Tension de la paroi cellulaire chez les plantes. Paroi cellulaire
Lire aussi
A l’aube du développement de la vie sur Terre, toutes les formes cellulaires étaient représentées par des bactéries. Ils absorbaient les substances organiques dissoutes dans l’océan primordial à travers la surface du corps.
Au fil du temps, certaines bactéries se sont adaptées pour produire des substances organiques à partir de substances inorganiques. Pour ce faire, ils ont utilisé l’énergie du soleil. Le premier est apparu système écologique, dans lequel ces organismes étaient producteurs. En conséquence, l’oxygène libéré par ces organismes est apparu dans l’atmosphère terrestre. Avec son aide, vous pouvez obtenir beaucoup plus d'énergie du même aliment et utiliser l'énergie supplémentaire pour compliquer la structure du corps : diviser le corps en parties.
Un des réalisations importantes vie - séparation du noyau et du cytoplasme. Le noyau contient des informations héréditaires. Une membrane spéciale autour du noyau permettait de se protéger contre les dommages accidentels. Selon les besoins, le cytoplasme reçoit des commandes du noyau qui dirigent la vie et le développement de la cellule.
Les organismes dans lesquels le noyau est séparé du cytoplasme ont formé le superrègne nucléaire (ceux-ci comprennent les plantes, les champignons et les animaux).
Ainsi, la cellule - base de l'organisation des plantes et des animaux - est née et s'est développée au cours de l'évolution biologique.
Même à l'œil nu, ou mieux encore à la loupe, on peut constater que la chair d'une pastèque mûre est constituée de très petits grains, ou grains. Ce sont des cellules – les plus petits « éléments constitutifs » qui constituent le corps de tous les organismes vivants, y compris les plantes.
La vie d'une plante s'effectue grâce à l'activité combinée de ses cellules, créant un tout unique. Avec la multicellularité des parties végétales, il existe une différenciation physiologique de leurs fonctions, une spécialisation de diverses cellules en fonction de leur localisation dans le corps végétal.
Une cellule végétale diffère d’une cellule animale en ce qu’elle possède une membrane dense qui recouvre le contenu interne de tous les côtés. La cellule n'est pas plate (comme on le représente habituellement), elle ressemble très probablement à une très petite bulle remplie de contenu muqueux.
Structure et fonctions d'une cellule végétale
Considérons une cellule comme une unité structurelle et fonctionnelle d'un organisme. L’extérieur de la cellule est recouvert d’une paroi cellulaire dense dans laquelle se trouvent des sections plus minces appelées pores. En dessous se trouve un film très fin – une membrane recouvrant le contenu de la cellule – le cytoplasme. Dans le cytoplasme se trouvent des cavités - des vacuoles remplies de jus cellulaire. Au centre ou près du centre de la cellule paroi cellulaire il y a un corps dense - un noyau avec un nucléole. Le noyau est séparé du cytoplasme par l'enveloppe nucléaire. De petits corps appelés plastes sont répartis dans tout le cytoplasme.
Structure cellule de plante
Structure et fonctions des organites des cellules végétales
| Organoïde | Dessin | Description | Fonction | Particularités |
Paroi cellulaire ou membrane plasmique | Incolore, transparent et très résistant | Fait passer des substances dans et hors de la cellule. | La membrane cellulaire est semi-perméable |
|
Cytoplasme | Substance visqueuse épaisse | Toutes les autres parties de la cellule s'y trouvent | Est en mouvement constant |
|
Noyau (partie importante de la cellule) | Rond ou ovale | Assure le transfert des propriétés héréditaires aux cellules filles lors de la division | Partie centrale de la cellule |
|
De forme sphérique ou irrégulière | Participe à la synthèse des protéines | |||
 | Réservoir séparé du cytoplasme par une membrane. Contient de la sève cellulaire | Les nutriments de rechange et les déchets dont la cellule n’a pas besoin s’accumulent. | Au fur et à mesure que la cellule grandit, les petites vacuoles fusionnent en une seule grande vacuole (centrale). |
|
Plastides | Chloroplastes | Ils utilisent l'énergie lumineuse du soleil et créent de la matière organique à partir de matière inorganique. | La forme des disques délimités du cytoplasme par une double membrane |
|
Chromoplastes | Formé à la suite de l’accumulation de caroténoïdes | Jaune, orange ou marron |
||
 | Leucoplastes | Plastides incolores | ||
Enveloppe nucléaire | Se compose de deux membranes (externe et interne) avec des pores | Sépare le noyau du cytoplasme | Permet les échanges entre le noyau et le cytoplasme |
La partie vivante d'une cellule est un système ordonné et structuré de biopolymères et de structures membranaires internes impliqués dans un ensemble de processus métaboliques et énergétiques qui maintiennent et reproduisent l'ensemble du système dans son ensemble.
Une caractéristique importante est que la cellule ne possède pas de membranes ouvertes aux extrémités libres. Les membranes cellulaires limitent toujours les cavités ou zones, les fermant de tous côtés.

Schéma généralisé moderne d'une cellule végétale
Plasmalemme(membrane cellulaire externe) est un film ultramicroscopique de 7,5 nm d'épaisseur, constitué de protéines, de phospholipides et d'eau. Il s'agit d'un film très élastique, bien mouillé par l'eau et qui rétablit rapidement son intégrité après un dommage. Il a une structure universelle, c'est-à-dire typique de toutes les membranes biologiques. Dans les cellules végétales, à l’extérieur de la membrane cellulaire se trouve une paroi cellulaire solide qui crée un support externe et maintient la forme de la cellule. Il est constitué de fibres (cellulose), un polysaccharide insoluble dans l'eau.
Plasmodesmes les cellules végétales, sont des tubules submicroscopiques qui pénètrent dans les membranes et sont tapissés d'une membrane plasmique, qui passe ainsi d'une cellule à l'autre sans interruption. Avec leur aide, une circulation intercellulaire de solutions contenant des nutriments organiques se produit. Ils transmettent également des biopotentiels et d’autres informations.
Porami appelées ouvertures dans la membrane secondaire, où les cellules ne sont séparées que par la membrane primaire et la lame médiane. Les zones de la membrane primaire et de la plaque médiane séparant les pores adjacents des cellules adjacentes sont appelées membrane poreuse ou film de fermeture du pore. Le film de fermeture du pore est percé de tubules plasmodesmiques, mais aucun trou traversant n'est généralement formé dans les pores. Les pores facilitent le transport de l'eau et des solutés d'une cellule à l'autre. Des pores se forment dans les parois des cellules voisines, généralement les unes en face des autres.
Membrane cellulaire a une coque bien définie et relativement épaisse de nature polysaccharidique. L'enveloppe d'une cellule végétale est le produit de l'activité du cytoplasme. L'appareil de Golgi et le réticulum endoplasmique participent activement à sa formation.

Structure de la membrane cellulaire
La base du cytoplasme est sa matrice, ou hyaloplasme, un système colloïdal complexe, incolore et optiquement transparent, capable de transitions réversibles du sol au gel. Le rôle le plus important du hyaloplasme est d'unir toutes les structures cellulaires en système unifié et assurer l'interaction entre eux dans les processus du métabolisme cellulaire.
Hyaloplasme(ou matrice cytoplasmique) est environnement interne cellules. Constitué d'eau et de divers biopolymères (protéines, acides nucléiques, polysaccharides, lipides), dont la majeure partie sont des protéines de spécificité chimique et fonctionnelle variable. Le hyaloplasme contient également des acides aminés, des monosaccharides, des nucléotides et d'autres substances de faible poids moléculaire.
Les biopolymères forment un milieu colloïdal avec l'eau qui, selon les conditions, peut être dense (sous forme de gel) ou plus liquide (sous forme de sol), à la fois dans tout le cytoplasme et dans ses sections individuelles. Dans le hyaloplasme, divers organites et inclusions sont localisés et interagissent entre eux et avec l'environnement hyaloplasmique. De plus, leur localisation est le plus souvent spécifique à certains types de cellules. A travers la membrane bilipide, le hyaloplasme interagit avec l'environnement extracellulaire. Le hyaloplasme est donc un milieu dynamique et joue rôle important dans le fonctionnement des organites individuels et dans la vie des cellules dans leur ensemble.
Formations cytoplasmiques - organites
Les organites (organites) sont des composants structurels du cytoplasme. Ils ont une certaine forme et taille et constituent des structures cytoplasmiques obligatoires de la cellule. S’ils sont absents ou endommagés, la cellule perd généralement sa capacité à continuer d’exister. De nombreux organites sont capables de se diviser et de s’auto-reproduire. Leurs tailles sont si petites qu’elles ne peuvent être vues qu’au microscope électronique.
Cœur
Le noyau est l’organite le plus important et généralement le plus grand de la cellule. Il a été étudié pour la première fois en détail par Robert Brown en 1831. Le noyau assure les fonctions métaboliques et génétiques les plus importantes de la cellule. Sa forme est assez variable : elle peut être sphérique, ovale, lobée ou en forme de lentille.
Le noyau joue un rôle important dans la vie de la cellule. Une cellule dont le noyau a été retiré ne sécrète plus de membrane et cesse de croître et de synthétiser des substances. Les produits de décomposition et de destruction s'y intensifient, ce qui entraîne sa mort rapide. La formation d'un nouveau noyau à partir du cytoplasme ne se produit pas. Les nouveaux noyaux se forment uniquement en divisant ou en écrasant les anciens.
Le contenu interne du noyau est la caryolymphe (suc nucléaire), qui remplit l'espace entre les structures du noyau. Il contient un ou plusieurs nucléoles, ainsi qu'un nombre important de molécules d'ADN liées à des protéines spécifiques - les histones.

Structure de base
Nucléole
Le nucléole, comme le cytoplasme, contient majoritairement de l'ARN et des protéines spécifiques. Sa fonction la plus importante est de former des ribosomes, qui réalisent la synthèse des protéines dans la cellule.
Appareil de Golgi
L'appareil de Golgi est un organite universellement distribué dans tous les types de cellules eucaryotes. Il s'agit d'un système à plusieurs niveaux de sacs membranaires plats qui s'épaississent le long de la périphérie et forment des processus vésiculaires. Elle est le plus souvent située à proximité du noyau.

Appareil de Golgi
L'appareil de Golgi comprend nécessairement un système de petites vésicules (vésicules), qui se détachent de citernes épaissies (disques) et sont situées le long de la périphérie de cette structure. Ces vésicules jouent le rôle de cellules intracellulaires Système de transport des granules sectoriels spécifiques peuvent servir de source de lysosomes cellulaires.
Les fonctions de l'appareil de Golgi consistent également en l'accumulation, la séparation et la libération à l'extérieur de la cellule à l'aide de vésicules de produits de synthèse intracellulaire, de produits de désintégration et de substances toxiques. Les produits de l'activité synthétique de la cellule, ainsi que diverses substances entrant dans la cellule depuis l'environnement par les canaux du réticulum endoplasmique, sont transportés vers l'appareil de Golgi, s'accumulent dans cet organite, puis sous forme de gouttelettes ou de grains pénètrent dans le cytoplasme et sont soit utilisés par la cellule elle-même, soit excrétés à l'extérieur. Dans les cellules végétales, l'appareil de Golgi contient des enzymes pour la synthèse des polysaccharides et le matériau polysaccharidique lui-même, qui est utilisé pour construire la paroi cellulaire. On pense qu’il participe à la formation des vacuoles. L'appareil de Golgi doit son nom au scientifique italien Camillo Golgi, qui l'a découvert pour la première fois en 1897.
Lysosomes
Les lysosomes sont de petites vésicules délimitées par une membrane dont la fonction principale est d'effectuer la digestion intracellulaire. L'utilisation de l'appareil lysosomal se produit lors de la germination d'une graine végétale (hydrolyse de la réserve nutriments).
Structure d'un lysosome
Microtubules
Les microtubules sont des structures membraneuses supramoléculaires constituées de globules protéiques disposés en spirale ou en rangées droites. Les microtubules remplissent une fonction principalement mécanique (motrice), assurant la mobilité et la contractilité des organites cellulaires. Situés dans le cytoplasme, ils donnent à la cellule une certaine forme et assurent la stabilité de la disposition spatiale des organites. Les microtubules favorisent le mouvement des organites vers des endroits déterminés Besoins psycologiques cellules. Un nombre important de ces structures sont localisées dans le plasmalemme, à proximité de la membrane cellulaire, où elles participent à la formation et à l'orientation des microfibrilles de cellulose des parois cellulaires végétales.

Structure des microtubules
Vacuole
La vacuole est le composant le plus important des cellules végétales. C'est une sorte de cavité (réservoir) dans la masse du cytoplasme, remplie d'une solution aqueuse des sels minéraux, acides aminés, acides organiques, pigments, glucides et séparés du cytoplasme par une membrane vacuolaire - le tonoplaste.
Le cytoplasme remplit toute la cavité interne uniquement dans les cellules végétales les plus jeunes. Au fur et à mesure que la cellule grandit, la disposition spatiale de la masse initialement continue de cytoplasme change de manière significative : de petites vacuoles remplies de sève cellulaire apparaissent et la masse entière devient spongieuse. Avec la poursuite de la croissance cellulaire, les vacuoles individuelles fusionnent, poussant les couches de cytoplasme vers la périphérie, de sorte que la cellule formée contient généralement une grande vacuole et que le cytoplasme avec tous les organites est situé près de la membrane.
Les composés organiques et minéraux solubles dans l'eau des vacuoles déterminent les propriétés osmotiques correspondantes des cellules vivantes. Cette solution d'une certaine concentration est une sorte de pompe osmotique permettant une pénétration contrôlée dans la cellule et la libération de l'eau, des ions et des molécules métabolites.
En combinaison avec la couche cytoplasmique et ses membranes, caractérisées par des propriétés semi-perméables, la vacuole forme un système osmotique efficace. Les indicateurs de cellules végétales vivantes tels que le potentiel osmotique, la force d'aspiration et la pression de turgescence sont déterminés de manière osmotique.

Structure de la vacuole
Plastides
Les plastes sont les plus grands organites cytoplasmiques (après le noyau), inhérents uniquement aux cellules des organismes végétaux. On ne les trouve pas uniquement dans les champignons. Les plastes jouent un rôle important dans le métabolisme. Ils sont séparés du cytoplasme par une double membrane et certains types possèdent un système de membranes internes bien développé et ordonné. Tous les plastes sont de la même origine.
Chloroplastes- les plastes les plus courants et les plus importants sur le plan fonctionnel des organismes photoautotrophes qui effectuent des processus photosynthétiques conduisant finalement à la formation matière organique et libération d'oxygène libre. Les chloroplastes des plantes supérieures ont un complexe structure interne.

Structure chloroplastique
Les tailles des chloroplastes dans différentes plantes ne sont pas les mêmes, mais leur diamètre moyen est de 4 à 6 microns. Les chloroplastes sont capables de se déplacer sous l'influence du mouvement du cytoplasme. De plus, sous l'influence de l'éclairage, on observe un mouvement actif des chloroplastes de type amiboïde vers la source lumineuse.
La chlorophylle est la substance principale des chloroplastes. Grâce à la chlorophylle plantes vertes capable d'utiliser l'énergie lumineuse.
Leucoplastes(plastes incolores) sont des corps cytoplasmiques clairement définis. Leurs tailles sont légèrement inférieures à celles des chloroplastes. Leur forme est également plus uniforme, se rapprochant de la sphérique.

Structure leucoplastique
Trouvé dans les cellules épidermiques, les tubercules et les rhizomes. Lorsqu'ils sont illuminés, ils se transforment très rapidement en chloroplastes avec une modification correspondante de la structure interne. Les leucoplastes contiennent des enzymes à l'aide desquelles l'amidon est synthétisé à partir de l'excès de glucose formé lors de la photosynthèse, dont l'essentiel se dépose dans les tissus ou organes de stockage (tubercules, rhizomes, graines) sous forme de grains d'amidon. Dans certaines plantes, les graisses se déposent dans les leucoplastes. La fonction de réserve des leucoplastes se manifeste parfois par la formation de protéines de réserve sous forme de cristaux ou d'inclusions amorphes.
Chromoplastes dans la plupart des cas, ce sont des dérivés de chloroplastes, parfois de leucoplastes.

Structure chromoplastique
La maturation des cynorrhodons, des poivrons et des tomates s'accompagne de la transformation des chloro- ou leucoplastes des cellules pulpaires en plastes caratinoïdes. Ces derniers contiennent principalement des pigments plastidiques jaunes - des caroténoïdes, qui, à maturité, y sont intensément synthétisés, formant des gouttelettes lipidiques colorées, des globules solides ou des cristaux. Dans ce cas, la chlorophylle est détruite.
Mitochondries
Les mitochondries sont des organites caractéristiques de la plupart des cellules végétales. Ils ont une forme variable de bâtons, de grains et de fils. Découvert en 1894 par R. Altman à l'aide d'un microscope optique, la structure interne a ensuite été étudiée à l'aide d'un microscope électronique.
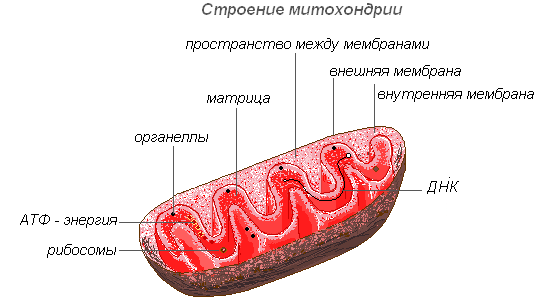
La structure des mitochondries
Les mitochondries ont une structure à double membrane. La membrane externe est lisse, la membrane interne forme diverses formes les excroissances sont des tubes dans les cellules végétales. L'espace à l'intérieur de la mitochondrie est rempli d'un contenu semi-liquide (matrice), qui comprend des enzymes, des protéines, des lipides, des sels de calcium et de magnésium, des vitamines, ainsi que de l'ARN, de l'ADN et des ribosomes. Le complexe enzymatique des mitochondries accélère le mécanisme complexe et interconnecté des réactions biochimiques qui aboutissent à la formation d'ATP. Dans ces organites, les cellules reçoivent de l'énergie - l'énergie des liaisons chimiques des nutriments est convertie en liaisons ATP à haute énergie au cours du processus de respiration cellulaire. C'est dans les mitochondries que se produit la dégradation enzymatique des glucides, des acides gras et des acides aminés avec la libération d'énergie et sa conversion ultérieure en énergie ATP. L'énergie accumulée est dépensée en processus de croissance, en nouvelles synthèses, etc. Les mitochondries se multiplient par division et vivent environ 10 jours, après quoi elles sont détruites.
Réticulum endoplasmique
Le réticulum endoplasmique est un réseau de canaux, tubes, vésicules et citernes situés à l'intérieur du cytoplasme. Découvert en 1945 par le scientifique anglais K. Porter, il s'agit d'un système de membranes à structure ultramicroscopique.

Structure du réticulum endoplasmique
L'ensemble du réseau est combiné en un seul tout avec le réseau externe membrane cellulaire coque nucléaire. Il existe des RE lisses et rugueux, qui portent des ribosomes. Sur les membranes du RE lisse se trouvent des systèmes enzymatiques impliqués dans le métabolisme des graisses et des glucides. Ce type de membrane prédomine dans les cellules graines riches en substances de stockage (les protéines, les glucides, les huiles sont attachées à la membrane granulaire du RE, et lors de la synthèse d'une molécule protéique, la chaîne polypeptidique avec les ribosomes est immergée dans le canal ER. Les fonctions du réticulum endoplasmique sont très diverses : transport de substances aussi bien au sein de la cellule qu'entre cellules voisines ; division d'une cellule en sections distinctes dans lesquelles divers processus physiologiques et réactions chimiques se déroulent simultanément.
Ribosomes
Les ribosomes sont des organites cellulaires non membranaires. Chaque ribosome est constitué de deux particules de taille différente et pouvant être divisées en deux fragments, qui conservent la capacité de synthétiser des protéines après s'être combinés en un ribosome entier.
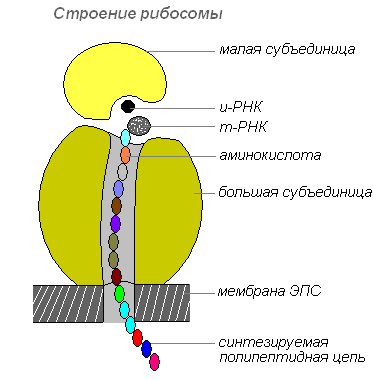
Structure des ribosomes
Les ribosomes sont synthétisés dans le noyau, puis le quittent et se déplacent vers le cytoplasme, où ils sont attachés à la surface externe des membranes du réticulum endoplasmique ou sont localisés librement. Selon le type de protéine synthétisée, les ribosomes peuvent fonctionner seuls ou être combinés en complexes - les polyribosomes.
La paroi cellulaire est un dérivé du protoplaste, c'est-à-dire se forme au cours de son activité vitale (Fig. 61). Il donne à la cellule une certaine forme, protège le protoplaste et, en résistant à la pression intracellulaire, prévient la rupture cellulaire. Remplissant les fonctions du squelette interne d'une plante, les parois cellulaires confèrent à ses organes la résistance mécanique nécessaire.
Les parois cellulaires transmettent bien la lumière du soleil et l'eau et les minéraux qui y sont dissous se déplacent facilement à travers elles. Entre les murs des cellules voisines il y a plaque intermédiaire - couche de pectine qui, étant en fait une substance intercellulaire, maintient ensemble les parois des cellules voisines. Dans les endroits où les parois cellulaires des cellules voisines ne se ferment pas, remplies d'eau espaces intercellulaires. Le processus de destruction de la substance intercellulaire, à la suite duquel les parois des cellules voisines sont séparées, est appelé macération. Une macération naturelle peut être observée
Riz. 61.
UN- schéma de la structure de la paroi cellulaire ; B- schéma de la participation de l'appareil de Golgi à la construction de la paroi cellulaire ; DANS- structure détaillée de la paroi cellulaire : 1 - plaque médiane ; 2 - C'est l'heure; 3 - mur secondaire;
- 4 - mur primaire; 5 - dictyosome ; 6 - Vésicules de Golgi ;
- 7- plasmalemme ; 8- paroi cellulaire; 9- macrofibrille;
- 10- microfibrilles ; 11 - micelle ; 12 - molécule de cellulose;
- 13 - structure d'un fragment de molécule de cellulose
dans les fruits trop mûrs de pomme, de sorbier, de melon, etc. Une macération artificielle est réalisée, par exemple, lors du trempage des tiges de lin pour en séparer les fibres libériennes ; Cela se produit également lors du traitement thermique des fruits.
La paroi cellulaire contient des polysaccharides : pectines, hémicellulose et cellulose. De très longues molécules de cellulose sont disposées parallèlement les unes aux autres (40 à 60 chacune), formant micelles. Les micelles sont collectées en paquets - des microfibrilles, représentant l’unité structurelle de base de la cellulose. Les microfibrilles, à leur tour, sont combinées en macrofibrilles- des fibres très fines de longueur incertaine. Les macrofibrilles de cellulose sont immergées dans un milieu très arrosé matrice, constitué de pectines, d'hémicelluloses et de quelques autres substances. La résistance de la paroi cellulaire est assurée par des microfibrilles de cellulose élastiques, dont la résistance à la traction est proche de celle de l'acier. La force et l’élasticité de la paroi cellulaire sont à la base de sa capacité à s’étirer de manière réversible. Grâce aux pectines et à l'hémicellulose, la paroi cellulaire est très perméable à l'eau - l'eau et les substances qui y sont dissoutes se déplacent facilement d'une cellule à l'autre.
La paroi cellulaire est adjacente à l’extérieur du plasmalemme, qui participe activement à sa croissance. Des molécules de pectines, d'hémicellulose, de cellulose et d'autres substances sont synthétisées et accumulées dans les citernes dictyosomes de l'appareil de Golgi. Les vésicules de Golgi les délivrent à la périphérie du protoplaste - au plasmalemme. Au point de contact entre la vésicule et le plasmalemme, ce dernier se dissout et le contenu de la vésicule, une fois à l'extérieur du plasmalemme, sert à construire la paroi cellulaire. La membrane vésiculaire restaure non seulement l'intégrité du plasmalemme, mais assure également sa croissance en surface. La croissance de la paroi cellulaire est réalisée grâce à l'activité enzymatique du plasmalemme.
Les parois des cellules en division et en croissance sont appelées primaire. Ils contiennent beaucoup d'eau (60-90%), leur matière sèche est dominée par les pectines et l'hémicellulose - la cellulose n'y contient pas plus de 30%. Lorsqu'une cellule se divise au cours de la télophase de la mitose, la cellule mère est divisée en deux cellules filles suite à la formation d'un septum dans son plan équatorial - plaque médiane. Des deux côtés de la lame médiane, chacune des deux cellules filles commence à créer sa propre paroi cellulaire primaire. La croissance de la plaque médiane et des parois primaires des deux cellules filles se produit dans une direction centrifuge - du centre de la cellule mère vers sa périphérie. La plaque médiane est très fine et constituée de pectine.
La nouvelle cellule formée à la suite de la division commence à croître et son volume peut augmenter de 100 fois ou plus. La croissance cellulaire s'effectue principalement par entorses en raison de l'absorption d'eau et de l'augmentation du volume des vacuoles. La pression interne qui en résulte étire la paroi primaire, dans laquelle s'incrustent facilement les micelles de cellulose, les pectines et les hémicelluloses. Méthode de croissance de la paroi cellulaire par mise en œuvre le matériau de construction entre les structures existantes est appelé intussusception.
Dans la paroi cellulaire primaire, il y a initialement des zones plus fines où les fibrilles de cellulose sont disposées de manière plus lâche - champs de pores primaires. Les champs de pores primaires des parois de deux cellules voisines coïncident généralement. Ici, les tubules du réticulum endoplasmique passent d'une cellule à l'autre - plasmodesmes. Les chemins par lesquels les plasmodesmes passent d'une cellule à une autre sont appelés tubules plasmodesminaux. Grâce à ces tubules, les hyaloplasmes des cellules voisines sont reliés entre eux. Les plasmodesmes assurent le transport intercellulaire de substances (hormones, acides aminés, ATP, sucres, etc.). Les protoplastes des cellules du corps réunis en un seul tout à l'aide de plasmodesmes sont appelés symplastes. Le transport de substances le long des plasmodesmes est appelé symplastique.(La totalité des parois cellulaires, de la lame médiane et des espaces intercellulaires est appelée apoplaste, marche dessus apoplastique transport de substances.)
Une fois la croissance cellulaire terminée, sa paroi primaire peut rester mince (dans les cellules du tissu éducatif) ou commencer à s'épaissir (dans les cellules du tissu permanent). La croissance de l’épaisseur de la paroi cellulaire est appelée épaississement secondaire. De ce fait, il se dépose sur la surface interne de la paroi primaire. mur secondaire, qui grandit de apposition- application de micelles de cellulose sur un mur existant. Dans ce cas, les couches les plus jeunes de la paroi cellulaire secondaire sont situées à côté du plasmalemme. La paroi cellulaire secondaire remplit principalement des fonctions de support et mécaniques. Il contient de manière significative moins d'eau que dans le primaire, et la cellulose prédomine dans la matière sèche (jusqu'à 50 %). Par exemple, dans les parois secondaires des poils unicellulaires en fibres libériennes de coton et de lin, la teneur en cellulose peut atteindre 95 %.
L'épaississement secondaire de la paroi cellulaire se produit de manière inégale. Des sections de la paroi cellulaire secondaire aux emplacements des champs de pores primaires restent généralement non épaissies. Ces zones non épaissies de la paroi cellulaire sont appelées a l'heure. En règle générale, les pores dans les parois de deux cellules voisines coïncident, formant deux-trois fois. Le canal poreux formé par une paire de pores est obstrué film de fermeture des pores - un septum constitué d'une lame médiane et de deux parois primaires de cellules adjacentes. Le film de fermeture du pore est pénétré par de nombreux plasmodesmes, à travers lesquels passent les plasmodesmes.
Les pores sont distingués simple Et bordé(Fig. 62). Dans les pores simples, le diamètre de la section de leur canal poreux est le même sur toute la longueur, c'est-à-dire il est de forme cylindrique. Les pores simples sont typiques des cellules du parenchyme. Les pores bordés sont caractéristiques des parois des cellules qui conduisent l'eau contenant des minéraux dissous - trachéides et segments vasculaires. Dans de tels pores, leur partie du canal poreux a la forme d'un entonnoir dont le côté large est adjacent au film de fermeture du pore.
Dans les cellules des tissus conducteurs des conifères, le film de fermeture du pore n'est perméable à l'eau que sur les bords, puisque sa partie centrale en forme de disque épaissie et lignifiée est tore - ne laisse pas passer l'eau. Le tore joue le rôle de valve. Si la pression de l'eau dans les cellules voisines n'est pas la même, le film de fermeture est dévié et le tore bloque le mouvement de l'eau à travers le canal poreux.

Riz. 62.
UN- simple; B- bordé ; DANS- semi-tranchant :
1 - film de clôture ; 2 - canal poreux ; 3 - tore
En plus des pores, dans les parois des cellules conductrices d'eau, perforation- trous traversants (segments de vaisseaux, cellules de stockage d'eau de sphaigne).
Modifications de la paroi cellulaire. En fonction des fonctions remplies par la cellule, sa paroi peut être modifiée en raison du dépôt d'éventuelles substances qui s'y trouvent. Ses modifications habituelles sont : la lignification, la subérisation, la cutinisation, la minéralisation et le mucilage.
Lignification de la paroi cellulaire, ou lignification, se produit à la suite du dépôt dans les espaces intermicellaires de lignine - une substance de nature aromatique avec un complexe structure chimique. La résistance et la dureté du mur augmentent, mais son élasticité diminue. Les murs lignifiés sont capables de laisser passer l’eau et l’air. Avec une paroi cellulaire lignifiée, le protoplaste de la cellule peut rester vivant, mais meurt généralement. Dans certaines plantes ligneuses, jusqu'à 30 % de lignine s'accumule dans le bois. La lignine peut également s'accumuler dans les parois cellulaires des pousses de graminées vieillissantes, ce qui réduit considérablement leur valeur alimentaire et détermine le moment de la récolte du foin. Lors du processus d'obtention de pâte de cellulose à partir du bois nécessaire à la production de papier, un déboisement artificiel est effectué. Une lignification naturelle de la paroi cellulaire est possible, mais rare.
Échantillonnage, ou subérinisation, - dépôt d'une substance amorphe persistante ressemblant à de la graisse, la subérine (polymère hydrophobe) dans la paroi cellulaire. Les parois cellulaires subérisées sont imperméables aux gaz et à l'eau, ce qui provoque la mort du protoplaste. Les cellules aux parois subérisées protègent les plantes de manière fiable contre la perte d'eau, les températures extrêmes, les bactéries pathogènes et les champignons.
Cutinisation - dépôt dans les parois cellulaires de cutine (une substance similaire composition chimiqueà la subérine). La cutine se dépose généralement dans les couches superficielles des parois externes des cellules et à leur surface. Sous forme de film - cuticule - il recouvre par exemple la surface des cellules du tissu tégumentaire - l'épiderme.
Minéralisation la destruction de la paroi cellulaire se produit en raison du dépôt de sels de calcium et de silice. Ces substances confèrent au mur dureté et fragilité. Le processus de minéralisation est particulièrement bien exprimé dans les parois cellulaires de l'épiderme des pousses de céréales, de carex et de prêle. Pour cette raison, il est recommandé de tondre les pousses de carex et de graminées avant leur floraison. Plus tard, en raison d'une forte minéralisation, elles deviennent plus grossières, ce qui réduit la qualité du foin.
Vase- transformation de la cellulose et des pectines des parois cellulaires en polysaccharides spéciaux - mucus et gommes, capables de fortement gonfler au contact de l'eau. Le glissement de la paroi est observé dans les cellules de la peau des graines, par exemple chez le coing, le lin, le concombre et le plantain. Le mucilage collant peut favoriser la dispersion des graines (plantain) ; Lorsque les graines germent, le mucilage, absorbant et retenant l’eau, les protège du dessèchement. Dans la coiffe racinaire, le mucus joue le rôle de lubrifiant, facilitant le passage de la racine entre les mottes de terre. Le mucus et les gencives peuvent se former en quantités importantes lorsque les parois cellulaires se dissolvent en raison de leurs dommages. Les cerises et les prunes produisent souvent de la gomme lorsque les branches et les troncs sont blessés. La soi-disant colle de cerise est une gomme qui durcit sous forme de billes, qui recouvre la surface des plaies, des trous de gel, empêchant ainsi l'infection d'y pénétrer. La boue de cette nature est appelée gumose et est considéré comme un phénomène pathologique.
Puisque les parois cellulaires secondaires agissent comme le squelette interne de la plante, donnant la force nécessaire à ses organes (ce qui est particulièrement important pour Plantes terrestres), ils sont souvent capables de s'épaissir de manière significative - localement ou complètement - afin de donner une plus grande résistance aux tissus, et donc à l'organe végétal. L'épaississement de la paroi cellulaire est dû au dépôt de cellulose.
Les fonctions des cellules sont souvent assurées exclusivement par leurs parois, puisque les protoplastes des cellules meurent. Cela concerne les alvéoles de liège,

Riz. 63.
trachéides, segments vasculaires, fibres tissulaires mécaniques. Le bois qui occupe la plupart des énormes troncs d'arbres est, par exemple, principalement constitué de parois cellulaires lignifiées, dont les protoplastes sont morts depuis longtemps.
Les parois cellulaires jouent un rôle important dans nos vies. On en obtient des matières premières textiles (poils de graines de coton, fibres de lin, etc.) et des matières premières pour la production de cordes et cordages (fibres de chanvre, fibres de corde, sisal, etc.). La cellulose extraite des parois cellulaires est utilisée pour fabriquer du papier (bois d’épicéa et de tremble), de la soie acétate, de la viscose, des plastiques, de la cellophane et bien plus encore. Tissu constitué de cellules mortes aux parois subérisées, le liège est utilisé depuis longtemps comme un précieux matériau d'isolation thermique étanche à l'eau et à l'air et est de plus en plus utilisé dans la construction moderne.
La paroi cellulaire végétale est formée avec la participation de la membrane plasmique et constitue une formation multicouche extracellulaire (extracellulaire) qui protège la surface cellulaire et sert de squelette externe de la cellule végétale (Fig. 1). La paroi cellulaire végétale est constituée de deux composants : une matrice (base) de type gel plastique amorphe à haute teneur en eau et un système fibrillaire de soutien. Des substances polymères et des sels supplémentaires, souvent inclus dans la composition des coquilles, leur confèrent de la rigidité et les rendent non mouillables.
Riz. 1. Schéma de la structure de la paroi cellulaire végétale : 0 -- plaque médiane ; 1 -- coque primaire (deux couches de chaque côté de 0) ; 2 -- couches de la coque secondaire ; 3 - coque tertiaire ; PM - membrane plasmique ; B - vacuole ; je suis le noyau
Chimiquement, les principaux composants des coquilles végétales appartiennent aux polysaccharides structuraux. La matrice des coquilles végétales comprend des groupes hétérogènes de polysaccharides qui se dissolvent dans des alcalis concentrés, des hémicelluloses et des substances pectiques. Les hémicelluloses sont des chaînes polymères ramifiées constituées de divers hexoses (glucose, mannose, galactose, etc.), de pentoses (xylose, arabinose) et d'acides uroniques (glucuronique et galacturonique). Ces composants des hémicelluloses se combinent entre eux dans différents rapports quantitatifs et forment diverses combinaisons. Les chaînes de molécules d'hémicellulose ne cristallisent pas et ne forment pas de fibrilles élémentaires. En raison de la présence de groupes polaires d'acides uroniques, ils sont hautement hydratés.
Les substances pectiques constituent un groupe hétérogène qui comprend des polymères ramifiés hautement hydratés qui portent des charges négatives dues à de nombreux résidus d'acide galacturonique. De par les propriétés de ses composants, la matrice est une masse plastique souple renforcée de fibrilles.
Les composants fibreux des parois cellulaires végétales sont généralement constitués de cellulose, un polymère de glucose linéaire et non ramifié. Le poids moléculaire de la cellulose varie de 5,104 à 5,105, ce qui correspond à 300-3000 résidus glucose. De telles molécules linéaires de cellulose peuvent être combinées en faisceaux ou en fibres. Dans la paroi cellulaire, la cellulose forme des fibrilles, constituées de microfibrilles submicroscopiques atteignant 25 nm d'épaisseur, elles-mêmes constituées de nombreuses chaînes parallèles de molécules de cellulose.
Les rapports quantitatifs cellulose/substances matricielles (hémicellulose) peuvent être très différents selon les objets. Plus de 60 % de la masse sèche des coquilles primaires est constituée de leur matrice et environ 30 % est constituée de la substance squelettique - la cellulose. Dans les membranes cellulaires brutes, presque toute l'eau est associée aux hémicelluloses, de sorte que la masse de la substance principale à l'état gonflé atteint 80 % du poids humide de la membrane entière, tandis que la teneur en substances fibreuses est réduite à seulement 12 %. Dans les poils de coton, le composant cellulosique représente 90 % ; Dans le bois, la cellulose représente 50 % des composants de la paroi cellulaire.
Outre la cellulose, l'hémicellulose et les pectines, la composition des parois cellulaires comprend composants supplémentaires, leur conférant des propriétés particulières. Ainsi, l'incrustation (incorporation à l'intérieur) des membranes avec de la lignine (polymère d'alcool coniférylique) conduit à une lignification des parois cellulaires, augmentant leur résistance (Fig. 2). La lignine mélange les substances plastiques de la matrice dans de telles coques et joue le rôle de substance principale à haute résistance. Souvent la matrice est renforcée par des minéraux (SiO2, CaCO3, etc.).

Riz. 2. Incrustation de la membrane cellulaire : a - charpente fibrillaire et matrice interfibrillaire ; b - coque incrustée de lignine avec des résidus matriciels qui ont perdu leur capacité à s'étirer ; c -- incrustation ultérieure de phénols et (ou) de minéraux, conduisant à une augmentation de la dureté de la coque
Diverses substances incrustantes, telles que la cutine et la subérine, peuvent s'accumuler à la surface de la membrane cellulaire, entraînant une subérisation des cellules. Dans les cellules épidermiques, la cire se dépose à la surface des membranes cellulaires, formant une couche imperméable qui empêche la cellule de perdre de l'eau.
En raison de sa structure poreuse et lâche, la paroi cellulaire végétale est largement perméable aux composés de faible poids moléculaire tels que l'eau, les sucres et les ions. Mais les macromolécules pénètrent mal dans les coques cellulosiques : la taille des pores des coques, permettant la libre diffusion des substances, n'est que de 3 à 5 nm.
Des expériences avec des composés marqués ont montré que lorsque la membrane cellulaire se développe, les substances à partir desquelles elle est construite sont libérées sur toute la surface de la cellule. Les substances matricielles amorphes, les hémicelluloses et les pectines sont synthétisées dans les vacuoles de l'appareil de Golgi et libérées par le plasmalemme par exocytose. Les fibrilles de cellulose sont synthétisées par des enzymes spéciales intégrées au plasmalemme.
Les membranes des cellules différenciées et matures sont généralement multicouches ; dans les couches, les fibrilles de cellulose sont orientées différemment et leur nombre peut également fluctuer considérablement. En règle générale, les parois cellulaires primaires, secondaires et tertiaires sont décrites (voir Fig. 1). Afin de comprendre la structure et l’apparence de ces membranes, il est nécessaire de se familiariser avec la façon dont elles se forment après la division cellulaire.
Lorsque les cellules végétales se divisent après une divergence chromosomique, un groupe de petites vésicules membranaires apparaît dans le plan équatorial des cellules, qui commencent à fusionner les unes avec les autres dans la partie centrale des cellules (Fig. 3). Ce processus de fusion de petites vacuoles se produit du centre de la cellule vers la périphérie et se poursuit jusqu'à ce que les vésicules membranaires fusionnent entre elles et avec la membrane plasmique de la surface latérale de la cellule. C'est ainsi que se forme une plaque cellulaire, ou phragmoplaste. Dans sa partie centrale se trouve une substance matricielle amorphe qui remplit les vésicules en fusion. Il a été prouvé que ces vacuoles primaires proviennent des membranes de l'appareil de Golgi. La paroi cellulaire primaire comprend également une petite quantité de protéine (environ 10 %), riche en hydroxyproline et possédant de nombreuses chaînes oligosaccharidiques courtes, ce qui définit cette protéine comme une glycoprotéine. Le long de la périphérie de la plaque cellulaire, lorsqu'elle est observée en lumière polarisée, une biréfringence notable est détectée, provoquée par le fait que des fibrilles de cellulose orientées se trouvent à cet endroit. Ainsi, la paroi cellulaire primaire en croissance se compose déjà de trois couches : la couche centrale - la plaque médiane, constituée uniquement d'une matrice amorphe, et deux périphériques - la membrane primaire contenant de l'hémicellulose et des fibrilles de cellulose. Si la lame médiane est le produit de l'activité de la cellule mère, alors la membrane primaire est formée par la sécrétion d'hémicellulose et de fibrilles de cellulose par deux nouveaux corps cellulaires. Et toute augmentation supplémentaire de l'épaisseur de la paroi cellulaire (ou plutôt intercellulaire) se produira en raison de l'activité de deux cellules filles, qui sécrètent des côtés opposés des substances de la membrane cellulaire, qui s'épaississent en superposant de plus en plus de nouvelles couches. Dès le début, la libération des substances matricielles s'effectue en approchant la membrane plasmique des vésicules de l'appareil de Golgi, en les fusionnant avec la membrane et en libérant leur contenu au-delà du cytoplasme. Ici, à l'extérieur de la cellule, sur sa membrane plasmique, se produisent la synthèse et la polymérisation des fibrilles de cellulose. C’est ainsi que se forme progressivement une membrane cellulaire secondaire. Il est difficile de déterminer et de pouvoir distinguer la coque primaire de la coque secondaire avec une précision suffisante, car elles sont reliées entre elles par plusieurs couches intermédiaires.
La masse principale de la paroi cellulaire qui a achevé sa formation est la membrane secondaire. Elle lui donne la cage forme définitive. Une fois la cellule divisée en deux cellules filles, de nouvelles cellules se développent, leur volume augmente et leur forme change ; les cellules s'allongent souvent. Parallèlement, on observe une augmentation de l’épaisseur de la membrane cellulaire et une restructuration de sa structure interne.
Lorsque la membrane cellulaire primaire est formée, sa composition contient encore peu de fibrilles de cellulose, et elles sont situées plus ou moins perpendiculairement au futur axe longitudinal de la cellule. Plus tard, pendant la période d'élongation (allongement de la cellule dû à la croissance de vacuoles dans le cytoplasme), l'orientation de ces fibrilles dirigées transversalement subit des changements passifs : les fibrilles commencent à se situer perpendiculairement les unes aux autres et finissent par se retrouver allongée plus ou moins parallèlement à l'axe longitudinal de la cellule. Le processus est en constante évolution : dans les anciennes couches (plus proches du centre de la coquille), les fibrilles subissent des déplacements passifs, et le dépôt de nouvelles fibrilles dans les couches internes (les plus proches de la membrane cellulaire) se poursuit conformément à la conception originale de la coquille. Ce processus crée la possibilité de glissement des fibrilles les unes par rapport aux autres, et la restructuration du renforcement de la membrane cellulaire est possible grâce à l'état gélatineux des composants de sa matrice. Par la suite, lorsque l'hémicellulose est remplacée par la lignine dans la matrice, la mobilité des fibrilles diminue fortement, la coque devient dense et une lignification se produit.
Riz. 3. Schéma de croissance de la membrane cellulaire depuis son initiation lors de la division cellulaire (I) jusqu'à la maturation complète (V) : 1 - membrane primaire ; 2 -- couches de la coque secondaire ; 3 - coque tertiaire ; B - vacuoles ; SP - plaque médiane ; PM - membranes plasmiques de deux cellules voisines
Souvent, sous la coque secondaire, on trouve une coque tertiaire, qui peut être considérée comme un résidu séché de la couche dégénérée du cytoplasme lui-même.
Il est à noter que lorsque les cellules végétales se divisent, la formation de la membrane primaire n'est pas dans tous les cas précédée par la formation d'une plaque cellulaire. Ainsi, chez l'algue verte Spirogyra, de nouvelles cloisons transversales apparaissent par formation de saillies sur les parois latérales de la cellule d'origine, qui, en s'agrandissant progressivement vers le centre de la cellule, ferment et divisent la cellule en deux.
Comme déjà mentionné, si une cellule est privée de sa membrane dans un environnement aqueux hypotonique, alors une lyse, une rupture de la cellule, se produira. Il s’est avéré qu’en sélectionnant des concentrations appropriées de sels et de sucres, il est possible d’égaliser la pression osmotique à l’extérieur et à l’intérieur des cellules privées de leurs membranes. Dans ce cas, ces protoplastes acquièrent forme sphérique(sphéroplastes). Si dans l'environnement où se trouvent les protoplastes, il y a une quantité suffisante de nutriments et de sels (le Ca2+ est nécessaire parmi eux), alors les cellules restaurent et régénèrent à nouveau leur membrane cellulaire. De plus, en présence d'hormones (auxines), elles sont capables de se diviser et de créer des colonies cellulaires, ce qui peut donner lieu à la croissance de la plante entière dont la cellule a été extraite.
Le principal composant fibreux de la paroi cellulaire de grands groupes de champignons (basibiomycètes, ascomycètes, zygomycètes) est la chitine ; c'est un polysaccharide dont le principal saccharide est la N-acétylglucosamine. La composition de la paroi cellulaire des champignons, en plus de la chitine, peut comprendre des substances matricielles, des glycoprotéines et diverses protéines, synthétisé dans le cytoplasme et libéré par la cellule vers l'extérieur.
Nouvelle coque formé lors de la division cellulaire au stade final de la mitose - télophase. Après la divergence des chromosomes, un groupe de petites vésicules membranaires apparaît dans le plan équatorial des cellules, qui commencent à fusionner les unes avec les autres dans la partie centrale des cellules. Ce processus de fusion de petites vacuoles se produit du centre de la cellule vers la périphérie et se poursuit jusqu'à ce que les vésicules membranaires fusionnent entre elles et avec la membrane plasmique de la surface latérale de la cellule. C'est ainsi qu'il se forme plaque cellulaire, ou phragmoplaste.
DANS partie centrale il contient une substance matricielle amorphe qui remplit les vésicules en fusion. Il a été prouvé que ces vacuoles primaires proviennent de membranes de l'appareil de Golgi. La paroi cellulaire primaire comprend également une petite quantité de protéine (environ 10 %), riche en hydroxyproline et possédant de nombreuses chaînes oligosaccharidiques courtes, ce qui définit cette protéine comme une glycoprotéine.
Après la formation de la plaque médiane, le protoplaste des cellules voisines y dépose la membrane primaire. La couche de cellulose déposée lors de la croissance cellulaire est appelée membrane cellulaire primaire.
En plus de la cellulose, de l'hémicellulose et de la pectine, les coquilles primaires contiennent également protéine structurelle – glycoprotéine. Les coquilles primaires peuvent également être lignifiées, bien que, en règle générale, la lignine ne leur soit pas caractéristique. Cependant, la partie la plus caractéristique de la coque primaire est composant pectine. Il donne de la plasticité à la coquille, lui permettant de s'étirer au fur et à mesure que les organes s'allongent : racine, tige, feuille. Les substances pectiques sont capables de gonfler considérablement, de sorte que les coquilles primaires contiennent beaucoup d'eau (60 à 90 %). Les hémicelluloses et les substances pectiques représentent 50 à 60 % du poids sec de la coque primaire, la teneur en cellulose ne dépasse pas 30 %, les protéines structurelles occupent jusqu'à 10 %.
Le processus continu de libération des substances matricielles s'effectue en approchant la membrane plasmique des vésicules de l'appareil de Golgi, en les fusionnant avec la membrane et en libérant leur contenu au-delà du cytoplasme. Ici, à l'extérieur de la cellule, sur sa membrane plasmique, se produisent la synthèse et la polymérisation des fibrilles de cellulose. C'est ainsi qu'il se forme progressivement paroi cellulaire secondaire. Il est difficile de déterminer avec suffisamment de précision et de pouvoir distinguer la coque primaire de la coque secondaire, puisqu'elles sont interconnectées par plusieurs couches intermédiaires.
En gros Une fois que la paroi cellulaire a terminé sa formation, elle forme la membrane secondaire. Il donne à la cellule sa forme définitive. Une fois la cellule divisée en deux cellules filles, de nouvelles cellules se développent, leur volume augmente et leur forme change ; les cellules s'allongent souvent. Parallèlement, on observe une augmentation de l’épaisseur de la membrane cellulaire et une restructuration de sa structure interne.
Pendant la période d'élongation, les fibrilles commencent à se situer à angle droit les unes par rapport aux autres et finissent par s'allonger plus ou moins parallèlement à l'axe longitudinal de la cellule. Le processus est en constante évolution : dans les anciennes couches (plus proches du centre de la coquille), les fibrilles subissent des déplacements passifs, et le dépôt de nouvelles fibrilles dans les couches internes (les plus proches des membranes cellulaires) se poursuit conformément à la conception originale de la coquille. Ce processus crée la possibilité de glissement des fibrilles les unes par rapport aux autres, et la restructuration du renforcement de la membrane cellulaire est possible grâce à l'état gélatineux des composants de sa matrice. Par la suite, lors du remplacement de l'hémicellulose dans la matrice par de la lignine la mobilité des fibrilles diminue fortement, la coquille devient dense et une lignification se produit. Contenu diverses substances environ ceci : très peu d'eau, cellulose 40-50 %, lignine 25-30 %, hémicellulose 20-30 % et pratiquement aucune substance pectine.
La coque secondaire n'est pas toujours uniformément répartie. Dans certaines cellules spécialisées conductrices d’eau, cela ressemble à anneaux ou bandes spirales. Ces cellules conservent la capacité de s’étirer longitudinalement même après la mort.
Souvent trouvé sous la coque secondaire coque tertiaire, qui peut être considéré comme un reste séché de la couche dégénérée du cytoplasme lui-même.
Paroi cellulaire (membrane cellulaire)- un trait caractéristique d'une cellule végétale qui la distingue d'une cellule animale. La paroi cellulaire donne à la cellule sa forme spécifique. Les cellules végétales cultivées sur des milieux nutritifs spéciaux, dans lesquels la paroi est éliminée par voie enzymatique, prennent toujours une forme sphérique. La paroi cellulaire donne de la force à la cellule et protège le protoplaste ; elle équilibre la pression de turgescence et empêche ainsi la rupture du plasmalemme. L’ensemble des parois cellulaires forme le squelette interne qui soutient le corps végétal et lui confère sa résistance mécanique.
La paroi cellulaire est incolore et transparente et transmet facilement la lumière du soleil. Habituellement, les murs sont saturés d’eau. Le système de paroi cellulaire transporte l'eau et les composés de faible poids moléculaire qui y sont dissous (transport le long de l'apoplaste).
La paroi cellulaire est principalement constituée de polysaccharides, qui peuvent être divisés en substances squelettiques Et substances matricielles.
Substance squelettique la paroi cellulaire végétale est cellulose (fibre), qui est un bêta-1,4-D-glucane. C'est la substance organique la plus répandue dans la biosphère. Les molécules de cellulose sont de très longues chaînes non ramifiées ; elles sont situées parallèlement les unes aux autres en groupes de plusieurs dizaines et sont maintenues ensemble par de nombreuses liaisons hydrogène. Par conséquent, microfibrilles, qui créent la charpente structurelle du mur et déterminent sa résistance. Les microfibrilles de cellulose ne sont visibles qu'au microscope électronique, leur diamètre est de 10 à 30 nm et leur longueur atteint plusieurs microns.
La cellulose est insoluble et ne gonfle pas dans l'eau. Il est très chimiquement inerte, insoluble dans les solvants organiques, les alcalis concentrés et les acides dilués. Les microfibrilles de cellulose sont élastiques et très résistantes à la traction (semblables à l'acier). Ces propriétés déterminent l'utilisation généralisée de la cellulose et de ses produits. La production mondiale de fibre de coton, constituée presque entièrement de cellulose, est de 1,5 10 7 tonnes par an. La poudre à canon sans fumée, la soie et la viscose en acétate, la cellophane et le papier sont obtenus à partir de cellulose. Réaction qualitative réalisée sur cellulose avec un réactif chlore-zinc-iode, la paroi cellulaire cellulosique devient bleu-violet.
Chez les champignons, la substance squelettique de la paroi cellulaire est chitine– un polysaccharide construit à partir de résidus de glucosamine. La chitine est encore plus durable que la cellulose.
Les microfibrilles sont immergées dans un milieu amorphe matrice, généralement un gel plastique saturé d’eau. La matrice est un mélange complexe de polysaccharides dont les molécules sont constituées de résidus de plusieurs sucres différents et sont des chaînes plus courtes et ramifiées que la cellulose. Les polysaccharides matriciels déterminent les propriétés de la paroi cellulaire telles qu'un fort gonflement, une perméabilité élevée à l'eau et aux composés de faible poids moléculaire qui y sont dissous, ainsi que des propriétés d'échange de cations. Les polysaccharides matriciels sont divisés en deux groupes - substances pectiques Et hémicelluloses.
Substances pectiques gonfler fortement ou se dissoudre dans l'eau. Ils sont facilement détruits par les alcalis et les acides. Les représentants les plus simples des substances pectiques sont solubles dans l'eau acides pectiques– produits de polymérisation de l'acide alpha-D-galacturonique (jusqu'à 100 unités), liés par des liaisons 1,4 en chaînes linéaires (alpha-1,4-D-galacturonane). Acides pectiques (pectines)– il s’agit de composés polymères de poids moléculaire plus élevé (100-200 unités) de l’acide alpha-D-galacturonique, dans lesquels les groupes carboxyle sont partiellement méthylés. Pectates Et pectinés– les sels de calcium et de magnésium des acides pectiques et pectiques. Les acides pectiques, pectates et pectinates sont solubles dans l'eau en présence de sucres et d'acides organiques pour former des gels denses.
Les parois cellulaires des plantes contiennent principalement protopectines– les polymères de haut poids moléculaire de l'acide polygalacturonique méthoxylé avec des arabinanes et des galactanes ; chez les plantes dicotylédones, les chaînes galacturonanes contiennent une faible quantité de rhamnose. Les protopectines sont insolubles dans l'eau.
Hémicelluloses les chaînes ramifiées sont-elles construites à partir de résidus de sucre neutres, le glucose, le galactose, le mannose et le xylose sont plus courants ; degré de polymérisation 50-300. Les hémicelluloses sont chimiquement plus stables que les substances pectiques ; elles sont plus difficiles à hydrolyser et gonflent moins facilement dans l’eau. Les hémicelluloses peuvent se déposer dans les parois cellulaires des graines comme substances de réserve (palmier dattier, kaki). Les substances pectiques et les hémicelluloses sont reliées par des transitions mutuelles. En plus des polysaccharides, une protéine structurelle spéciale est présente dans la matrice de la paroi cellulaire. Il est lié aux résidus de sucre arabinose et est donc une glycoprotéine.
Les polysaccharides matriciels font plus que simplement remplir les espaces entre les microfibrilles de cellulose. Leurs chaînes sont disposées de manière ordonnée et forment de nombreuses liaisons entre elles et avec les microfibrilles, ce qui augmente considérablement la résistance de la paroi cellulaire.
Les parois cellulaires végétales subissent souvent des modifications chimiques. Lignification, ou lignification se produit lorsqu'il est déposé dans la matrice lignine– un composé polymère de nature phénolique, insoluble dans l’eau. La paroi cellulaire lignifiée perd son élasticité, sa dureté et sa résistance à la compression augmentent fortement et sa perméabilité à l'eau diminue. Les réactifs pour la lignine sont : 1) phloroglucinol Et acide chlorhydrique concentré ou acide sulfurique(les murs lignifiés acquièrent une couleur rouge cerise) et 2) sulfate aniline, sous l'influence de laquelle les parois lignifiées deviennent jaune citron. La lignification est caractéristique des parois cellulaires du tissu conducteur du xylème (bois) et du tissu mécanique du sclérenchyme.
Échantillonnage, ou subérinisation se produit à la suite du dépôt de polymères hydrophobes à l’intérieur de la paroi cellulaire - subérine Et la cire. La subérine est un mélange d'esters d'acides gras polymères. Les monomères de cire sont des alcools gras et des esters de cire. La cire est facilement extraite par les solvants organiques et fond rapidement pour former des cristaux. La subérine est un composé amorphe qui ne fond pas et ne se dissout pas dans les solvants organiques. La subérine et la cire, formant des couches parallèles alternées, tapissent toute la cavité cellulaire de l'intérieur sous la forme d'un film. Le film de subérine est pratiquement imperméable à l'eau et aux gaz, donc après sa formation, la cellule meurt généralement. La subérisation est caractéristique des parois cellulaires du tissu tégumentaire du liège. Le réactif pour les parois cellulaires subérisées est SoudanIII, couleur rouge orangé.
Coutinisation Les parois externes des cellules du tissu épidermique sont exposées. Kutine Et la cire déposé en couches alternées sur la surface externe de la paroi cellulaire sous la forme d'un film - cuticules. La cutine est un composé polymère ressemblant à de la graisse, similaire en nature chimique et en propriétés à la subérine. La cuticule protège la plante de l’évaporation excessive de l’eau de la surface de la plante. Vous pouvez le colorer avec un réactif SoudanIII de couleur rouge orangé.
Minéralisation la paroi cellulaire est due au dépôt dans la matrice grande quantité substances minérales, le plus souvent de la silice (oxyde de silicium), moins souvent de l'oxalate et du carbonate de calcium. Minéraux donner au mur dureté et fragilité. Les dépôts de silice sont caractéristiques des cellules épidermiques des prêles, des carex et des graminées. La rigidité des tiges et des feuilles acquise à la suite de la silicification sert d'agent protecteur contre les escargots et réduit également considérablement l'appétence et la valeur nutritionnelle des plantes.
Certaines cellules spécialisées ont mucus paroi cellulaire. Dans ce cas, au lieu d'une paroi secondaire cellulosique, des polysaccharides acides amorphes et hautement hydratés sont déposés sous la forme vase Et des gencives, proche par sa nature chimique des substances pectines. Le mucus se dissout bien dans l'eau pour former des solutions de mucus. Les gencives sont collantes et s'étirent en fils. Une fois secs, ils ont une consistance cornée. Lorsque le mucus se dépose, le protoplaste est progressivement poussé vers le centre de la cellule, son volume ainsi que celui de la vacuole diminuent progressivement. Finalement, la cavité cellulaire peut se remplir complètement de mucus et la cellule meurt. Dans certains cas, le mucus peut traverser la paroi cellulaire primaire jusqu'à la surface. L'appareil de Golgi joue un rôle majeur dans la synthèse et la sécrétion du mucus.
Le mucus sécrété par les cellules végétales remplit diverses fonctions. Ainsi, le mucus de la coiffe racinaire sert de lubrifiant, facilitant la croissance de l’extrémité de la racine dans le sol. Les glandes muqueuses des plantes insectivores (rosrosa) sécrètent du mucus piège auquel les insectes se collent. Le mucilage sécrété par les cellules externes de l'enveloppe de la graine (lin, coing, plantain) fixe la graine à la surface du sol et protège la plantule du dessèchement. Le mucus est coloré avec un réactif bleu de méthylène en couleur bleue.
La libération des gommes se produit généralement lorsque les plantes sont blessées. Par exemple, des écoulements de gomme provenant des zones blessées des troncs et des branches sont souvent observés chez les cerises et les prunes. La colle de cerise est une gomme durcie. La gomme remplit une fonction protectrice en recouvrant la plaie de la surface. Les gommes sont formées principalement dans les plantes ligneuses de la famille des légumineuses (acacia, adragante astragale) et des Rosacées de la sous-famille des pruniers (cerisier, prunier, abricot). Les gommes et les mucilages sont utilisés en médecine.
La paroi cellulaire est un produit de l'activité vitale du protoplaste. Les polysaccharides matriciels, la glycoprotéine murale, la lignine et le mucus se forment dans l'appareil de Golgi. La synthèse de la cellulose, la formation et l'orientation des microfibrilles sont réalisées par le plasmalemme. Un rôle majeur dans l'orientation des microfibrilles appartient aux microtubules, situés parallèlement aux microfibrilles déposées près du plasmalemme. Si les microtubules sont détruits, seules des cellules isodiamétriques se forment.
La formation de la paroi cellulaire commence lors de la division cellulaire. Dans le plan de division, se forme une plaque cellulaire, couche unique commune aux deux cellules filles. Il se compose de substances pectiques ayant une consistance semi-liquide ; il n'y a pas de cellulose. Dans une cellule adulte, la plaque cellulaire est préservée, mais subit des modifications, c'est pourquoi on l'appelle médian, ou plaque intercellulaire (substance intercellulaire) (riz. 2.16). La plaque médiane est généralement très fine et presque impossible à distinguer.
Immédiatement après la formation de la plaque cellulaire, les protoplastes des cellules filles commencent à former leur propre paroi cellulaire. Il se dépose de l'intérieur à la fois sur la surface de la plaque cellulaire et sur la surface d'autres parois cellulaires ayant appartenu auparavant à la cellule mère. Après la division, la cellule entre dans la phase de croissance par élongation, provoquée par une absorption osmotique intense de l'eau par la cellule associée à la formation et à la croissance de la vacuole centrale. La pression de turgescence commence à étirer la paroi, mais elle ne se déchire pas car de nouvelles portions de microfibrilles et de substances matricielles s'y déposent constamment. Le dépôt de nouvelles portions de matériau se produit uniformément sur toute la surface du protoplaste, de sorte que l'épaisseur de la paroi cellulaire ne diminue pas.
Les parois des cellules en division et en croissance sont appelées primaire. Ils contiennent beaucoup (60 à 90 %) d'eau. La matière sèche est dominée par les polysaccharides matriciels (60-70 %), la teneur en cellulose ne dépasse pas 30 % et il n'y a pas de lignine. L'épaisseur de la paroi primaire est très faible (0,1-0,5 microns).
Pour de nombreuses cellules, le dépôt de la paroi cellulaire cesse simultanément à l’arrêt de la croissance cellulaire. Ces cellules sont entourées d’une fine paroi primaire jusqu’à la fin de leur vie ( riz. 2.16).
Riz. 2.16. Cellule de parenchyme avec une paroi primaire.
Dans d’autres cellules, le dépôt sur la paroi continue même après que la cellule ait atteint sa taille finale. Dans ce cas, l'épaisseur de la paroi augmente et le volume occupé par la cavité cellulaire diminue. Ce processus est appelé épaississement secondaire murs, et le mur lui-même est appelé secondaire(riz. 2.17).
Le mur secondaire peut être considéré comme supplémentaire, remplissant principalement une fonction de support mécanique. C'est la paroi secondaire qui est responsable des propriétés du bois, des fibres textiles et du papier. Le mur secondaire contient nettement moins d’eau que le mur primaire ; elle est dominée par des microfibrilles de cellulose (40 à 50 % du poids de matière sèche), situées parallèlement les unes aux autres. Parmi les polysaccharides matriciels, les hémicelluloses (20 à 30 %) sont typiques et il existe très peu de substances pectiques. Les parois cellulaires secondaires subissent généralement une lignification. Dans les parois secondaires non lignifiées (fibres libériennes de lin, poils de coton), la teneur en cellulose peut atteindre 95 %. La teneur élevée et l'orientation strictement ordonnée des microfibrilles déterminent une haute propriétés mécaniques murs secondaires. Souvent, les cellules qui ont une paroi cellulaire lignifiée secondaire meurent une fois l’épaississement secondaire terminé.
La lame médiane colle les cellules voisines entre elles. S'il est dissous, les parois cellulaires perdent le contact les unes avec les autres et se séparent. Ce processus est appelé macération. La macération naturelle est assez courante, dans laquelle les substances pectiques de la plaque médiane sont transformées en un état soluble à l'aide de l'enzyme pectinase puis lavées à l'eau (fruits trop mûrs de poire, melon, pêche, banane). On observe souvent une macération partielle, dans laquelle la plaque médiane ne se dissout pas sur toute la surface, mais uniquement dans les coins des cellules. En raison de la pression de turgescence, les cellules voisines à ces endroits sont arrondies, entraînant la formation espaces intercellulaires(riz. 2.16). Les espaces intercellulaires forment un réseau ramifié unique, rempli de vapeur d'eau et de gaz. Ainsi, les espaces intercellulaires améliorent les échanges gazeux des cellules.
Une caractéristique du mur secondaire est son dépôt inégal au-dessus du mur primaire, ce qui fait que des zones non épaissies subsistent dans le mur secondaire - pores. Si la paroi secondaire n’atteint pas une épaisseur importante, les pores ressemblent à de petites dépressions. Dans les cellules dotées d'une paroi secondaire solide, les pores en section transversale ont la forme de canaux radiaux s'étendant de la cavité cellulaire à la paroi primaire. Selon la forme du canal poreux, il existe deux types de pores : simple Et à propos bordé(Fig. 2.17).

Riz. 2.17. Types de pores: A – cellules avec parois secondaires et nombreux pores simples ; B – une paire de pores simples ; B – paire de pores bordés.
U pores simples Le diamètre du canal poreux est le même sur toute sa longueur et a la forme d'un cylindre étroit. Les pores simples sont caractéristiques des cellules du parenchyme, des fibres libériennes et du bois.
Les pores de deux cellules adjacentes ont tendance à apparaître l’un en face de l’autre. Ces pores communs Ils ressemblent à un seul canal, séparé par une fine cloison entre la plaque médiane et la paroi primaire. Cette combinaison de deux pores de parois adjacentes de cellules voisines est appelée paires de pores et fonctionne comme un tout. La section de mur qui les sépare s'appelle film de clôture pores, ou membrane poreuse. Dans les cellules vivantes, le film de fermeture des pores est imprégné de nombreux plasmodesmes(riz. 2.18).
Plasmodesmes ne se trouvent que dans les cellules végétales. Ce sont des brins de cytoplasme qui traversent la paroi des cellules adjacentes. Le nombre de plasmodesmes dans une cellule est très important - de plusieurs centaines à des dizaines de milliers ; les plasmodesmes sont généralement collectés en groupes. Le diamètre du canal plasmodesmal est de 30 à 60 nm. Ses parois sont tapissées de plasmalemme, en continuité avec le plasmalemme des cellules adjacentes. Au centre des plasmodesmes se trouve un cylindre membranaire - tige centrale plasmodesmes, en continuité avec les membranes des éléments du réticulum endoplasmique des deux cellules. Entre la tige centrale et la membrane plasmique du canal se trouve l'hyaloplasme, en continuité avec l'hyaloplasme des cellules adjacentes.

Riz. 2.18. Plasmodesmes au microscope électronique (schéma) : 1 – sur une coupe longitudinale ; 2 – sur une section transversale ; PL– le plasmalemme ; Californie– bâtonnet central des plasmodesmes ; urgence– élément du réticulum endoplasmique.
Ainsi, les protoplastes cellulaires ne sont pas complètement isolés les uns des autres, mais communiquent via des canaux plasmodesmes. Ils assurent le transport intercellulaire d'ions et de petites molécules et transmettent également des stimuli hormonaux. Grâce aux plasmodesmes, les protoplastes des cellules d'un organisme végétal forment un tout unique appelé simple, et le transport de substances à travers les plasmodesmes est appelé symplastique Contrairement à apoplastique transport le long des parois cellulaires et des espaces intercellulaires.
U pores bordés(riz. 2.17) le canal se rétrécit fortement lors du dépôt de la paroi cellulaire, de sorte que l'ouverture interne du pore, ouvrant dans la cavité cellulaire, est beaucoup plus étroite que l'ouverture externe, attenante à la paroi primaire. Les pores bordés sont caractéristiques des cellules en voie de mort précoce des éléments du bois conducteurs d’eau. Dans ceux-ci, le canal poreux s'étend en forme d'entonnoir vers le film de fermeture et la paroi secondaire pend sous la forme d'un rouleau sur la partie élargie du canal, formant une chambre poreuse. Le nom de pore bordé vient du fait que vu de la surface, le trou interne ressemble à un petit cercle ou à une fente étroite, tandis que le trou extérieur semble border le trou interne sous la forme d'un cercle de plus grand diamètre ou d'une largeur plus large. fente.
Les pores facilitent le transport de l'eau et des solutés d'une cellule à l'autre sans réduire la résistance de la paroi cellulaire.