucleare în fizică. Succesul în acest domeniu este asociat cu numele omului de știință american M. Nirenberg, în laboratorul căruia a fost descifrat primul codon, YYY. Cu toate acestea, întregul proces de descifrare a durat mai mult de 10 ani, mulți oameni de știință celebri de la tari diferite, și nu numai biologi, ci și fizicieni, matematicieni, cibernetici. O contribuție decisivă la dezvoltarea mecanismului de înregistrare a informațiilor genetice a avut-o G. Gamow, care a fost primul care a sugerat că un codon este format din trei nucleotide. Eforturile comune ale oamenilor de știință au dat caracteristică completă cod genetic.
Literele din cercul interior sunt bazele în prima poziție în codon, literele din al doilea cerc sunt
bazele în poziţia a 2-a şi literele din afara celui de-al doilea cerc sunt bazele în poziţia a 3-a.
În ultimul cerc - nume prescurtate ale aminoacizilor. NP - nepolar,
P - reziduuri polare de aminoacizi.
Principalele proprietăți ale codului genetic sunt: tripletate, degenerareși nesuprapunere. Tripletatea înseamnă că secvența a trei baze determină includerea unui aminoacid specific în molecula proteică (de exemplu, AUG - metionină). Degenerarea codului este că același aminoacid poate fi codificat de doi sau mai mulți codoni. Nesuprapunerea înseamnă că aceeași bază nu poate fi prezentă în doi codoni adiacenți.
S-a constatat că codul este universal, adică Principiul înregistrării informațiilor genetice este același în toate organismele.
Tripleții care codifică același aminoacid se numesc codoni sinonimi. De obicei au aceleași baze în pozițiile 1 și 2 și diferă doar în baza a treia. De exemplu, includerea aminoacidului alanină într-o moleculă de proteină este codificată de codoni sinonimi în molecula de ARN - GCA, GCC, GCG, GCY. Codul genetic conține trei tripleți necodificatori (codoni nonsens - UAG, UGA, UAA), care joacă rolul de semnale de stop în procesul de citire a informațiilor.
S-a stabilit că universalitatea codului genetic nu este absolută. În timp ce se menține principiul de codificare comun tuturor organismelor și caracteristicile codului, în unele cazuri se observă o schimbare a încărcăturii semantice a cuvintelor cod individuale. Acest fenomen a fost numit ambiguitatea codului genetic, iar codul în sine a fost numit cvasi-universal.
Citeste si alte articole subiectele 6 „Bazele moleculare ale eredității”:
Mergeți la citirea altor subiecte ale cărții "Genetica și selecția. Teorie. Sarcini. Răspunsuri".
După ce ați lucrat la aceste subiecte, ar trebui să puteți:
- Descrieți următoarele concepte și explicați relația dintre ele:
- polimer, monomer;
- carbohidrat, monozaharid, dizaharid, polizaharid;
- lipide, acizi grași, glicerol;
- aminoacid, legătură peptidică, proteină;
- catalizator, enzimă, situs activ;
- acid nucleic, nucleotidă.
- Enumerați 5-6 motive pentru care apa este o componentă atât de importantă a sistemelor vii.
- Numiți cele patru clase principale compusi organici conținute în organismele vii; descrie rolul fiecăruia.
- Explicați de ce reacțiile controlate de enzime depind de temperatură, pH și prezența coenzimelor.
- Descrieți rolul ATP-ului în economia energetică a celulei.
- Numiți materiile prime, etapele principale și produsele finale ale reacțiilor induse de lumină și ale reacțiilor de fixare a carbonului.
- A da scurta descriere schema generala respirația celulară, din care ar fi clar ce loc ocupă reacțiile de glicoliză, ciclul G. Krebs (ciclul acid citric) și lanțul de transport de electroni.
- Comparați respirația și fermentația.
- Descrieți structura moleculei de ADN și explicați de ce numărul de resturi de adenină este egal cu numărul de resturi de timină, iar numărul de resturi de guanină este egal cu numărul de resturi de citozină.
- Realizați o scurtă schemă pentru sinteza ARN-ului în ADN (transcripție) la procariote.
- Descrieți proprietățile codului genetic și explicați de ce ar trebui să fie triplet.
- Pe baza acestui lanț de ADN și a tabelului de codoni, determinați secvența complementară a ARN-ului matricei, indicați codonii ARN-ului de transfer și secvența de aminoacizi care se formează ca rezultat al translației.
- Enumeră pașii sinteza proteinei la nivelul ribozomului.
Algoritm pentru rezolvarea problemelor.
Tip 1. Autocopie ADN.
Unul dintre lanțurile de ADN are următoarea secvență de nucleotide:
AGTACCGATACCGATTTCG...
Ce secvență de nucleotide are al doilea lanț al aceleiași molecule?
Pentru a scrie secvența de nucleotide a celei de-a doua catene a unei molecule de ADN, când este cunoscută secvența primei catene, este suficient să înlocuiți timina cu adenină, adenina cu timină, guanina cu citozină și citozina cu guanină. Făcând această înlocuire, obținem succesiunea:
TACTGGCTATGAGCTAAATG...
Tip 2. Codificarea proteinelor.
Lanțul de aminoacizi al proteinei ribonuclează are următorul început: lizină-glutamină-treonină-alanină-alanină-alanină-lizină...
Ce secvență de nucleotide pornește gena corespunzătoare acestei proteine?
Pentru a face acest lucru, utilizați tabelul codului genetic. Pentru fiecare aminoacid, găsim codul său sub forma trio-ului corespunzător de nucleotide și îl scriem. Dispunând aceste triplete unul după altul în aceeași ordine cu aminoacizii corespunzători, obținem formula pentru structura secțiunii de ARN mesager. De regulă, există mai multe astfel de triple, alegerea se face în funcție de decizia dvs. (dar se ia doar unul dintre triple). Pot exista mai multe soluții, respectiv.
AAACAAAATSUGTSGGTSUGTSGAAG
Cu ce secvență de aminoacizi începe o proteină dacă este codificată de o astfel de secvență de nucleotide:
ACGCCATGCCGGGT...
Conform principiului complementarității, găsim structura secțiunii informaționale de ARN formată pe un anumit segment al moleculei de ADN:
UGCGGGUACCCGCCCCA...
Apoi ne întoarcem la tabelul codului genetic și pentru fiecare trio de nucleotide, pornind de la prima, găsim și scriem aminoacidul corespunzător acestuia:
Cisteina-glicina-tirozina-arginina-prolina-...
Ivanova T.V., Kalinova G.S., Myagkova A.N. " Biologie generală". Moscova, "Iluminismul", 2000
- Subiectul 4." Compoziție chimică celule.” §2-§7 p. 7-21
- Subiectul 5. „Fotosinteza”. §16-17 p. 44-48
- Subiectul 6. „Respirația celulară”. §12-13 p. 34-38
- Subiectul 7. „Informații genetice”. §14-15 p. 39-44
COD GENETIC, sistem de înregistrare a informațiilor ereditare sub forma unei secvențe de baze nucleotidice din moleculele de ADN (la unele virusuri - ARN), care determină structura primară (aranjarea resturilor de aminoacizi) în moleculele proteice (polipeptide). Problema codului genetic a fost formulată după demonstrarea rolului genetic al ADN-ului (microbiologii americani O. Avery, K. McLeod, M. McCarthy, 1944) și descifrarea structurii acestuia (J. Watson, F. Crick, 1953), după stabilirea că genele determină structura și funcțiile enzimelor (principiul „o genă – o enzimă” de J. Beadle și E. Tatema, 1941) și că există o dependență a structurii și activității spațiale a unei proteine de structura sa primară (F. Senger, 1955). Întrebarea modului în care combinațiile a 4 baze de acizi nucleici determină alternanța a 20 de reziduuri comune de aminoacizi în polipeptide a fost ridicată pentru prima dată de G. Gamow în 1954.
Pe baza unui experiment în care au fost studiate interacțiunile inserțiilor și delețiilor unei perechi de nucleotide, într-una dintre genele bacteriofagului T4, F. Crick și alți oameni de știință au determinat în 1961 proprietățile generale ale codului genetic: triplet, i.e. , fiecare rest de aminoacid din lanţul polipeptidic corespunde unui set de trei baze (triplet sau codon) din ADN-ul unei gene; citirea codonilor în cadrul unei gene merge dintr-un punct fix, într-o direcție și „fără virgule”, adică codonii nu sunt separați prin semne unul de celălalt; degenerare, sau redundanță, - același rest de aminoacizi poate codifica mai mulți codoni (codoni sinonimi). Autorii au sugerat că codonii nu se suprapun (fiecare bază aparține unui singur codon). Studiul direct al capacității de codare a tripleților a fost continuat folosind un sistem de sinteză a proteinelor fără celule sub controlul ARN mesager sintetic (ARNm). Până în 1965, codul genetic a fost complet descifrat în lucrările lui S. Ochoa, M. Nirenberg și H. G. Korana. Dezvăluirea misterului codului genetic a fost una dintre realizările remarcabile ale biologiei în secolul al XX-lea.
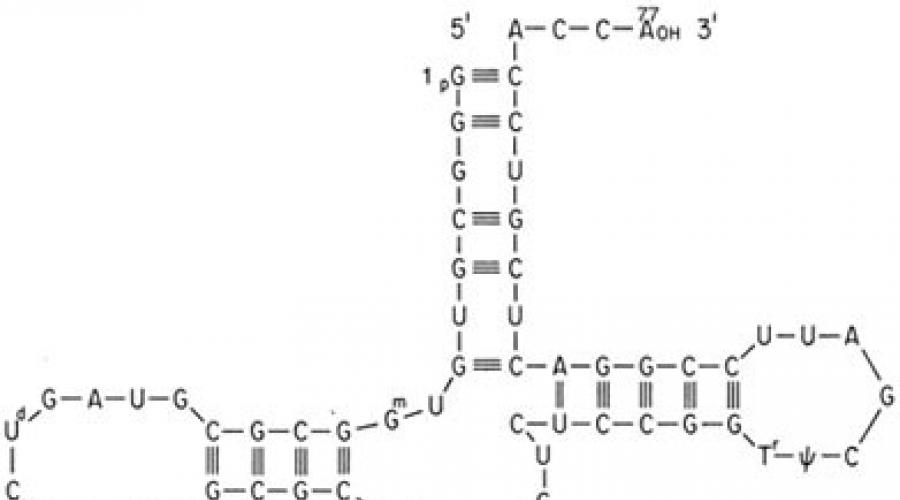
Implementarea codului genetic în celulă are loc în cursul a două procese matrice - transcripție și traducere. Mediatorul dintre o genă și o proteină este ARNm, care se formează în timpul transcripției pe una dintre catenele de ADN. În acest caz, secvența de baze ADN, care poartă informații despre structura primară a proteinei, este „rescrisă” sub forma unei secvențe de baze de ARNm. Apoi, în timpul translației pe ribozomi, secvența de nucleotide a ARNm este citită de ARN de transfer (ARNt). Acestea din urmă au un capăt acceptor, la care este atașat un rest de aminoacid și un capăt adaptor, sau triplet anticodon, care recunoaște codonul ARNm corespunzător. Interacțiunea codonului și anti-codonului are loc pe baza împerecherii de baze complementare: Adenină (A) - Uracil (U), Guanină (G) - Citozină (C); în acest caz, secvența de baze ARNm este tradusă în secvența de aminoacizi a proteinei sintetizate. Diverse organisme utilizați codoni-sinonime diferiți pentru același aminoacid cu frecvențe diferite. Citirea ARNm care codifică lanțul polipeptidic începe (inițiază) de la codonul AUG corespunzător aminoacidului metionină. Mai rar la procariote, codonii de inițiere sunt GUG (valină), UUG (leucină), AUU (izoleucină), la eucariote - UUG (leucină), AUA (izoleucină), ACG (treonină), CUG (leucină). Aceasta stabilește așa-numitul cadru, sau fază, de citire în timpul translației, adică atunci întreaga secvență de nucleotide a ARNm este citită triplet cu triplet de ARNt până când oricare dintre cei trei codoni terminatori, adesea numiți codoni stop, este găsit pe ARNm: UAA, UAG, UGA (tabel). Citirea acestor tripleți duce la finalizarea sintezei lanțului polipeptidic.
Codonii AUG și stop sunt localizați la începutul și, respectiv, la sfârșitul regiunilor ARNm care codifică polipeptide.
Cod geneticși cvasi-universală. Aceasta înseamnă că există mici variații în sensul unor codoni în diferite obiecte, iar aceasta se referă, în primul rând, la codonii terminatori, care pot fi semnificativi; de exemplu, în mitocondriile unor eucariote și în micoplasme, codurile UGA pentru triptofan. În plus, în unele ARNm de bacterii și eucariote, UGA codifică un aminoacid neobișnuit, selenocisteina, iar UAG, într-una dintre arhebacterii, codifică pirolizina.
Există un punct de vedere conform căruia codul genetic a apărut întâmplător (ipoteza „cazului înghețat”). Este mai probabil să fi evoluat. Această presupunere este susținută de existența unei versiuni mai simple și, aparent, mai veche a codului, care este citită în mitocondrii conform regulii „două din trei”, când doar două din cele trei baze din triplet determină amino. acid.
Lit.: Crick F. N. a. despre. Natura generală a codului genetic pentru proteine // Nature. 1961 Vol. 192; Codul genetic. N.Y., 1966; Ichas M. Cod biologic. M., 1971; Inge-Vechtomov S. G. Cum este citit codul genetic: reguli și excepții // Știința naturală modernă. M., 2000. T. 8; Ratner V. A. Codul genetic ca sistem // Soros Educational Journal. 2000. V. 6. Nr. 3.
S. G. Inge-Vechtomov.
Codul genetic este un sistem de înregistrare a informațiilor ereditare în moleculele de acid nucleic, bazat pe o anumită alternanță de secvențe de nucleotide din ADN sau ARN care formează codoni corespunzători aminoacizilor dintr-o proteină.
Proprietățile codului genetic.
Codul genetic are mai multe proprietăți.
Tripletate.
Degenerare sau redundanță.
Neambiguitate.
Polaritate.
Nesuprapunere.
Compactitate.
Versatilitate.
Trebuie remarcat faptul că unii autori oferă și alte proprietăți ale codului legate de caracteristici chimice nucleotidele incluse în cod sau cu frecvența de apariție a aminoacizilor individuali în proteinele corpului etc. Cu toate acestea, aceste proprietăți decurg din cele de mai sus, așa că le vom lua în considerare acolo.
A. Tripletate. Codul genetic, ca multe sisteme organizate complex, are cea mai mică unitate structurală și cea mai mică unitate funcțională. Un triplet este cea mai mică unitate structurală a codului genetic. Este format din trei nucleotide. Un codon este cea mai mică unitate funcțională a codului genetic. De regulă, tripleții ARNm sunt numiți codoni. În codul genetic, un codon îndeplinește mai multe funcții. În primul rând, funcția sa principală este aceea de a codifica un aminoacid. În al doilea rând, un codon poate să nu codifice pentru un aminoacid, dar în acest caz are o funcție diferită (vezi mai jos). După cum se poate vedea din definiție, un triplet este un concept care caracterizează elementar unitate structurală cod genetic (trei nucleotide). codonul caracterizează unitate semantică elementară genom - trei nucleotide determină atașarea la lanțul polipeptidic a unui aminoacid.
Unitatea structurală elementară a fost mai întâi descifrată teoretic, iar apoi existența ei a fost confirmată experimental. Într-adevăr, 20 de aminoacizi nu pot fi codificați de una sau două nucleotide. acestea din urmă sunt doar 4. Trei din patru nucleotide dau 4 3 = 64 de variante, ceea ce acoperă mai mult decât numărul de aminoacizi prezenți în organismele vii (vezi Tabelul 1).
Combinațiile de nucleotide prezentate în Tabelul 64 au două caracteristici. În primul rând, dintre cele 64 de variante de tripleți, doar 61 sunt codoni și codifică orice aminoacid, se numesc codoni de sens. Trei tripleți nu codifică
aminoacizii a sunt semnale de stop care marchează sfârșitul translației. Există trei astfel de tripleți UAA, UAG, UGA, ele sunt numite și „fără sens” (codoni aiurea). Ca rezultat al unei mutații, care este asociată cu înlocuirea unei nucleotide într-un triplet cu altul, un codon fără sens poate apărea dintr-un codon sens. Acest tip de mutație se numește mutație aiurea. Dacă se formează un astfel de semnal de oprire în interiorul genei (în partea sa informațională), atunci în timpul sintezei proteinelor în acest loc, procesul va fi întrerupt constant - va fi sintetizată doar prima parte (înainte de semnalul de oprire) a proteinei. O persoană cu o astfel de patologie va experimenta o lipsă de proteine și va experimenta simptome asociate cu această lipsă. De exemplu, acest tip de mutație a fost găsit în gena care codifică lanțul beta al hemoglobinei. Se sintetizează un lanț scurt de hemoglobină inactiv, care este distrus rapid. Ca rezultat, se formează o moleculă de hemoglobină lipsită de lanț beta. Este clar că este puțin probabil ca o astfel de moleculă să-și îndeplinească pe deplin sarcinile. Există o boală gravă care se dezvoltă în funcție de tipul de anemie hemolitică (talasemie beta-zero, din cuvântul grecesc „Talas” – Marea Mediterană, unde a fost descoperită prima dată această boală).
Mecanismul de acțiune al codonilor stop este diferit de mecanismul de acțiune al codonilor de sens. Acest lucru rezultă din faptul că pentru toți codonii care codifică aminoacizi, s-au găsit ARNt-urile corespunzătoare. Nu s-au găsit ARNt pentru codonii nonsens. Prin urmare, ARNt nu ia parte la procesul de oprire a sintezei proteinelor.
codonAUG (uneori GUG în bacterii) nu numai că codifică aminoacidul metionină și valină, dar este șiinițiator de difuzare .
b. Degenerare sau redundanță.
61 din cele 64 de tripleți codifică 20 de aminoacizi. Un astfel de exces de trei ori al numărului de tripleți față de numărul de aminoacizi sugerează că două opțiuni de codare pot fi utilizate în transferul de informații. În primul rând, nu toți cei 64 de codoni pot fi implicați în codificarea a 20 de aminoacizi, ci doar 20, iar în al doilea rând, aminoacizii pot fi codificați de mai mulți codoni. Studiile au arătat că natura a folosit această ultimă opțiune.
Preferința lui este clară. Dacă doar 20 din 64 de variante triplete ar fi implicate în codificarea aminoacizilor, atunci 44 de tripleți (din 64) ar rămâne necodificatori, adică. fără sens (codoni aiurea). Mai devreme, am subliniat cât de periculoasă pentru viața celulei este transformarea tripletului de codificare ca urmare a mutației într-un codon fără sens - acest lucru perturbă în mod semnificativ funcționarea normală a ARN polimerazei, ducând în cele din urmă la dezvoltarea bolilor. În prezent există trei codoni nonsens în genomul nostru și acum imaginați-vă ce s-ar întâmpla dacă numărul de codoni nonsens crește de aproximativ 15 ori. Este clar că într-o astfel de situație tranziția codonilor normali la codoni nonsens va fi nemăsurat mai mare.
Un cod în care un aminoacid este codificat de mai multe triplete se numește degenerat sau redundant. Aproape fiecare aminoacid are mai mulți codoni. Deci, aminoacidul leucina poate fi codificat de șase tripleți - UUA, UUG, CUU, CUC, CUA, CUG. Valina este codificată de patru tripleți, fenilalanina de două și numai triptofan și metionină codificat de un codon. Proprietatea care este asociată cu înregistrarea aceleiași informații cu caractere diferite este numită degenerare.
Numărul de codoni alocați unui aminoacid se corelează bine cu frecvența de apariție a aminoacidului în proteine.
Și cel mai probabil acest lucru nu este întâmplător. Cu cât frecvența de apariție a unui aminoacid într-o proteină este mai mare, cu atât codonul acestui aminoacid este mai des reprezentat în genom, cu atât este mai mare probabilitatea de deteriorare a acestuia. factori mutageni. Prin urmare, este clar că un codon mutant are mai multe șanse de a codifica același aminoacid dacă este foarte degenerat. Din aceste poziții, degenerarea codului genetic este un mecanism care protejează genomul uman de deteriorare.
Trebuie remarcat faptul că termenul de degenerescență este folosit în genetica moleculară și într-un alt sens. Deoarece partea principală a informațiilor din codon cade pe primele două nucleotide, baza din a treia poziție a codonului se dovedește a fi de mică importanță. Acest fenomen se numește „degenerarea bazei a treia”. Ultima caracteristică minimizează efectul mutațiilor. De exemplu, se știe că funcția principală a globulelor roșii este de a transporta oxigenul de la plămâni la țesuturi și dioxid de carbon de la țesuturi la plămâni. Această funcție este îndeplinită de pigmentul respirator - hemoglobina, care umple întreaga citoplasmă a eritrocitei. Este format dintr-o parte proteică - globină, care este codificată de gena corespunzătoare. Pe lângă proteine, hemoglobina conține hem, care conține fier. Mutațiile în genele globinei au ca rezultat diverse opțiuni hemoglobinelor. Cel mai adesea, mutațiile sunt asociate cu înlocuirea unei nucleotide cu alta și apariția unui nou codon în genă, care poate codifica un nou aminoacid din lanțul polipeptidic al hemoglobinei. Într-un triplet, ca urmare a unei mutații, orice nucleotidă poate fi înlocuită - prima, a doua sau a treia. Se știe că câteva sute de mutații afectează integritatea genelor globinei. Aproape 400 dintre care sunt asociate cu înlocuirea nucleotidelor individuale în genă și substituția corespunzătoare a aminoacizilor în polipeptidă. Dintre acestea, numai 100 substituțiile duc la instabilitatea hemoglobinei și diferite tipuri de boli de la ușoare până la foarte severe. 300 (aproximativ 64%) mutații de substituție nu afectează funcția hemoglobinei și nu duc la patologie. Unul dintre motivele pentru aceasta este „degenerarea bazei a treia” menționată mai sus, când înlocuirea celei de-a treia nucleotide în tripletul care codifică serină, leucină, prolină, arginină și alți alți aminoacizi duce la apariția unui codon sinonim. care codifică același aminoacid. Fenotipic, o astfel de mutație nu se va manifesta. În schimb, orice înlocuire a primei sau celei de-a doua nucleotide într-un triplet în 100% din cazuri duce la apariția unei noi variante de hemoglobină. Dar chiar și în acest caz, este posibil să nu existe tulburări fenotipice severe. Motivul pentru aceasta este înlocuirea unui aminoacid din hemoglobină cu altul similar cu primul. proprietati fizice si chimice. De exemplu, dacă un aminoacid cu proprietăți hidrofile este înlocuit cu un alt aminoacid, dar cu aceleași proprietăți.
Hemoglobina este formată dintr-un grup de porfirină de fier de hem (la acesta sunt atașate moleculele de oxigen și dioxid de carbon) și o proteină - globină. Hemoglobina adultă (HbA) conține două identice - lanțuri și două -lanţuri. Moleculă - lanțul conține 141 de resturi de aminoacizi, - lanț - 146, - și -lanţurile diferă în multe resturi de aminoacizi. Secvența de aminoacizi a fiecărui lanț de globină este codificată de propria sa genă. Codificarea genei - lanțul este situat pe brațul scurt al cromozomului 16, -genă - în brațul scurt al cromozomului 11. Modificare a codificării genei - lanțul hemoglobinei din primul sau al doilea nucleotid duce aproape întotdeauna la apariția de noi aminoacizi în proteină, perturbarea funcțiilor hemoglobinei și consecințe grave pentru pacient. De exemplu, înlocuirea „C” într-unul dintre tripleții CAU (histidină) cu „U” va duce la apariția unui nou triplet UAU care codifică un alt aminoacid - tirozină. Fenotipic, aceasta se va manifesta într-o boală gravă.. A înlocuire similară în poziția 63 -lantul polipeptidei histidinei la tirozină va destabiliza hemoglobina. Se dezvoltă boala methemoglobinemie. Schimbarea, ca urmare a mutației, a acidului glutamic în valină în poziția a 6-a lanțul este cauza unei boli severe - anemia falciforme. Să nu continuăm lista tristă. Remarcăm doar că la înlocuirea primelor două nucleotide, un aminoacid poate apărea similar în proprietăți fizico-chimice cu cel anterior. Astfel, înlocuirea celei de-a doua nucleotide într-unul dintre tripleții care codifică acidul glutamic (GAA) în -lanțul pe „Y” duce la apariția unui nou triplet (GUA) care codifică valină, iar înlocuirea primei nucleotide cu „A” formează un triplet AAA care codifică aminoacidul lizină. Acidul glutamic și lizina sunt similare ca proprietăți fizico-chimice - ambele sunt hidrofile. Valina este un aminoacid hidrofob. Prin urmare, înlocuirea acidului glutamic hidrofil cu valină hidrofobă modifică semnificativ proprietățile hemoglobinei, ceea ce duce în cele din urmă la dezvoltarea anemiei falciforme, în timp ce înlocuirea acidului glutamic hidrofil cu lizină hidrofilă modifică într-o măsură mai mică funcția hemoglobinei - pacienții dezvolta o formă ușoară de anemie. Ca urmare a înlocuirii celei de-a treia baze, noul triplet poate codifica aceiași aminoacizi ca și precedentul. De exemplu, dacă uracilul a fost înlocuit cu citozină în tripletul CAH și a apărut un triplet CAC, atunci practic nu vor fi detectate modificări fenotipice la o persoană. Acest lucru este de înțeles, pentru că Ambele triplete codifică același aminoacid, histidina.
În concluzie, este oportun să subliniem că degenerarea codului genetic și degenerarea bazei a treia dintr-o poziție biologică generală sunt mecanisme de protecție care sunt încorporate în evoluție în structura unică a ADN-ului și ARN-ului.
în. Neambiguitate.
Fiecare triplet (cu excepția celor fără sens) codifică doar un aminoacid. Astfel, în direcția codon - aminoacid, codul genetic este lipsit de ambiguitate, în direcția aminoacid - codon - este ambiguu (degenerat).
lipsit de ambiguitate
codon aminoacid
degenerat
Și în acest caz, necesitatea lipsei de ambiguitate în codul genetic este evidentă. Într-o altă variantă, în timpul translației aceluiași codon, în lanțul proteic ar fi inserați diferiți aminoacizi și, ca urmare, s-ar forma proteine cu structuri primare diferite și funcții diferite. Metabolismul celulei ar trece la modul de operare „o genă – mai multe polipeptide”. Este clar că într-o astfel de situație funcția de reglare a genelor s-ar pierde complet.
g. Polaritatea
Citirea informațiilor din ADN și din ARNm are loc doar într-o singură direcție. Polaritatea este esențială pentru definirea structurilor de ordin superior (secundar, terțiar etc.). Mai devreme am vorbit despre faptul că structurile de ordin inferior determină structuri de ordin superior. Structura terțiară și structurile de ordin superior în proteine se formează imediat de îndată ce lanțul de ARN sintetizat se îndepărtează de molecula de ADN sau lanțul polipeptidic se îndepărtează de ribozom. În timp ce capătul liber al ARN-ului sau al polipeptidei capătă o structură terțiară, celălalt capăt al lanțului continuă să fie sintetizat pe ADN (dacă ARN-ul este transcris) sau ribozom (dacă polipeptida este transcrisă).
Prin urmare, procesul unidirecțional de citire a informațiilor (în sinteza ARN-ului și a proteinei) este esențial nu numai pentru determinarea secvenței nucleotidelor sau aminoacizilor din substanța sintetizată, ci și pentru determinarea rigidă a secundare, terțiare etc. structurilor.
e. Nesuprapunere.
Codul se poate suprapune sau nu. În majoritatea organismelor, codul nu se suprapune. Un cod care se suprapune a fost găsit la unii fagi.
Esența unui cod care nu se suprapun este că nucleotida unui codon nu poate fi nucleotida altui codon în același timp. Dacă codul s-ar suprapune, atunci secvența de șapte nucleotide (GCUGCUG) ar putea codifica nu doi aminoacizi (alanină-alanină) (Fig. 33, A) ca în cazul unui cod care nu se suprapun, ci trei (dacă o nucleotidă). este comun) (Fig. 33, B) sau cinci (dacă două nucleotide sunt comune) (vezi Fig. 33, C). În ultimele două cazuri, o mutație a oricărei nucleotide ar duce la o încălcare a secvenței de doi, trei etc. aminoacizi.
Cu toate acestea, s-a descoperit că o mutație a unei nucleotide perturbă întotdeauna includerea unui aminoacid într-o polipeptidă. Acesta este un argument semnificativ în favoarea faptului că codul nu se suprapune.
Să explicăm acest lucru în Figura 34. Liniile aldine arată tripleți care codifică aminoacizi în cazul codului care nu se suprapun și se suprapune. Experimentele au arătat fără ambiguitate că codul genetic nu se suprapune. Fără a intra în detaliile experimentului, observăm că dacă înlocuim a treia nucleotidă din secvența de nucleotide (vezi Fig. 34)La (marcat cu un asterisc) la altul atunci:
1. Cu un cod care nu se suprapune, proteina controlată de această secvență ar avea un înlocuitor pentru un (primul) aminoacid (marcat cu asteriscuri).
2. Cu un cod care se suprapune în opțiunea A, o înlocuire ar avea loc în doi (primul și al doilea) aminoacizi (marcați cu asteriscuri). În cadrul opțiunii B, substituția ar afecta trei aminoacizi (marcați cu asteriscuri).
Cu toate acestea, numeroase experimente au arătat că atunci când o nucleotidă din ADN este spartă, proteina afectează întotdeauna doar un aminoacid, ceea ce este tipic pentru un cod care nu se suprapune.
ГЦУГЦУГ ГЦУГЦУГ ГЦУГЦУГ
HCC HCC HCC UHC CUG HCC CUG UGC HCU CUG
*** *** *** *** *** ***
Alanine - Alanine Ala - Cys - Lei Ala - Lei - Lei - Ala - Lei
A B C
cod care nu se suprapune cod care se suprapune
Orez. 34. Schemă care explică prezența unui cod care nu se suprapune în genom (explicație în text).
Nesuprapunerea codului genetic este asociată cu o altă proprietate - citirea informației începe de la un anumit punct - semnalul de inițiere. Un astfel de semnal de inițiere în ARNm este codonul care codifică AUG metionina.
Trebuie remarcat faptul că o persoană are încă un număr mic de gene care deviază de la regula generalași se suprapun.
e. Compactitate.
Nu există semne de punctuație între codoni. Cu alte cuvinte, tripleții nu sunt separați unul de celălalt, de exemplu, printr-o singură nucleotidă fără sens. Absența „semnelor de punctuație” în codul genetic a fost dovedită în experimente.
și. Versatilitate.
Codul este același pentru toate organismele care trăiesc pe Pământ. Dovada directă a universalității codului genetic a fost obținută prin compararea secvențelor de ADN cu secvențele de proteine corespunzătoare. S-a dovedit că aceleași seturi de valori de cod sunt folosite în toți genomii bacterieni și eucarioți. Sunt excepții, dar nu multe.
Primele excepții de la universalitatea codului genetic s-au găsit în mitocondriile unor specii de animale. Aceasta se referă la codonul terminator UGA, care se citește la fel cu codonul UGG care codifică aminoacidul triptofan. Au fost găsite și alte abateri mai rare de la universalitate.
Sistemul de coduri ADN.
Codul genetic al ADN-ului este format din 64 de tripleți de nucleotide. Acești tripleți se numesc codoni. Fiecare codon codifică unul dintre cei 20 de aminoacizi utilizați în sinteza proteinelor. Acest lucru dă o oarecare redundanță în cod: majoritatea aminoacizilor sunt codificați de mai mult de un codon.
Un codon îndeplinește două funcții interdependente: semnalează începutul translației și codifică încorporarea aminoacidului metionină (Met) în lanțul polipeptidic în creștere. Sistemul de coduri ADN este conceput astfel încât codul genetic să poată fi exprimat fie ca codoni ARN, fie ca codoni ADN. Codonii ARN apar în ARN (ARNm) și acești codoni sunt capabili să citească informații în timpul sintezei polipeptidelor (un proces numit traducere). Dar fiecare moleculă de ARNm dobândește o secvență de nucleotide în transcripție de la gena corespunzătoare.
Toți, cu excepția a doi aminoacizi (Met și Trp) pot fi codificați de 2 până la 6 codoni diferiți. Cu toate acestea, genomul majorității organismelor arată că anumiți codoni sunt favorizați față de alții. La oameni, de exemplu, alanina este codificată de GCC de patru ori mai des decât în GCG. Acest lucru indică probabil o eficiență de traducere mai mare a aparatului de traducere (de exemplu, ribozomul) pentru unii codoni.

Codul genetic este aproape universal. Aceiași codoni sunt alocați aceleiași porțiuni de aminoacizi și aceleași semnale de pornire și oprire sunt în mare parte aceleași la animale, plante și microorganisme. Cu toate acestea, au fost găsite unele excepții. Cele mai multe dintre acestea implică atribuirea unuia sau doi dintre cei trei codoni stop unui aminoacid.
1. Codul este triplet.
2. Codul este degenerat.
3. Codul este lipsit de ambiguitate.
4. Codul este coliniar.
5. Codul nu se suprapune.
6. Codul este universal.
1) cod triplet. 3 nucleotide adiacente transportă informații despre o proteină. Pot exista 64 de astfel de tripleți (acest lucru arată redundanța codului genetic), dar doar 61 dintre ei poartă informații despre proteină (codoni). 3 tripleți se numesc anticodoni, sunt semnale de stop la care sinteza proteinelor se oprește.
2) Codul este degenerat. Un aminoacid poate fi codificat de mai mulți codoni.
3) Codul este lipsit de ambiguitate. Fiecare codon codifică doar un aminoacid.
4) Codul este coliniar. Secvența de nucleotide dintr-o genă corespunde secvenței de aminoacizi dintr-o proteină.
5) Codul nu se suprapune. aceeași nucleotidă nu poate face parte din doi codoni diferiți, citirea merge continuu, la rând, până la codonul stop. Nu există „semne de punctuație” în cod.
6) Codul este universal. Este același pentru toate ființele vii, adică. aceleași triplete codifică același aminoacid.
61. În ce cazuri o modificare a secvenței de nucleotide într-o genă nu afectează structura și funcțiile proteinei codificatoare?
1) dacă, ca urmare a unei substituții de nucleotide, apare un alt codon care codifică același aminoacid;
2) dacă codonul format ca urmare a unei substituții de nucleotide codifică un aminoacid diferit, dar cu similar proprietăți chimice, care nu modifică structura proteinei;
3) dacă apar modificări de nucleotide în regiunile ADN intergenetice sau nefuncționale.
№62. Replicarea ADN-ului.
replicare- procesul de sinteză a moleculei fiice deoxiribo acid nucleic pe șablonul moleculei de ADN părinte. În timpul diviziunii ulterioare a celulei mamă, fiecare celulă fiică primește o copie a unei molecule de ADN care este identică cu ADN-ul celulei mamă inițiale. Acest proces asigură transmiterea exactă a informațiilor genetice de la o generație la alta. Replicarea ADN-ului este realizată de un complex enzimatic complex, format din 15-20 de proteine diferite, numit replizom.
Până în momentul divizării, ADN-ul trebuie replicat complet și o singură dată. Replicarea are loc în trei etape:
1. Inițierea replicării (ADN-polimeraza începe replicarea ADN-ului prin legarea de un segment al unui lanț de nucleotide. La un anumit loc (punctul de început al replicării), are loc denaturarea locală a ADN-ului, lanțurile diverg și se formează două furculițe de replicare, mișcându-se. în direcţii opuse.).
2. Alungirea (o etapă a biosintezei moleculelor de acid nucleic, care constă în adăugarea secvenţială de monomeri (nucleotide) la un lanţ de ADN în creştere).
3. Terminarea replicării (etapa finală are loc în momentul în care zonele goale sunt umplute cu nucleotide între fragmentele Okazaki).
Parte principală:
Întrucât ADN-ul este molecula eredității, pentru a realiza această calitate, el trebuie să se copieze exact pe sine și astfel să păstreze toate informațiile disponibile în molecula originală de ADN sub forma unei anumite secvențe de nucleotide. Acest lucru este asigurat de un proces special care precede diviziunea oricărei celule din organism, care se numește replicare ADN - procesul de sinteză a unei molecule fiice de acid dezoxiribonucleic pe șablonul moleculei de ADN părinte.
Replicarea ADN-ului are loc în trei etape:
1. Iniţiere. Constă în faptul că enzimele speciale -ADN helicaze, care desfășoară spirala ADN-ului dublu catenar, rupe legăturile slabe de hidrogen care leagă nucleotidele celor două lanțuri. Ca rezultat, catenele de ADN sunt deconectate, iar bazele azotate libere „ie ies” din fiecare catenă (aspectul așa-numitei furculițe de replicare).
2. Elongaţie(o etapă a biosintezei moleculelor de acid nucleic, care constă în atașarea succesivă a monomerilor (nucleotide) la un lanț de ADN în creștere). Fiecare dintre cele două catene de ADN servește ca șablon pentru sinteza unei noi catene. Deoarece catenele parentale sunt antiparalele, replicarea continuă a ADN-ului are loc doar pe o singură catenă, care se numește lider (lider). O enzimă specială, ADN polimeraza, începe să se deplaseze de-a lungul lanțului de ADN liber de la capătul 5 „la capătul 3”, ajutând la atașarea nucleotidelor libere sintetizate în mod constant în celulă la capătul de 3” al catenei de ADN nou sintetizate. Sinteza a unei noi catene pe o catenă întârziată necesită formarea constantă de noi semințe (așa-numiții primeri - fragmente scurte de acid nucleic utilizate de ADN - polimeraze pentru a iniția sinteza ADN-ului) pentru a începe replicarea și se efectuează în segmente mici de 1000-2000 de nucleotide fiecare (fragmente Okazaki). Semințele se degradează după finalizarea sintezei următorului fragment Okazaki. Fragmentele de ADN adiacente rezultate sunt conectate prin ADN ligază. Topoizomeraza îndepărtează superbobinele, helicaza se desfășoară dublu helix, proteina SSB oferă stabilitate ADN-ului monocatenar.
3. Terminarea (finalizarea) replicării are loc atunci când golurile dintre fragmentele Okazaki sunt umplute cu nucleotide (cu participarea ADN ligazei) cu formarea a două catene duble continue de ADN și când două furculițe de replicare se întâlnesc. Apoi, ADN-ul sintetizat este răsucit pentru a forma superbobine.
63. Descrieți secvența proceselor care au loc în timpul replicării ADN-ului la eucariote
Mecanismele de replicare a ADN-ului procariotelor și eucariotelor diferă semnificativ prin aceea că, în al doilea caz, sinteza catenelor de ADN conducătoare și întârziate este realizată de diferite ADN polimeraze (alfa și respectiv delta), în timp ce la E. coli ambele catene de ADN sunt sintetizat de dimerul ADN polimerazei III. ADN polimeraza alfa inițiază sinteza catenei conducătoare în punctele de origine a replicării, iar ADN polimeraza delta realizează reinițializarea ciclică a sintezei fragmentelor Okazaki, recunoscând aparent prezența nucleotidei 5’-terminale a următorului primer, urmată de disociere. din ADN șablon și atașarea la acesta pentru reinițializarea sintezei următorului fragment Okazaki.
Maturarea fragmentelor Okazaki la eucariote necesită îndepărtarea primerilor ARN cu ajutorul 5’->3’-exonucleazei (factori proteici FEN-1 sau MF-1) și RNază H1, precum și atașarea covalentă a fragmentelor la reciproc sub acțiunea ADN ligazei I.
În prezent, nu se știe ce anume servește drept semnal de pornire pentru începerea replicării ADN-ului în faza S. Evenimentul de inițiere, după care începe sinteza ADN-ului, are loc în locuri specifice numite „furci de replicare”. În timpul fazei S, grupurile de furcuri de replicare sunt activate simultan pe toți cromozomii.
Poziția originilor replicării în gene poate fi importantă semnificație biologică. Faptul că un număr de virusuri animale încep replicarea la anumite locuri ale genomului sugerează că originile replicării sunt secvențe specializate în ADN-ul cromozomial. Distanța medie dintre originile de replicare este comparabilă cu distanța medie dintre buclele de cromatină adiacente. Astfel, este posibil să existe o singură origine de replicare în fiecare buclă.
Când două furculițe de replicare diverg de la un punct de pornire al replicării pe părțile opuse ale acestui punct, nucleozomii parentali vor cădea în diferite elice ADN copil. În acest caz, distribuția histonelor parentale preexistente între cele două gene fiice va depinde de locația exactă a originii replicării în unitatea de transcripție. Nu toți nucleozomii sunt exact la fel - în diferite zone ale materialului genetic, structura cromatinei este diferită. Poziția exactă a originii replicării într-o genă ar putea avea, prin urmare, o mare importanță biologică, deoarece ar determina structura cromatinei acelei gene în următoarea generație de celule.
Declanșatorul de replicare a ADN-ului funcționează în mod clar pe o bază de totul sau nimic, deoarece replicarea ADN-ului, care a început în faza S, continuă până la finalizarea acestui proces. Controlul procesului de replicare conform principiului „totul sau nimic” poate fi efectuat de cel puțin doi căi diferite:
1) unele sistem general poate recunoaște în mod specific fiecare bandă de cromozomi, o decondensează și, astfel, să facă toate originile de replicare accesibile simultan proteinelor responsabile de formarea bulelor de replicare;
2) proteinele replicative pot recunoaște doar câteva origini de replicare dintr-un set dat, după care replicarea locală care a început va schimba structura restului cromatinei unității de replicare în așa fel încât replicarea la toate celelalte origini să devină posibil.
Este posibil ca momentul critic din lanțul de evenimente care inițiază replicarea ADN-ului să fie realizarea unei anumite etape în procesul de dublare a centriolului, care acționează atât ca parte a unui important centru organizator al microtubulilor strâns asociat cu nucleul de interfaza, cât și ca componentă. a fiecăruia dintre polii fusului în timpul mitozei. Aparent, centriolul se dublează printr-un proces de matrice o dată pe ciclu celular (Fig. 11-19).
De asemenea, nu se știe încă ce determină secvența de replicare fixă a benzilor cromozomiale. Au fost propuse două ipoteze pentru a explica această secvență. Potrivit uneia dintre ele, diferite proteine replicative, fiecare dintre ele specifice pentru un anumit tip de benzi cromozomiale, sunt sintetizate în faza S în timp diferit. Conform unei alte ipoteze, care acum pare mai plauzibilă, proteinele replicative acționează pur și simplu asupra acelor părți ale ADN-ului care le sunt mai accesibile; de exemplu, în timpul fazei S, poate apărea decondensarea continuă a cromozomilor, iar benzile cromozomilor una câte una devin disponibile pentru proteinele replicative.