Mitose cellulaire. Division mitotique
Lire aussi
Manuel pour les classes 10-11
Section II. Reproduction et développement des organismes
Chapitre V. Reproduction des organismes
Chaque seconde sur Terre, un nombre astronomique d'êtres vivants meurent de vieillesse, de maladies et de prédateurs, et ce n'est que grâce à la reproduction, cette propriété universelle des organismes, que la vie sur Terre ne s'arrête pas.
Il peut sembler que les processus de reproduction chez les êtres vivants sont très divers, mais tous peuvent être réduits à deux formes : asexuée et sexuée. Certains organismes ont différentes formes reproduction. Par exemple, de nombreuses plantes peuvent être propagées par bouturage, marcottage, tubercules (reproduction asexuée) et graines (sexuée).
Au cours de la reproduction sexuée, chaque organisme se développe à partir d'une cellule, formée de la fusion de deux cellules germinales - mâle et femelle.
La base de la reproduction et du développement individuel du corps est le processus de division cellulaire.
§ 20. Division cellulaire. Mitose
La capacité à se diviser est la propriété la plus importante des cellules. Sans division, il est impossible d'imaginer une augmentation du nombre d'êtres unicellulaires, le développement d'un organisme multicellulaire complexe à partir d'un seul œuf fécondé, le renouvellement des cellules, des tissus et même des organes perdus au cours de la vie de l'organisme.
La division cellulaire s'effectue par étapes. A chaque étape de la division, certains processus se produisent. Ils conduisent au dédoublement du matériel génétique (synthèse d'ADN) et à sa répartition entre les cellules filles. La période de la vie d'une cellule d'une division à l'autre s'appelle le cycle cellulaire.
Préparation à la division. Les organismes eucaryotes, constitués de cellules avec des noyaux, commencent à se préparer à la division à un certain stade du cycle cellulaire, en interphase.
C'est pendant la période d'interphase dans la cellule que se produit le processus de biosynthèse des protéines, les chromosomes se doublent. Le long du chromosome d'origine à partir des composés chimiques présents dans la cellule, sa copie exacte est synthétisée, la molécule d'ADN est doublée. Un chromosome doublé se compose de deux moitiés - les chromatides. Chaque chromatide contient une molécule d'ADN.
L'interphase dans les cellules végétales et animales dure en moyenne 10 à 20 heures, puis vient le processus de division cellulaire - la mitose.
Au cours de la mitose, la cellule passe par une série de phases successives, à la suite desquelles chaque cellule fille reçoit le même ensemble de chromosomes que dans la cellule mère.
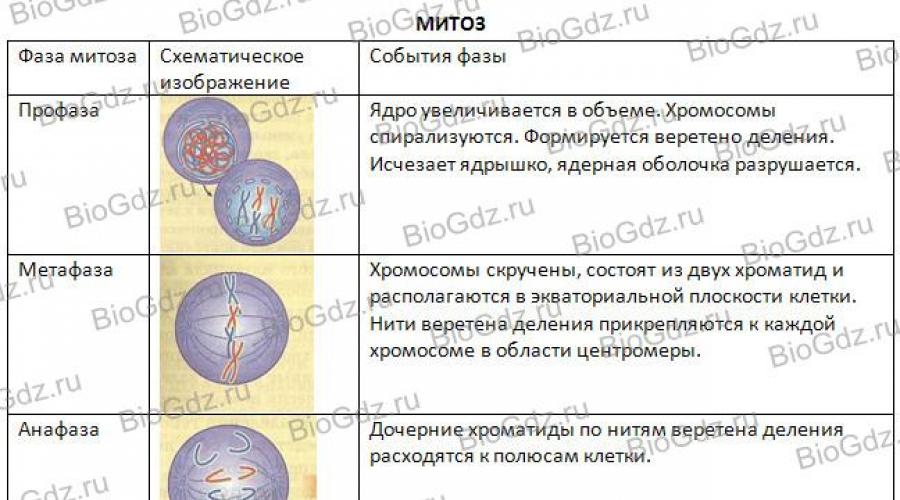
phases de la mitose. Il existe quatre phases de mitose : la prophase, la métaphase, l'anaphase et la télophase. La figure 29 montre schématiquement le déroulement de la mitose. En prophase, les centrioles sont clairement visibles - formations situées au centre de la cellule et jouant un rôle dans la divergence des chromosomes filles des animaux. (Rappelons que seules certaines plantes ont des centrioles au centre de la cellule, qui organisent la divergence des chromosomes.) Nous allons considérer la mitose en utilisant l'exemple cellule animale, car la présence d'un centriole rend plus visible le processus de ségrégation des chromosomes. Les centrioles doublent et divergent vers différents pôles de la cellule. Les microtubules s'étendent des centrioles, formant des fibres fusiformes, qui régulent la divergence des chromosomes vers les pôles de la cellule en division.
Riz. 29. Schéma de mitose
A la fin de la prophase, la membrane nucléaire se désagrège, le nucléole disparaît progressivement, les chromosomes se spiralisent et par conséquent se raccourcissent et s'épaississent, et ils peuvent déjà être observés au microscope optique. Ils sont encore mieux vus à l'étape suivante de la mitose - la métaphase.
En métaphase, les chromosomes sont situés dans le plan équatorial de la cellule. On voit clairement que chaque chromosome, composé de deux chromatides, a une constriction - le centromère. Les chromosomes sont attachés aux fibres du fuseau par leurs centromères. Après division du centromère, chaque chromatide devient un chromosome fille indépendant.
Vient ensuite l'étape suivante de la mitose - l'anaphase, au cours de laquelle les chromosomes filles (chromatides d'un chromosome) divergent vers différents pôles de la cellule.
La prochaine étape de la division cellulaire est la télophase. Il commence après que les chromosomes filles, constitués d'une chromatide, aient atteint les pôles de la cellule. A ce stade, les chromosomes déspiralisent à nouveau et acquièrent la même forme qu'ils avaient avant le début de la division cellulaire dans l'interphase (filaments longs et fins). Une enveloppe nucléaire se forme autour d'eux et un nucléole se forme dans le noyau, dans lequel les ribosomes sont synthétisés. Dans le processus de division du cytoplasme, tous les organites (mitochondries, complexe de Golgi, ribosomes, etc.) sont plus ou moins uniformément répartis entre les cellules filles.
Ainsi, à la suite de la mitose, deux cellules sont obtenues à partir d'une cellule, chacune ayant un nombre et une forme de chromosomes caractéristiques pour un type d'organisme donné et, par conséquent, une quantité constante d'ADN.
L'ensemble du processus de la mitose prend en moyenne 1 à 2 heures, sa durée est quelque peu différente pour différents types cellules. Cela dépend aussi des conditions environnement externe(température, régime lumineux et autres indicateurs).
L'importance biologique de la mitose réside dans le fait qu'elle assure la constance du nombre de chromosomes dans toutes les cellules de l'organisme. Dans le processus de mitose, l'ADN des chromosomes de la cellule mère est réparti de manière strictement égale entre les deux cellules filles qui en sont issues. À la suite de la mitose, toutes les cellules filles reçoivent la même information génétique.
- Quels changements dans la cellule précèdent la division ?
- Quand le fuseau est-il formé ? Quel est son rôle ?
- Décrivez les phases de la mitose et décrivez brièvement comment ce processus se produit.
- Qu'est-ce qu'une chromatide ? Quand devient-il un chromosome ?
- Qu'est-ce qu'un centromère ? Quel rôle joue-t-il dans la mitose ?
- Qu'est-ce que signification biologique mitose?
Rappelez-vous du cours de botanique, de zoologie, d'anatomie, de physiologie et d'hygiène humaine comment la reproduction se produit dans le monde organique.
1. Donner des définitions de concepts.
Interphase- la phase de préparation à la division mitotique, lors de la duplication de l'ADN.
Mitose- il s'agit d'une division, à la suite de laquelle il existe une répartition strictement identique des chromosomes exactement copiés entre les cellules filles, ce qui assure la formation de cellules génétiquement identiques.
Cycle de la vie
- la période de la vie cellulaire à partir du moment de son apparition dans le processus de division jusqu'à la mort ou la fin de la division ultérieure.
2. En quoi la croissance des organismes unicellulaires diffère-t-elle de la croissance des organismes multicellulaires ?
La croissance d'un organisme unicellulaire est une augmentation de la taille et de la complication de la structure d'une seule cellule, et la croissance d'organismes multicellulaires est également une division active des cellules - une augmentation de leur nombre.
3. Pourquoi une interphase existe-t-elle nécessairement dans le cycle de vie d'une cellule ?
Dans l'interphase, la préparation pour la division et la duplication de l'ADN se produit. Si cela ne se produisait pas, à chaque division cellulaire, le nombre de chromosomes serait divisé par deux et, très bientôt, il ne resterait plus du tout de chromosomes dans la cellule.
4. Renseignez le cluster "Phases of Mitosis".
5. A l'aide de la figure 52 du § 3.4, complétez le tableau.

6. Composez un syncwine pour le terme "mitose".
Mitose
Quatre phases, uniforme
Divise, distribue, divise
Fournit du matériel génétique aux cellules filles
la division cellulaire.
7. Établir une correspondance entre les phases du cycle mitotique et les événements qui s'y déroulent.
Étapes
1. Anaphase
2. Métaphase
3. Interphase
4. Télophase
5. Prophase
Développements
A. La cellule se développe, des organites se forment, l'ADN se double.
B. Les chromatides se séparent et deviennent des chromosomes indépendants.
B. La spiralisation des chromosomes commence, l'enveloppe nucléaire est détruite.
D. Les chromosomes sont situés dans le plan équatorial de la cellule. Les fibres du fuseau se fixent aux centromères.
D. Le fuseau de division disparaît, les membranes nucléaires se forment, les chromosomes se déroulent.
8. Pourquoi l'achèvement de la mitose - la division du cytoplasme se produit-il différemment dans les cellules animales et végétales ?
Pas dans les cellules animales paroi cellulaire, ils ont membrane cellulaire pousse vers l'intérieur, et la cellule se divise par constriction.
Dans les cellules végétales, la membrane se forme dans le plan équatorial à l'intérieur de la cellule et, s'étendant à la périphérie, divise la cellule en deux.
9. Pourquoi l'interphase prend-elle beaucoup plus de temps dans le cycle mitotique que la division elle-même ?
Pendant l'interphase, la cellule se prépare intensément à la mitose, des processus de synthèse s'y déroulent, l'ADN se double, la cellule se développe, la traverse. cycle de la vie, sans compter la division elle-même.
10. Choisissez la bonne réponse.
Essai 1
À la suite de la mitose, une cellule diploïde produit :
4) 2 cellules diploïdes.
Essai 2
La division du centromère et la divergence des chromatides aux pôles de la cellule se produisent dans :
3) anaphase ;
Essai 3
Le cycle de vie est :
2) vie cellulaire de la division à la fin de la prochaine division ou mort ;
Essai 4
Quel terme est mal orthographié ?
4) télophase.
11. Expliquez l'origine et sens général mot (terme), basé sur le sens des racines qui le composent.

12. Choisissez un terme et expliquez comment son sens moderne correspond au sens originel de ses racines.
Le terme choisi est interphase.
Conformité. Le terme correspond à et signifie la période entre les phases de la mitose, lorsque se produit la préparation de la division.
13. Formulez et notez les idées principales du § 3.4.
Le cycle de vie est la vie d'une cellule de la division à la fin de la prochaine division ou de la mort. Entre les divisions, la cellule s'y prépare pendant l'interphase. A ce moment, il y a synthèse de substances, duplication d'ADN.
La cellule se divise par mitose. Il se compose de 4 étapes :
Prophase.
Métaphase.
Anaphase.
Télophase.
Le but de la mitose: à la suite de cela, 2 cellules filles avec un ensemble identique de gènes sont formées à partir d'une cellule mère. La quantité de matériel génétique et de chromosomes reste la même, assurant la stabilité génétique des cellules.
Le composant le plus important du cycle cellulaire est le cycle mitotique (prolifératif). C'est un complexe de phénomènes interdépendants et coordonnés au cours de la division cellulaire, ainsi qu'avant et après celle-ci. Le cycle mitotique est un ensemble de processus se produisant dans une cellule d'une division à l'autre et se terminant par la formation de deux cellules de la génération suivante. Par ailleurs, la notion de cycle de vie inclut également la période d'exécution par la cellule de ses fonctions et les périodes de repos. À ce stade, le destin ultérieur de la cellule est incertain : la cellule peut commencer à se diviser (entrer en mitose) ou commencer à se préparer à remplir des fonctions spécifiques.
Principales étapes de la mitose
1.
Reduplication (auto-doublement) de l'information génétique de la cellule mère et sa distribution uniforme entre les cellules filles. Cela s'accompagne de modifications de la structure et de la morphologie des chromosomes, dans lesquels se concentrent plus de 90% de l'information d'une cellule eucaryote.
2.
Le cycle mitotique comprend quatre périodes successives : G1 présynthétique (ou postmitotique), S synthétique, G2 postsynthétique (ou prémitotique) et la mitose proprement dite. Ils constituent l'interphase autocatalytique (période préparatoire).
Phases du cycle cellulaire :
1) présynthétique (G1). Se produit immédiatement après la division cellulaire. La synthèse d'ADN n'a pas encore eu lieu. La cellule grossit activement, stocke les substances nécessaires à la division: protéines (histones, protéines structurelles, enzymes), ARN, molécules d'ATP. Il existe une division des mitochondries et des chloroplastes (c'est-à-dire des structures capables d'autoreproduction). Les caractéristiques de l'organisation de la cellule interphase sont restaurées après la division précédente;
2)
synthétique (S). Le matériel génétique est dupliqué par réplication de l'ADN. Elle se produit de manière semi-conservative, lorsque la double hélice de la molécule d'ADN diverge en deux brins et qu'un brin complémentaire est synthétisé sur chacun d'eux.
Le résultat est deux identiques doubles hélices ADN, dont chacun se compose d'un nouveau et vieille chaîne ADN. La quantité de matériel héréditaire est doublée. De plus, la synthèse d'ARN et de protéines se poursuit. De plus, une petite partie de l'ADN mitochondrial subit une réplication (sa partie principale est répliquée dans la période G2) ;
3) post-synthétique (G2). L'ADN n'est plus synthétisé, mais il y a une correction des défauts apportés lors de sa synthèse en période S (réparation). Ils accumulent également de l'énergie et nutriments, la synthèse d'ARN et de protéines (principalement nucléaires) se poursuit.
S et G2 sont directement liés à la mitose, ils sont donc parfois isolés dans une période distincte - la préprophase.
Ceci est suivi par la mitose elle-même, qui se compose de quatre phases. Le processus de division comprend plusieurs phases successives et est un cycle. Sa durée est différente et varie de 10 à 50 heures dans la plupart des cellules.Dans le même temps, dans les cellules du corps humain, la durée de la mitose elle-même est de 1 à 1,5 heure, la période d'interphase G2 est de 2 à 3 heures, la La période S de l'interphase est de 6 à 10 heures.
La durée des étapes individuelles est différente et varie selon le type de tissu, état physiologique organisme, facteurs externes. Les étapes les plus longues sont associées aux processus de synthèse intracellulaire : prophase et télophase. Les phases les plus fugaces de la mitose, au cours desquelles se produit le mouvement des chromosomes : métaphase et anaphase. Le processus réel de divergence chromosomique vers les pôles ne dépasse généralement pas 10 minutes.
Prophase
Les principaux événements de la prophase comprennent la condensation des chromosomes dans le noyau et la formation d'un fuseau de fission dans le cytoplasme de la cellule. La désintégration du nucléole en prophase est une caractéristique, mais pas obligatoire pour toutes les cellules.
Classiquement, le moment d'apparition des chromosomes visibles au microscope en raison de la condensation de la chromatine intranucléaire est considéré comme le début de la prophase. Le compactage des chromosomes se produit en raison de l'hélice à plusieurs niveaux de l'ADN. Ces modifications s'accompagnent d'une augmentation de l'activité des phosphorylases qui modifient les histones directement impliquées dans l'assemblage de l'ADN. En conséquence, l'activité transcriptionnelle de la chromatine diminue fortement, les gènes nucléolaires sont inactivés et la plupart des protéines nucléolaires se dissocient. Les chromatides sœurs condensées au début de la prophase restent appariées sur toute leur longueur à l'aide de protéines cohésines, cependant, au début de la prométaphase, la connexion entre les chromatides n'est préservée que dans la région du centromère. À la fin de la prophase, des kinétochores matures se forment sur chaque centromère des chromatides soeurs, qui sont nécessaires pour que les chromosomes se fixent aux microtubules du fuseau dans la prométaphase.
Parallèlement aux processus de condensation intranucléaire des chromosomes, le fuseau mitotique commence à se former dans le cytoplasme - l'une des principales structures de l'appareil de division cellulaire responsable de la distribution des chromosomes entre les cellules filles. Dans la formation du fuseau de division dans toutes les cellules eucaryotes, les corps polaires, les microtubules et les kinétochores des chromosomes participent.
Avec le début de la formation du fuseau mitotique en prophase, des changements spectaculaires dans les propriétés dynamiques des microtubules sont associés. La demi-vie d'un microtubule moyen diminue d'environ 20 fois de 5 minutes à 15 secondes. Cependant, leur taux de croissance augmente d'environ 2 fois par rapport aux mêmes microtubules interphases. Les extrémités plus polymérisantes sont "dynamiquement instables" et passent brusquement d'une croissance uniforme à un raccourcissement rapide, qui dépolymérise souvent l'ensemble du microtubule. Il est à noter que pour le bon fonctionnement du fuseau mitotique, un certain équilibre est nécessaire entre les processus d'assemblage et de dépolymérisation des microtubules, puisque ni les microtubules du fuseau stabilisés ni dépolymérisés ne sont capables de déplacer les chromosomes.
Parallèlement aux changements observés dans les propriétés dynamiques des microtubules qui composent les filaments du fuseau, des pôles de fission se forment dans la prophase. Les centrosomes répliqués en phase S divergent dans des directions opposées en raison de l'interaction des microtubules polaires se développant les uns vers les autres. Avec leurs extrémités négatives, les microtubules sont immergés dans la substance amorphe des centrosomes et les processus de polymérisation se déroulent du côté des extrémités positives faisant face au plan équatorial de la cellule. Parallèlement, le mécanisme probable de la divergence des pôles s'explique comme suit : les protéines de type dynéine sont orientées dans sens parallèle polymérisant les extrémités positives des microtubules polaires et les protéines de type kinésine, à leur tour, les poussent vers les pôles de division.
Parallèlement à la condensation des chromosomes et à la formation du fuseau mitotique, au cours de la prophase, se produit une fragmentation du réticulum endoplasmique qui se décompose en petites vacuoles, qui divergent ensuite vers la périphérie cellulaire. Dans le même temps, les ribosomes perdent le contact avec les membranes du RE. Les citernes de l'appareil de Golgi changent également leur localisation périnucléaire, se désintégrant en dictyosomes séparés, distribués dans le cytoplasme sans ordre particulier.
prométaphase
La fin de la prophase et le début de la prométaphase sont généralement marqués par la désintégration de la membrane nucléaire. Toute la ligne les protéines de la lamina sont phosphorylées, à la suite de quoi l'enveloppe nucléaire est fragmentée en petites vacuoles et les complexes de pores disparaissent. Après la destruction de la membrane nucléaire, les chromosomes sont disposés au hasard dans la région du noyau. Cependant, bientôt ils commencent tous à bouger.
En prométaphase, un mouvement intensif mais aléatoire des chromosomes est observé. Initialement, les chromosomes individuels dérivent rapidement vers le pôle le plus proche du fuseau mitotique à une vitesse pouvant atteindre 25 µm/min. Près des pôles de division, la probabilité d'interaction des extrémités positives nouvellement synthétisées des microtubules du fuseau avec les kinétochores chromosomiques augmente. À la suite de cette interaction, les microtubules du kinétochore sont stabilisés de la dépolymérisation spontanée et leur croissance assure en partie la distance du chromosome qui leur est connecté dans la direction du pôle au plan équatorial du fuseau. D'autre part, le chromosome est dépassé par des brins de microtubules provenant du pôle opposé du fuseau mitotique. Interagissant avec le kinétochore, ils participent également au mouvement du chromosome. En conséquence, les chromatides sœurs sont associées aux pôles opposés du fuseau. La force développée par les microtubules de différents pôles non seulement stabilise l'interaction de ces microtubules avec les kinétochores, mais aussi, in fine, amène chaque chromosome dans le plan de la plaque métaphasique.
Dans les cellules de mammifères, la prométaphase se déroule généralement en 10 à 20 minutes. Dans les neuroblastes de sauterelle, cette étape ne prend que 4 minutes, alors que dans l'endosperme d'Haemanthus et les fibroblastes de triton, elle prend environ 30 minutes.
métaphase
En fin de prométaphase, les chromosomes sont situés dans le plan équatorial du fuseau approximativement à égale distance des deux pôles de division, formant une plaque métaphasique. La morphologie de la plaque métaphasique dans les cellules animales se distingue généralement par une disposition ordonnée des chromosomes: les régions centromériques font face au centre du fuseau et les épaules font face à la périphérie de la cellule. Dans les cellules végétales, les chromosomes se situent souvent dans le plan équatorial du fuseau sans ordre strict.
La métaphase occupe une partie importante de la période de mitose et se caractérise par un état relativement stable. Pendant tout ce temps, les chromosomes sont maintenus dans le plan équatorial du fuseau en raison des forces de tension équilibrées des microtubules du kinétochore, effectuant des mouvements oscillatoires de faible amplitude dans le plan de la plaque métaphasique.
En métaphase, ainsi que pendant d'autres phases de la mitose, le renouvellement actif des microtubules du fuseau se poursuit par l'assemblage intensif et la dépolymérisation des molécules de tubuline. Malgré une certaine stabilisation des faisceaux de microtubules de kinétochore, il existe un tri constant des microtubules interpolaires, dont le nombre en métaphase atteint un maximum.
À la fin de la métaphase, on observe une nette séparation des chromatides sœurs, dont la connexion n'est préservée que dans les régions centromériques. Les bras des chromatides sont disposés parallèlement les uns aux autres et l'espace qui les sépare devient clairement visible.
Anaphase
L'anaphase est l'étape la plus courte de la mitose, qui commence par la séparation soudaine et la séparation subséquente des chromatides sœurs vers les pôles opposés de la cellule. Les chromatides se séparent à une vitesse uniforme allant jusqu'à 0,5-2 µm/min, et elles prennent souvent une forme en V. Leur mouvement est dû à l'action de forces importantes, estimées à 10 dynes par chromosome, soit 10 000 fois plus que la force nécessaire pour déplacer simplement le chromosome dans le cytoplasme à la vitesse observée.
En règle générale, la ségrégation des chromosomes en anaphase consiste en deux processus relativement indépendants appelés anaphase A et anaphase B.
L'anaphase A est caractérisée par la séparation des chromatides sœurs aux pôles opposés de la division cellulaire. Dans ce cas, les mêmes forces qui maintenaient auparavant les chromosomes dans le plan de la plaque métaphasique sont responsables de leur mouvement. Le processus de séparation des chromatides s'accompagne d'un raccourcissement de la longueur des microtubules de kinétochore en dépolymérisation. De plus, leur désintégration est observée principalement dans la région des kinétochores, du côté des extrémités positives. Probablement, la dépolymérisation des microtubules au niveau des kinétochores ou dans la région des pôles de division est condition nécessaire pour déplacer les chromatides sœurs, car leur mouvement s'arrête lorsque du taxol ou de l'eau lourde est ajoutée, ce qui a un effet stabilisateur sur les microtubules. Le mécanisme sous-jacent à la ségrégation chromosomique dans l'anaphase A est encore inconnu.
Au cours de l'anaphase B, les pôles de la division cellulaire eux-mêmes divergent et, contrairement à l'anaphase A, ce processus se produit en raison de l'assemblage de microtubules polaires à partir des extrémités positives. Les fils antiparallèles polymérisants de la broche, lorsqu'ils interagissent, créent en partie la force qui écarte les pôles. L'ampleur du mouvement relatif des pôles dans ce cas, ainsi que le degré de chevauchement des microtubules polaires dans la zone équatoriale de la cellule, varient considérablement chez les individus d'espèces différentes. En plus des forces répulsives, les pôles de division sont affectés par les forces de traction des microtubules astraux, qui sont créées à la suite de l'interaction avec des protéines de type dynéine sur la membrane plasmique de la cellule.
La séquence, la durée et la contribution relative de chacun des deux processus qui composent l'anaphase peuvent être extrêmement différentes. Ainsi, dans les cellules de mammifères, l'anaphase B commence immédiatement après le début de la divergence des chromatides aux pôles opposés et se poursuit jusqu'à l'allongement du fuseau mitotique de 1,5 à 2 fois par rapport à celui de la métaphase. Dans certaines autres cellules, l'anaphase B ne commence qu'après que les chromatides ont atteint les pôles de division. Chez certains protozoaires, au cours de l'anaphase B, le fuseau s'allonge 15 fois par rapport à la métaphase. L'anaphase B est absente des cellules végétales.
Télophase
La télophase est considérée comme l'étape finale de la mitose ; son début est considéré comme le moment où les chromatides sœurs séparées s'arrêtent aux pôles opposés de la division cellulaire. Au début de la télophase, on observe une décondensation des chromosomes et, par conséquent, leur augmentation de volume. A proximité des chromosomes individuels groupés, commence la fusion des vésicules membranaires, qui donne lieu à la reconstruction de l'enveloppe nucléaire. Le matériau pour la construction des membranes des noyaux filles nouvellement formés sont des fragments de la membrane nucléaire initialement décomposée de la cellule mère, ainsi que des éléments du réticulum endoplasmique. Dans ce cas, les vésicules individuelles se lient à la surface des chromosomes et fusionnent. Les membranes nucléaires externe et interne sont progressivement restaurées, la lame nucléaire et les pores nucléaires sont restaurés. Dans le processus de réparation de l'enveloppe nucléaire, des vésicules membranaires discrètes se connectent probablement à la surface des chromosomes sans reconnaître des séquences nucléotidiques spécifiques, puisque des expériences ont montré que la réparation de la membrane nucléaire se produit autour de molécules d'ADN empruntées à n'importe quel organisme, même à un virus bactérien. À l'intérieur des noyaux cellulaires nouvellement formés, la chromatine passe dans un état dispersé, la synthèse d'ARN reprend et les nucléoles deviennent visibles.
Parallèlement aux processus de formation des noyaux des cellules filles dans la télophase, le désassemblage des microtubules du fuseau de fission commence et se termine. La dépolymérisation se déroule dans le sens des pôles de division vers le plan équatorial de la cellule, des extrémités négatives vers les extrémités positives. Dans le même temps, les microtubules sont stockés le plus longtemps dans la partie médiane du fuseau, qui forment le corps de Fleming résiduel.
La fin de la télophase coïncide principalement avec la division du corps de la cellule mère - la cytokinèse. Dans ce cas, deux ou plusieurs cellules filles sont formées. Les processus conduisant à la division du cytoplasme commencent dès le milieu de l'anaphase et peuvent se poursuivre après la fin de la télophase. La mitose n'est pas toujours accompagnée d'une division du cytoplasme, de sorte que la cytokinèse n'est pas classée comme une phase distincte de la division mitotique et est généralement considérée comme faisant partie de la télophase.
Il existe deux principaux types de cytokinèse : la division par la constriction transversale de la cellule et la division par la formation d'une plaque cellulaire. Le plan de division cellulaire est déterminé par la position du fuseau mitotique et s'étend perpendiculairement au grand axe du fuseau.
Lors de la division par une constriction transversale de la cellule, le site de division du cytoplasme est préalablement défini pendant la période d'anaphase, lorsqu'un anneau contractile de filaments d'actine et de myosine apparaît dans le plan de la plaque métaphasique sous la membrane cellulaire. Par la suite, en raison de l'activité de l'anneau contractile, un sillon de fission se forme, qui s'approfondit progressivement jusqu'à ce que la cellule soit complètement divisée. À la fin de la cytokinèse, l'anneau contractile se désintègre complètement et la membrane plasmique se contracte autour du corps de Fleming résiduel, qui consiste en une accumulation de restes de deux groupes de microtubules polaires étroitement emballés avec un matériau de matrice dense.
La division par la formation d'une plaque cellulaire commence par le mouvement de petites vésicules limitées par la membrane vers le plan équatorial de la cellule. Ici, ils fusionnent pour former une structure en forme de disque entourée d'une membrane, la plaque cellulaire précoce. Les petites vésicules proviennent principalement de l'appareil de Golgi et se déplacent vers le plan équatorial le long des microtubules polaires résiduels du fuseau, formant une structure cylindrique appelée phragmoplaste. Au fur et à mesure que la plaque cellulaire se dilate, les microtubules du phragmoplaste précoce se déplacent simultanément vers la périphérie cellulaire, où, en raison de nouvelles vésicules membranaires, la croissance de la plaque cellulaire se poursuit jusqu'à sa fusion finale avec la membrane de la cellule mère. Après la séparation finale des cellules filles, des microfibrilles de cellulose sont déposées dans la plaque cellulaire, complétant la formation d'une paroi cellulaire rigide.
Pour déterminer l'achèvement de chaque phase du cycle cellulaire, il est nécessaire d'y avoir des points de contrôle. Si la cellule "passe" le point de contrôle, elle continue à "se déplacer" dans le cycle cellulaire. Si certaines circonstances, telles que des dommages à l'ADN, empêchent la cellule de passer par un point de contrôle, qui peut être comparé à une sorte de point de contrôle, alors la cellule s'arrête et une autre phase du cycle cellulaire ne se produit pas, du moins jusqu'à ce que les obstacles soient supprimés. , empêchant la cage de passer par le poste de contrôle.
Division cellulaire mitotique
Mitose(du grec. Mitos - fil), également appelé caryocinèse, ou division cellulaire indirecte, est un mécanisme universel de division cellulaire. La mitose suit la période G2 et complète le cycle cellulaire.
Il dure 1 à 3 heures et assure une distribution uniforme du matériel génétique dans les cellules filles. La mitose comporte 4 phases principales : la prophase, la métaphase, l'anaphase et la télophase.
La mitose est l'un des processus fondamentaux de l'ontogenèse. La division mitotique assure la croissance des eucaryotes multicellulaires en augmentant les populations de cellules tissulaires.
À la suite de la division mitotique des cellules du méristème, le nombre de cellules de tissus végétaux augmente. La fragmentation d'un œuf fécondé et la croissance de la plupart des tissus chez les animaux se produisent également par divisions mitotiques.
Sur la base des caractéristiques morphologiques, la mitose est conditionnellement subdivisée en étapes: prophase, prométaphase, métaphase, anaphase, télophase. Les premières descriptions des phases de la mitose et l'établissement de leur séquence ont été entreprises dans les années 70-80 du XIXème siècle. À la fin des années 1870, l'histologue allemand Walter Flemming a inventé le terme «mitose» pour désigner le processus de division cellulaire indirecte.
La durée moyenne de la mitose est de 1 à 2 heures. La mitose des cellules animales dure généralement 30 à 60 minutes et les plantes - 2 à 3 heures. Depuis 70 ans, environ 10 14 divisions cellulaires sont réalisées dans le corps humain au total.
Les premières descriptions incomplètes concernant le comportement et les changements des noyaux dans les cellules en division se trouvent dans les travaux des scientifiques au début des années 1870.
Dans les travaux du botaniste russe Russov, datés de 1872, les plaques de métaphase et d'anaphase, constituées de chromosomes individuels, sont clairement décrites et représentées.
Un an plus tard, le zoologiste allemand G.A. Schneider est encore plus clair et cohérent, mais, bien sûr, n'a pas tout à fait décrit la division mitotique en utilisant l'exemple de l'écrasement des œufs de la turbellaire rectale Mesostomum. Dans son œuvre, pour l'essentiel, décrite et illustrée dans séquence correcte phases principales de la mitose : prophase, métaphase, anaphase (précoce et tardive). En 1874, le botaniste moscovite I.D. Chistyakov a également observé des phases distinctes de division cellulaire dans les spores de lycopodes et de prêles. Malgré les premiers succès, ni Russov, ni Schneider, ni Chistyakov n'ont pu donner une description claire et cohérente de la division mitotique.
En 1875, des articles contenant des descriptions plus détaillées des mitoses ont été publiés. O. Byuchli a décrit les modèles cytologiques dans les œufs broyés des ascaris et des mollusques et dans les cellules spermatogènes des insectes.
E. Strasburger a étudié la division mitotique dans les cellules de l'algue verte spirogyra, dans les cellules mères du pollen d'oignon et dans les cellules mères des spores de la lycopode. Se référant aux travaux d'O. Buechli et s'appuyant sur ses propres recherches, E. Strasburger a attiré l'attention sur l'unité des processus de division cellulaire dans les cellules végétales et animales.
À la fin de 1878 - début de 1879, il est apparu travaux détaillés Schleicher et W. Flemming. Dans son ouvrage de 1879, Schleicher proposa le terme « caryocinèse » pour désigner les processus complexes de division cellulaire, impliquant le mouvement parties constitutives graines. Walter Flemming a été le premier à introduire le terme «mitose» pour désigner la division cellulaire indirecte, qui est ensuite devenue généralement acceptée. Flemming possède également la formulation finale de la définition de la mitose en tant que processus cyclique, aboutissant à la division des chromosomes entre les cellules filles.
En 1880 O.V. Baranetsky a établi la structure hélicoïdale des chromosomes. Au cours de recherches ultérieures, des idées sur la spiralisation et la déspiralisation des chromosomes au cours du cycle mitotique ont été développées.
Au début des années 1900, les chromosomes ont été identifiés comme porteurs informations héréditaires, qui explique plus en détail rôle biologique la mitose, qui consiste en la formation de cellules filles génétiquement identiques.
Dans les années 1970, le décodage et l'étude détaillée des régulateurs de la division mitotique ont commencé, grâce à une série d'expériences sur la fusion de cellules à différents stades du cycle cellulaire. Dans ces expériences, lorsqu'une cellule en phase M était combinée avec une cellule à l'un des stades de l'interphase (G1, S ou G2), les cellules de l'interphase passaient à l'état mitotique (la condensation chromosomique commençait et l'enveloppe nucléaire se désintégrait) .
En conséquence, il a été conclu qu'il existe un facteur (ou des facteurs) dans la cellule cytoplasmique qui stimule la mitose, ou, en d'autres termes, le facteur M-stimulant (MSF, de l'anglais M-phase-promoting factor, MPF) .
Pour la première fois, le "facteur stimulant la mitose" a été découvert dans des œufs matures non fécondés de grenouille à griffes, qui se trouvent dans la phase M du cycle cellulaire. Le cytoplasme d'un tel ovule, injecté dans l'ovocyte, a conduit à une transition prématurée vers la phase M et au début de la maturation de l'ovocyte (à l'origine, l'abréviation MPF signifie Maturation Promoting Factor, qui se traduit par "maturation facteur promoteur »). Au cours d'autres expériences, une valeur universelle a été établie et en même temps haut degré le conservatisme du «facteur stimulant la mitose»: des extraits préparés à partir de cellules mitotiques d'une grande variété d'organismes, lorsqu'ils sont introduits dans des ovocytes de grenouille à griffes, les transfèrent en phase M.
Des études ultérieures ont révélé que le facteur qui stimule la mitose est un complexe hétérodimérique constitué d'une protéine cycline et d'une protéine kinase cycline-dépendante. La cycline est protéine régulatrice et se retrouve chez tous les eucaryotes. Sa concentration augmente périodiquement au cours du cycle cellulaire, atteignant un maximum dans la métaphase de la mitose. Avec le début de l'anaphase, une forte diminution de la concentration de cycline est observée, en raison de son clivage à l'aide de complexes protéolytiques complexes de protéines - protéosomes. La protéine kinase cycline-dépendante est une enzyme (phosphorylase) qui modifie les protéines en transférant un groupe phosphate de l'ATP aux acides aminés sérine et thréonine. Ainsi, avec l'établissement du rôle et de la structure du principal régulateur de la division mitotique, des études sur les mécanismes régulateurs subtils de la mitose ont commencé, qui se poursuivent à ce jour.
Le développement d'une typologie et d'une classification unifiées des mitoses est compliqué par toute une gamme de caractéristiques qui, dans diverses combinaisons, créent une variété et une hétérogénéité de modèles de division mitotique. Dans le même temps, les options de classification distinctes développées pour certains taxons sont inacceptables pour d'autres, car elles ne tiennent pas compte des spécificités de leurs mitoses. Par exemple, certaines variantes de la classification des mitoses, caractéristiques des organismes animaux ou végétaux, s'avèrent inacceptables pour les algues.
L'une des principales caractéristiques sous-jacentes aux différentes typologies et classifications de la division mitotique est le comportement de l'enveloppe nucléaire. Si la formation du fuseau et la division mitotique elle-même se déroulent à l'intérieur du noyau sans détruire la membrane nucléaire, alors ce type de mitose est appelé fermé. La mitose avec l'effondrement de l'enveloppe nucléaire, respectivement, est appelée ouverte, et la mitose avec l'effondrement de la membrane uniquement aux pôles du fuseau, avec la formation de "fenêtres polaires" - semi-fermées.
Une autre caractéristique est le type de symétrie du fuseau mitotique. Dans la pleuromitose, le fuseau de division est bilatéralement symétrique ou asymétrique et se compose généralement de deux demi-fuseaux situés dans la métaphase-anaphase à un angle l'un par rapport à l'autre. La catégorie des orthomitoses est caractérisée par une symétrie bipolaire du fuseau de fission et, en métaphase, il existe souvent une plaque équatoriale distincte.
Parmi les signes indiqués, le plus nombreux est une orthomitose ouverte typique, à l'exemple de laquelle les principes et les étapes de la division mitotique sont discutés ci-dessous. Ce type de mitose est caractéristique des animaux, des plantes supérieures et de certains protozoaires.
La prophase commence par la condensation des chromosomes, qui deviennent visibles au microscope optique sous forme de structures filamenteuses. Chaque chromosome est constitué de deux chromatides sœurs parallèles reliées au centromère. Le nucléole et l'enveloppe nucléaire disparaissent à la fin de la phase (cette dernière se décompose en vésicules membranaires similaires aux éléments EPS, et le complexe de pores et la lamina se dissocient en sous-unités). Le caryoplasme se mélange au cytoplasme.
Les centrioles migrent vers les pôles opposés de la cellule et donnent naissance aux filaments du fuseau mitotique (achromatine). Dans la région du centromère, des complexes protéiques spéciaux se forment - des kinétochores, auxquels sont attachés certains microtubules de fuseau (microtubules de kinétochore); il a été démontré que les kinétochores eux-mêmes sont capables d'induire l'assemblage des microtubules et peuvent donc servir de centres d'organisation des microtubules. Le reste des microtubules du fuseau est appelé microtubules polaires, car ils s'étendent d'un pôle à l'autre de la cellule; les microtubules situés à l'extérieur du fuseau, divergeant radialement des centres cellulaires vers le plasmalemme, ont reçu le nom d'astral ou de microtubules (fils) de rayonnement.
La métaphase correspond au niveau maximal de condensation des chromosomes, qui s'alignent dans la région équatoriale du fuseau mitotique, formant une image de la plaque équatoriale (métaphase) (vue de côté) ou de l'étoile mère (vue depuis les pôles). Les chromosomes se déplacent vers le plan équatorial et y sont maintenus en raison de la tension équilibrée des microtubules du kinétochore. À la fin de cette phase, les chromatides soeurs sont séparées par un espace, mais sont retenues dans la région du centromère.
L'anaphase commence par la division synchrone de tous les chromosomes en chromatides sœurs (dans la région du centromère) et le mouvement des chromosomes filles vers les pôles opposés de la cellule, qui se produit le long des microtubules du fuseau à une vitesse de 0,2 à 0,5 µm/min. Le signal d'apparition de l'anaphase comprend une forte augmentation (d'un ordre de grandeur) de la concentration de cations calcium dans l'hyaloplasme, sécrétée par des vésicules membranaires qui forment des amas aux pôles du fuseau. Le mécanisme du mouvement des chromosomes en anaphase n'a pas été complètement élucidé, cependant, il a été établi qu'en plus de l'actine, des protéines telles que la myosine et la dynéine, ainsi qu'un certain nombre de protéines régulatrices, sont présentes dans la région du fuseau. Selon certaines observations, il est dû au raccourcissement (désassemblage) des microtubules attachés aux kinétochores. L'anaphase est caractérisée par un allongement du fuseau mitotique dû à une certaine divergence des pôles cellulaires. Il se termine par l'accumulation de deux ensembles identiques de chromosomes aux pôles de la cellule, qui forment des images d'étoiles (le stade des étoiles filles). A la fin de l'anaphase, en raison de la contraction des microfilaments d'actine, se concentrant autour de la circonférence de la cellule (anneau contractile), une constriction cellulaire commence à se former, qui, en s'approfondissant, conduira à la cytotomie dans la phase suivante.
La télophase est la dernière étape de la mitose, au cours de laquelle les noyaux des cellules filles sont reconstruits et leur division est achevée. Autour des chromosomes condensés des cellules filles des vésicules membranaires (selon d'autres sources, d'EPS), le caryolemme est restauré, auquel la lame émergente est associée, les nucléoles réapparaissent, qui sont formés à partir de sections des chromosomes correspondants. Les noyaux cellulaires augmentent progressivement et les chromosomes déspiralisent et disparaissent progressivement, remplacés par le motif chromatinien du noyau interphase. Dans le même temps, la constriction cellulaire s'approfondit et les cellules restent reliées pendant un certain temps par un pont cytoplasmique rétréci contenant un faisceau de microtubules (corps moyen). Une ligature supplémentaire du cytoplasme se termine par la formation de deux cellules filles. En télophase, la distribution des organites entre les cellules filles se produit; L'uniformité de ce processus est facilitée par le fait que certains organites sont assez nombreux (par exemple, les mitochondries), tandis que d'autres (comme l'EPS et le complexe de Golgi) se décomposent en petits fragments et vésicules lors de la mitose.
Les mitoses atypiques se produisent lorsque l'appareil mitotique est endommagé et se caractérisent par une répartition inégale du matériel génétique entre les cellules - aneuploïdie (du grec an - not, eu - correct, ploon - add); dans de nombreux cas, la cytotomie est absente, entraînant la formation de cellules géantes. Les mitoses atypiques sont caractéristiques des tumeurs malignes et des tissus irradiés. Plus leur fréquence est élevée et plus le degré d'aneuploïdie est élevé, plus la tumeur est maligne. La violation de la division cellulaire mitotique normale peut être causée par des anomalies chromosomiques, appelées aberrations chromosomiques (du latin Aberratio - déviation). Les variantes des aberrations chromosomiques sont l'adhésion des chromosomes, leur rupture en fragments, la perte d'un site, l'échange de fragments, le doublement de sections individuelles de chromosomes, etc. Les aberrations chromosomiques peuvent survenir spontanément, mais le plus souvent elles se développent en raison de la action des mutagènes et des rayonnements ionisants sur les cellules.
Caryotypage - une étude diagnostique pour évaluer le caryotype (ensemble de chromosomes) est réalisée en examinant les chromosomes dans la plaque métaphasique. Pour le caryotypage, on obtient une culture cellulaire dans laquelle on introduit de la colchicine, une substance qui bloque la formation du fuseau mitotique. Les chromosomes sont extraits de ces cellules, qui sont ensuite colorées et identifiées. Le caryotype humain normal est représenté par 46 chromosomes - 22 paires d'autosomes et deux chromosomes sexuels (XY chez l'homme et XX chez la femme). Le caryotype peut diagnostiquer un certain nombre de maladies associées à anomalies chromosomiques, en particulier, le syndrome de Down (trisomie du 21e chromosome), Edwards (trisomie du 18e chromosome), Patau (trisomie du 13e chromosome), ainsi qu'un certain nombre de syndromes associés à des anomalies des chromosomes sexuels - le syndrome de Klinefelter ( génotype - XXY), Turner (génotype - XO) et autres.
On suppose que le processus mitotique complexe des organismes supérieurs s'est développé progressivement à partir des mécanismes de division procaryote. Cette hypothèse est étayée par le fait que les procaryotes sont apparus environ un milliard d'années plus tôt que les premiers eucaryotes. De plus, des protéines similaires sont impliquées dans la mitose eucaryote et la fission binaire procaryote.
Des étapes intermédiaires possibles entre la fission binaire et la mitose peuvent être tracées chez les eucaryotes unicellulaires, dans lesquels la membrane nucléaire n'est pas détruite lors de la division. Chez la plupart des autres eucaryotes, y compris les plantes et les animaux, le fuseau de fission se forme à l'extérieur du noyau et la membrane nucléaire est détruite pendant la mitose. Bien que la mitose chez les eucaryotes unicellulaires ne soit pas encore bien comprise, on peut supposer qu'elle provient d'une fission binaire et a finalement atteint le niveau de complexité qui existe dans les organismes multicellulaires.
Chez de nombreux eucaryotes protozoaires, la mitose est également restée un processus associé à la membrane, mais maintenant ce n'est plus du plasma, mais du nucléaire.
Les principaux mécanismes de régulation de la mitose sont les processus de phosphorylation et de protéolyse.
Les réactions de phosphorylation et de déphosphorylation réversibles permettent des événements mitotiques réversibles tels que l'assemblage/la désintégration du fuseau ou la désintégration/la réparation de l'enveloppe nucléaire. La protéolyse sous-tend les événements irréversibles de la mitose, tels que la séparation des chromatides soeurs en anaphase ou la destruction des cyclines mitotiques dans les derniers stades de la mitose.
La division de toutes les cellules eucaryotes est associée à la formation d'un appareil spécial pour la division cellulaire.
Un rôle actif dans la division cellulaire mitotique est souvent attribué aux structures du cytosquelette. Le fuseau mitotique bipolaire, composé de microtubules et de protéines associées, est universel pour les cellules animales et végétales. Le fuseau de division assure une répartition strictement identique des chromosomes entre les pôles de division, dans la région desquels se forment les noyaux des cellules filles dans la télophase.
Le processus de mitose assure une distribution strictement uniforme des chromosomes entre deux noyaux filles, de sorte que dans un organisme multicellulaire, toutes les cellules ont exactement les mêmes ensembles de chromosomes (en nombre et en caractères).
Les chromosomes contiennent des informations génétiques codées dans l'ADN et, par conséquent, un processus mitotique régulier et ordonné assure également le transfert complet de toutes les informations vers chacun des noyaux filles. de ce fait, chaque cellule possède toute l'information génétique nécessaire au développement de toutes les caractéristiques de l'organisme. À cet égard, il devient clair pourquoi une cellule prélevée sur une plante adulte entièrement différenciée peut, dans des conditions appropriées, se développer en une plante entière. Nous avons décrit la mitose dans une cellule diploïde, mais ce processus se déroule de manière similaire dans les cellules haploïdes, par exemple dans les cellules de la génération gamétophyte des plantes.
Le chiot grandit et grossit grâce à la division des cellules somatiques, appelées mitose. La mitose est une division indirecte d'une cellule somatique lorsque des changements complexes se produisent dans son noyau et son cytoplasme. Après la fécondation (ovogamie) de l'ovule par le spermatozoïde (fusion ou copulation de gamètes), une zygote(oocyste) - un nouvel organisme constitué d'une seule cellule. Le processus de croissance et de développement d'un nouvel organisme commence à partir du moment de la première division mitotique de cette cellule (maternelle), lorsque deux cellules filles (plus précisément sœurs), tout à fait similaires, en découlent et se poursuivent jusqu'à la mort .
Fig.1 Structure cellulaire
Au cours de la mitose, il se produit :
1- doublement de la substance des chromosomes ;
2- modification de l'état physique et de l'organisation chimique des chromosomes ;
3- divergence des chromosomes frères aux pôles de la cellule ;
4- division ultérieure du cytoplasme et restauration complète de deux noyaux dans de nouvelles cellules.
En mitose, le cycle de vie des gènes nucléaires est défini : duplication, distribution et fonctionnement. La période entre les divisions cellulaires est appelée interphase, au cours de laquelle s'y déroulent des processus de vie actifs et la préparation de la division suivante. L'ensemble du cycle des changements qui se produisent dans la cellule
d'une division à l'autre s'appelle cycle mitotique. Ce dernier se compose de deux périodes principales - l'interphase et la mitose elle-même.
À la suite de la mitose, deux cellules avec des chromosomes identiques sont formées à partir d'une cellule. Ainsi, la mitose assure la continuité et la constance du nombre et de l'ensemble, c'est-à-dire la spécificité qualitative des chromosomes dans les générations successives de cellules en division (voir Fig. 2).
En interphase, la période entre deux divisions cellulaires successives, la réplication (autoduplication ou auto-doublement) de l'ADN se produit dans le noyau, et donc le nombre de chromosomes dans la cellule (la formation de chromatides sœurs maintenues ensemble par le centromère, c'est-à-dire , l'organe qui remplit la fonction de centre mécanique du chromosome), ainsi que la déspiralisation de ce dernier.
En métaphase, ou phase centrale de la division nucléaire, un chromosome composé de deux chromatides se transforme en deux chromosomes filles.
Riz. 2. Mitose
 1 - trois paires de chromosomes ; 2 - réplication des chromosomes avec formation de chromatides soeurs connectées dans la région du centromère; 3 - aux pôles de la membrane nucléaire, les centrosomes sont visibles avec des rayons astérisques allant vers la région centromérique des chromatides, alignés le long de l'équateur afin de séparer les centromères des chromatides sœurs et de les séparer en pôles différents ; 4 - déspiralisation des chromosomes, restauration de la membrane nucléaire et formation d'un septum cellulaire avec formation de deux cellules filles identiques à la mère avec exactement le même nombre de chromosomes qu'en elle
1 - trois paires de chromosomes ; 2 - réplication des chromosomes avec formation de chromatides soeurs connectées dans la région du centromère; 3 - aux pôles de la membrane nucléaire, les centrosomes sont visibles avec des rayons astérisques allant vers la région centromérique des chromatides, alignés le long de l'équateur afin de séparer les centromères des chromatides sœurs et de les séparer en pôles différents ; 4 - déspiralisation des chromosomes, restauration de la membrane nucléaire et formation d'un septum cellulaire avec formation de deux cellules filles identiques à la mère avec exactement le même nombre de chromosomes qu'en elle
En anaphase, la division et la divergence des chromosomes filles aux pôles de la cellule se produisent, c'est-à-dire la restauration de leur nombre approprié. Dans la télophase, l'étape finale de la division cellulaire, les chromosomes prennent la même forme qu'avant le début de la division et la quantité d'ADN dans chaque noyau fille est réduite de moitié par rapport aux étapes précédentes. Ainsi, les deux cellules filles contiennent la même quantité de cytoplasme et des ensembles identiques de chromosomes et sont prêtes à subir une mitose.
Toutes les cellules somatiques du corps ne se divisent pas constamment. Au cours du processus de développement embryonnaire, la différenciation des organes et des tissus se produit, se développant selon leur propre chemin spécifique, génétiquement prédéterminé. Par conséquent, certaines cellules se transforment en cellules cérébrales, d'autres en cellules sanguines, etc. De plus, certaines d'entre elles se divisent constamment, tandis que d'autres seulement à un certain stade de développement ou, si nécessaire, sont responsables, par exemple, de la régénération
processus (réparateurs).