Fonctions des lysosomes et des mitochondries. En quoi les processus du métabolisme énergétique dans les mitochondries diffèrent-ils des processus se produisant dans les lysosomes ? Fonctions dans la cellule
Lire aussi
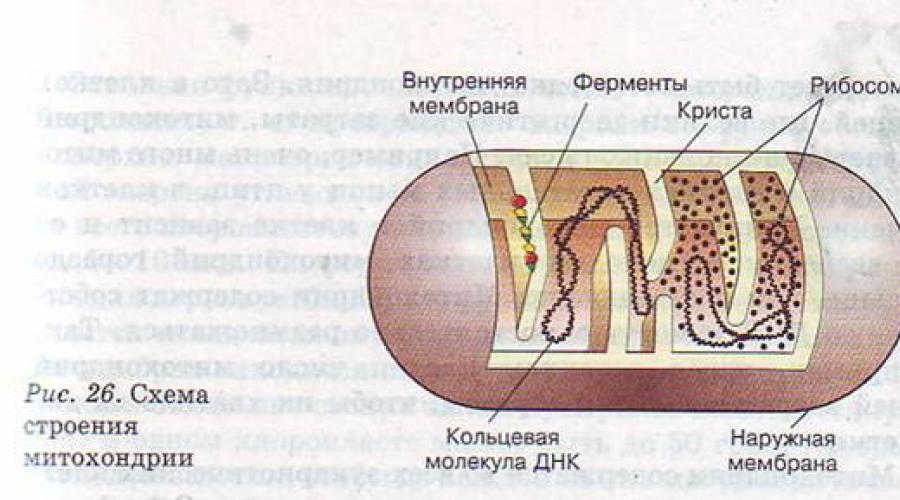
Lysosomes. Mitochondries. Plastides
1. Quelle est la structure et les fonctions ATP?
2. Quels types de plastes connaissez-vous ?
Lorsque divers nutriments pénètrent dans une cellule par phagocytose ou pinocytose, ils doivent être digérés. Où écureuils doit être décomposé en acides aminés individuels, polysaccharides - en molécules de glucose ou de fructose, lipides- au glycérol et aux acides gras. Pour que la digestion intracellulaire soit possible, la vésicule phagocytaire ou pinocytaire doit fusionner avec le lysosome (Fig. 25). Un lysosome est une petite bulle, de seulement 0,5 à 1,0 microns de diamètre, contenant un large éventail d'enzymes capables de détruire les nutriments. Un lysosome peut contenir 30 à 50 enzymes différentes.

Structures cellulaires permanentes, organes cellulaires qui assurent l'accomplissement de fonctions spécifiques au cours de la vie de la cellule - stockage et transmission d'informations génétiques, transfert de substances, synthèse et transformation de substances et d'énergie, division, mouvement, etc.
Aux organoïdes (organites) des cellules eucaryotes se rapporter:
- chromosomes ;
- membrane cellulaire;
- les mitochondries ;
- Complexe de Golgi ;
- réticulum endoplasmique ;
- ribosomes;
- microtubules;
- microfilaments;
- lysosomes.
Les cellules animales contiennent également des centrioles et des microfibrilles, et les cellules végétales contiennent des plastes qui leur sont propres.
Parfois, le noyau dans son ensemble est classé parmi les organites des cellules eucaryotes.
Procaryotes manquent de la plupart des organites, ils n'ont qu'une membrane cellulaire et des ribosomes, qui diffèrent des ribosomes cytoplasmiques des cellules eucaryotes.
Les cellules eucaryotes spécialisées peuvent avoir des structures complexes basées sur des organelles universelles, telles que les microtubules et les centrioles, principaux composants des flagelles et des cils. Les microfibrilles sont à la base des tono- et des neurofibrilles. Les structures spéciales des organismes unicellulaires, telles que les flagelles et les cils (construits de la même manière que dans les cellules multicellulaires), remplissent la fonction d'organes de mouvement.
Le plus souvent dans la littérature moderne les termes « organoïdes " Et " organites " sont utilisés comme synonymes.
Structures communes aux cellules animales et végétales
| Illustration schématique | Structure | Les fonctions |
|
| Membrane plasmique (plasmalemme, membrane cellulaire) | Deux couches de lipides (bicouche) entre deux couches de protéines | Une barrière sélectivement perméable qui régule les échanges entre la cellule et l'environnement |
|
| Cœur | Le plus grand organite, enfermé dans une coquille de deux membranes, imprégné pores nucléaires. Contient chromatine- sous cette forme, les chromosomes déroulés sont en interphase. Contient également une structure appelée nucléole | Les chromosomes contiennent de l'ADN, la substance de l'hérédité. L'ADN est constitué de gènes qui régulent tous les types d'activité cellulaire. La division nucléaire est à la base de la reproduction cellulaire, et donc du processus de reproduction. Les ribosomes se forment dans le nucléole |
|
| Réticulum endoplasmique (RE) | Système de sacs à membrane aplatie - réservoirs- sous forme de tubes et de plaques. Forme une seule unité avec la membrane externe de l'enveloppe nucléaire | Si la surface du RE est recouverte de ribosomes, on parle alors de rugueux Les protéines synthétisées sur les ribosomes sont transportées à travers les réservoirs d'un tel RE. Lisse urgence(sans ribosomes) sert de site à la synthèse des lipides et des stéroïdes |
|
| Ribosomes | Très petits organites constitués de deux sous-particules - grandes et petites. Ils contiennent des protéines et de l'ARN dans des proportions à peu près égales. Les ribosomes, présents dans les mitochondries (et également dans les chloroplastes des plantes), sont encore plus petits | Site de synthèse des protéines, où diverses molécules en interaction sont maintenues dans la bonne position. Les ribosomes sont associés au RE ou sont libres dans le cytoplasme. De nombreux ribosomes peuvent se former polysome (polyribosome), dans lequel ils sont enfilés sur un seul brin d'ARN messager |
|
| Mitochondries | La mitochondrie est entourée d'une coque de deux membranes, la membrane interne forme des plis ( Cristaux). Contient une matrice contenant un petit nombre de ribosomes, une molécule d'ADN circulaire et des granules de phosphate | Au cours de la respiration aérobie, la phosphorylation oxydative et le transfert d'électrons se produisent dans les crêtes, et les enzymes impliquées dans le cycle de Krebs et l'oxydation des acides gras opèrent dans la matrice. |
|
| Appareil de Golgi | Une pile de sacs membranaires aplatis - réservoirs. À une extrémité, des piles de pochettes se forment en continu et à l’autre, elles sont entrelacées sous forme de bulles. Les piles peuvent exister sous forme de dictyosomes discrets, comme dans les cellules végétales, ou former un réseau spatial, comme dans de nombreuses cellules animales. | De nombreux matériaux cellulaires, comme les enzymes du RE, subissent des modifications dans les citernes et sont transportés dans des vésicules. L'appareil de Golgi est impliqué dans le processus de sécrétion et des lysosomes s'y forment |
|
| Lysosomes | Un simple sac membranaire sphérique (une seule membrane) rempli d'enzymes digestives (hydrolytiques). Le contenu semble homogène | Effectuer de nombreuses fonctions, toujours associées à la désintégration de toute structure ou molécule |
|
| Microcorps | L'organite n'est pas de forme sphérique tout à fait régulière, entouré d'une seule membrane. Le contenu a une structure granulaire, mais il contient parfois un cristalloïde ou un ensemble de fils. | Tous les microcorps contiennent de la catalase, une enzyme qui catalyse la dégradation du peroxyde d'hydrogène. Tous sont associés à des réactions oxydatives |
|
| Paroi cellulaire, lame médiane, plasmodesmes |
|||
| paroi cellulaire | La paroi cellulaire rigide entourant la cellule est constituée de microfibrilles de cellulose noyées dans une matrice contenant d'autres polysaccharides complexes, à savoir des hémicelluloses et des pectines. Dans certaines cellules, les parois cellulaires subissent un épaississement secondaire | Fournit un support mécanique et une protection. Grâce à cela, une pression de turgescence apparaît, ce qui améliore la fonction de support. Empêche la rupture des cellules osmotiques. Le mouvement de l’eau et des sels minéraux se produit le long de la paroi cellulaire. Diverses modifications, telles que l'imprégnation de lignine, fournissent des fonctions spécialisées |
|
| plaque intermédiaire | Une fine couche de substances pectiques (pectates de calcium et de magnésium) | Maintient les cellules ensemble |
|
| plasmodesma | Un mince filament cytoplasmique qui relie le cytoplasme de deux cellules adjacentes à travers un mince pore dans la paroi cellulaire. Le pore est tapissé d'une membrane plasmique. Le desmotubule traverse le pore, souvent relié aux deux extrémités au RE. | Unir les protoplastes des cellules voisines en un seul système continu - simple, à travers lequel les substances sont transportées entre ces cellules |
|
| Chloroplaste | Grand plaste contenant de la chlorophylle dans lequel se produit la photosynthèse. Le chloroplaste est entouré d'une double membrane et rempli de gélatineux stroma. Le stroma contient un système de membranes assemblées en piles, ou céréales. De l'amidon peut également s'y déposer. De plus, le stroma contient des ribosomes, une molécule d'ADN circulaire et des gouttelettes d'huile. | Dans cet organite se produit la photosynthèse, c'est-à-dire la synthèse de sucres et d'autres substances à partir du CO 2 et de l'eau grâce à l'énergie lumineuse capturée par la chlorophylle. L'énergie lumineuse est convertie en énergie chimique. |
|
| Grande vacuole centrale | Un sac formé d'une seule membrane appelée tonoplaste. La vacuole contient de la sève cellulaire - une solution concentrée de diverses substances, telles que des sels minéraux, sucres, pigments, acides organiques et enzymes. Dans les cellules matures, les vacuoles sont généralement grandes | Diverses substances y sont stockées, y compris les produits finaux du métabolisme. Les propriétés osmotiques de la cellule dépendent fortement du contenu de la vacuole. Parfois, la vacuole fonctionne comme un lysosome |
|
Caractéristiques comparatives de l'ARN et de l'ADN
| Panneaux | ARN | ADN |
| Emplacement dans la cage | Noyau, ribosomes, cytoplasme, mitochondries, chloroplastes | Noyau, mitochondries, chloroplastes |
| Localisation dans le noyau | Nucléole | Chromosomes |
| Structure d'une macromolécule | Chaîne polynucléotidique unique | Polymère linéaire double non ramifié, enroulé en hélice à droite |
| Monomères | Ribonucléotides | Désoxyribonucléotides |
| Composition nucléotidique | Base azotée (purine - adénine, guanine, pyrimidine - uracile, cytosine) ; ribose (glucide) : résidu d'acide phosphorique | Base azotée (purine - adénine, guanine, pyrimidine - thymine, cytosine) ; désoxyribose (glucide) : résidu d'acide phosphorique |
| Types de nucléotides | Alényle (A), guanyle (G), uridyle (U), cytidyle (C) | Alényle (A), guanyle (G), thymidyle (T), cytidyle (C) |
| Propriétés | Incapable de s'auto-doubler. Labilna | Capable d'auto-duplication selon le principe de complémentarité (réduplication) : A-T, T-A, G-C, C-G Stable |
| Les fonctions | Informationnel (ARNm) - transmet le code d'informations héréditaires sur la structure primaire de la molécule protéique ; ribosomal (ARNr) - partie des ribosomes ; transport (ARNt) - transfère les acides aminés aux ribosomes ; ARN mitochondrial et plastidique - font partie des ribosomes de ces organites | Base chimique du matériel génétique chromosomique (gène) ; Synthèse d'ADN, synthèse d'ARN, informations sur la structure des protéines |
Le contenu de l'article
CELLULE, unité élémentaire des êtres vivants. La cellule est délimitée des autres cellules ou de l'environnement extérieur par une membrane spéciale et possède un noyau ou son équivalent, dans lequel est concentrée l'essentiel de l'information chimique qui contrôle l'hérédité. La cytologie étudie la structure des cellules et la physiologie s'occupe de leur fonctionnement. La science qui étudie les tissus constitués de cellules s’appelle l’histologie.
Il existe des organismes unicellulaires dont le corps entier est constitué d'une seule cellule. Ce groupe comprend les bactéries et les protistes (protozoaires et algues unicellulaires). Parfois, ils sont également appelés acellulaires, mais le terme unicellulaire est plus souvent utilisé. Les véritables animaux multicellulaires (métazoaires) et plantes (métaphytes) contiennent de nombreuses cellules.
La grande majorité des tissus sont composés de cellules, à quelques exceptions près. Le corps des moisissures visqueuses (myxomycètes), par exemple, est constitué d'une substance homogène non divisée en cellules comportant de nombreux noyaux. Certains tissus animaux, notamment le muscle cardiaque, sont organisés de manière similaire. Le corps végétatif (thalle) des champignons est formé de fils microscopiques - des hyphes, souvent segmentés ; chacun de ces fils peut être considéré comme l'équivalent d'une cellule, bien que de forme atypique.
Certaines structures de l'organisme qui ne participent pas au métabolisme, notamment les coquillages, les perles ou la base minérale des os, ne sont pas formées par des cellules, mais par les produits de leur sécrétion. D'autres, comme le bois, l'écorce, les cornes, les cheveux et couche externe peau - non d'origine sécrétoire, mais formée de cellules mortes.
Les petits organismes, comme les rotifères, ne comportent que quelques centaines de cellules. A titre de comparaison : dans le corps humain, il y a env. 10 14 cellules, chaque seconde, 3 millions de globules rouges meurent et sont remplacés par de nouveaux, ce qui ne représente qu'un dix millionième du nombre total de cellules du corps.
Généralement, la taille des cellules végétales et animales varie de 5 à 20 microns de diamètre. Une cellule bactérienne typique est beaucoup plus petite : env. 2 microns, et le plus petit connu est de 0,2 microns.
Certaines cellules libres, comme les protozoaires comme les foraminifères, peuvent atteindre plusieurs centimètres ; ils ont toujours plusieurs cœurs. Les cellules des fibres végétales minces atteignent une longueur d'un mètre et les processus des cellules nerveuses atteignent plusieurs mètres chez les grands animaux. Avec une telle longueur, le volume de ces cellules est petit, mais la surface est très grande.
Les plus grosses cellules sont des œufs d’oiseaux non fécondés remplis de jaune. Le plus gros œuf(et donc la plus grande cellule) appartenait à un énorme oiseau disparu - apyornis ( Aepyornis). Vraisemblablement, son jaune pesait env. 3,5 kg. Le plus gros œuf parmi les espèces vivantes appartient à l'autruche ; son jaune pèse env. 0,5 kg.
En règle générale, les cellules des grands animaux et des plantes ne sont que légèrement plus grandes que celles des petits organismes. Un éléphant est plus gros qu’une souris, non pas parce que ses cellules sont plus grandes, mais principalement parce qu’il y a beaucoup plus de cellules elles-mêmes. Il existe des groupes d’animaux, comme les rotifères et les nématodes, dans lesquels le nombre de cellules du corps reste constant. Ainsi, bien que les grandes espèces de nématodes aient un plus grand nombre de cellules que les petites, la principale différence de taille est due dans ce cas à des cellules de plus grande taille.
Au sein d'un type cellulaire donné, leurs tailles dépendent généralement de la ploïdie, c'est-à-dire sur le nombre de jeux de chromosomes présents dans le noyau. Les cellules tétraploïdes (avec quatre jeux de chromosomes) sont deux fois plus volumineuses que les cellules diploïdes (avec deux jeux de chromosomes). La ploïdie d’une plante peut être augmentée en y introduisant de la colchicine, un médicament à base de plantes. Puisque les plantes exposées à cet effet ont des cellules plus grandes, elles sont elles-mêmes plus grandes. Cependant, ce phénomène ne peut être observé que chez les polyploïdes d'origine récente. Dans les plantes polyploïdes évolutives anciennes, la taille des cellules est soumise à une « régulation inverse » vers des valeurs normales malgré une augmentation du nombre de chromosomes.
STRUCTURE CELLULAIRE
Autrefois, la cellule était considérée comme une goutte plus ou moins homogène de matière organique, appelée protoplasme ou substance vivante. Ce terme est devenu obsolète après la découverte que la cellule est constituée de nombreuses structures clairement distinctes appelées organites cellulaires (« petits organes »).
Composition chimique.
En règle générale, 70 à 80 % de la masse cellulaire est constituée d'eau, dans laquelle sont dissous divers sels et substances de faible poids moléculaire. composés organiques. Les composants les plus caractéristiques d’une cellule sont les protéines et les acides nucléiques. Certaines protéines sont des composants structurels de la cellule, d'autres sont des enzymes, c'est-à-dire catalyseurs qui déterminent la vitesse et la direction des processus se produisant dans les cellules réactions chimiques. Les acides nucléiques servent de transporteurs informations héréditaires, qui est réalisé dans le processus de synthèse des protéines intracellulaires.
Les cellules contiennent souvent une certaine quantité de substances de stockage qui servent de réserve alimentaire. Les cellules végétales stockent principalement l’amidon, une forme polymère de glucides. Un autre polymère glucidique, le glycogène, est stocké dans les cellules hépatiques et musculaires. Les aliments fréquemment stockés contiennent également des graisses, bien que certaines graisses remplissent une fonction différente, à savoir qu’elles servent de composants structurels essentiels. Les protéines contenues dans les cellules (à l’exception des cellules séminales) ne sont généralement pas stockées.
Il n’est pas possible de décrire la composition typique d’une cellule, principalement parce qu’il existe de grandes différences dans la quantité de nourriture et d’eau stockée. Les cellules hépatiques contiennent par exemple 70 % d'eau, 17 % de protéines, 5 % de graisses, 2 % de glucides et 0,1 % d'acides nucléiques ; les 6 % restants proviennent de sels et de composés organiques de faible poids moléculaire, notamment d'acides aminés. Les cellules végétales contiennent généralement moins de protéines, beaucoup plus de glucides et un peu plus d'eau ; l'exception concerne les cellules qui sont au repos. Une cellule au repos d'un grain de blé, qui est une source de nutriments pour l'embryon, contient env. 12 % de protéines (principalement des protéines stockées), 2 % de matières grasses et 72 % de glucides. La quantité d'eau n'atteint le niveau normal (70 à 80 %) qu'au début de la germination des grains.
Principales parties de la cellule.
Certaines cellules, principalement végétales et bactériennes, possèdent une paroi cellulaire externe. Dans les plantes supérieures, il s'agit de cellulose. La paroi entoure la cellule elle-même, la protégeant des contraintes mécaniques. Les cellules, en particulier les cellules bactériennes, peuvent également sécréter des substances muqueuses, formant ainsi autour d'elles une capsule qui, comme la paroi cellulaire, a une fonction protectrice.
C'est à la destruction des parois cellulaires qu'est associée la mort de nombreuses bactéries sous l'influence de la pénicilline. Le fait est qu'à l'intérieur de la cellule bactérienne, la concentration de sels et de composés de faible poids moléculaire est très élevée et, par conséquent, en l'absence de paroi de renforcement, l'afflux d'eau dans la cellule provoqué par la pression osmotique peut conduire à sa rupture. La pénicilline, qui empêche la formation de sa paroi lors de la croissance cellulaire, entraîne la rupture cellulaire (lyse).
Les parois cellulaires et les capsules ne participent pas au métabolisme et peuvent souvent être séparées sans tuer la cellule. Ainsi, ils peuvent être considérés comme des parties auxiliaires externes de la cellule. Les cellules animales manquent généralement de parois cellulaires et de capsules.
La cellule elle-même se compose de trois parties principales. Sous paroi cellulaire, si elle est présente, il y a une membrane cellulaire. La membrane entoure un matériau hétérogène appelé cytoplasme. Un noyau rond ou ovale est immergé dans le cytoplasme. Ci-dessous, nous examinerons plus en détail la structure et les fonctions de ces parties de la cellule.
MEMBRANE CELLULAIRE
La membrane cellulaire est une partie très importante de la cellule. Il maintient tous les composants cellulaires ensemble et délimite les environnements interne et externe. De plus, les plis modifiés de la membrane cellulaire forment de nombreux organites cellulaires.
La membrane cellulaire est une double couche de molécules (couche bimoléculaire, ou bicouche). Il s'agit principalement de molécules de phospholipides et d'autres substances qui leur sont liées. Les molécules lipidiques ont une double nature, qui se manifeste dans leur comportement par rapport à l'eau. Les têtes des molécules sont hydrophiles, c'est-à-dire ont une affinité pour l'eau et leurs queues d'hydrocarbures sont hydrophobes. Par conséquent, lorsqu’ils sont mélangés à l’eau, les lipides forment à sa surface un film semblable à un film d’huile ; De plus, toutes leurs molécules sont orientées de la même manière : les têtes des molécules sont dans l'eau, et les queues d'hydrocarbures sont au-dessus de sa surface.
Il existe deux couches de ce type dans la membrane cellulaire, et dans chacune d'elles, les têtes des molécules sont tournées vers l'extérieur et les queues sont tournées vers l'intérieur de la membrane, l'une vers l'autre, n'entrant ainsi pas en contact avec l'eau. L'épaisseur d'une telle membrane est d'env. 7 nm. En plus des principaux composants lipidiques, il contient de grosses molécules protéiques capables de « flotter » dans la bicouche lipidique et disposées de manière à ce qu’un côté soit tourné vers l’intérieur de la cellule et que l’autre soit en contact avec l’environnement extérieur. Certaines protéines se trouvent uniquement sur la surface externe ou interne de la membrane ou ne sont que partiellement immergées dans la bicouche lipidique.
La fonction principale de la membrane cellulaire est de réguler le transport des substances vers et hors de la cellule. Étant donné que la membrane est physiquement similaire à l'huile, les substances solubles dans l'huile ou dans les solvants organiques, tels que l'éther, la traversent facilement. Il en va de même pour les gaz tels que l'oxygène et le dioxyde de carbone. En même temps, la membrane est pratiquement imperméable à la plupart des substances solubles dans l'eau, en particulier les sucres et les sels. Grâce à ces propriétés, il est capable de maintenir à l’intérieur de la cellule un environnement chimique différent de l’extérieur. Par exemple, dans le sang, la concentration en ions sodium est élevée et celle en ions potassium est faible, tandis que dans le liquide intracellulaire, ces ions sont présents dans des proportions opposées. Une situation similaire est typique pour de nombreux autres composés chimiques.
Il est évident que la cellule ne peut cependant pas être complètement isolée de environnement, puisqu'il doit recevoir les substances nécessaires au métabolisme et se débarrasser de ses produits finaux. De plus, la bicouche lipidique n’est pas complètement imperméable, même aux substances hydrosolubles, et à celles qui y pénètrent. les protéines « formant des canaux » créent des pores, ou canaux, qui peuvent s'ouvrir et se fermer (en fonction des changements dans la conformation des protéines) et, à l'état ouvert, conduire certains ions (Na +, K +, Ca 2+) le long d'un gradient de concentration . Par conséquent, la différence de concentrations à l’intérieur et à l’extérieur de la cellule ne peut être maintenue uniquement en raison de la faible perméabilité de la membrane. En fait, il contient des protéines qui remplissent la fonction de « pompe » moléculaire : elles transportent certaines substances à la fois dans et hors de la cellule, en travaillant contre un gradient de concentration. En conséquence, lorsque la concentration, par exemple, d'acides aminés à l'intérieur de la cellule est élevée et faible à l'extérieur, les acides aminés peuvent néanmoins circuler de l'environnement externe vers l'environnement interne. Ce transfert est appelé transport actif et utilise l’énergie fournie par le métabolisme. Les pompes à membrane sont très spécifiques : chacune d'entre elles est capable de transporter soit uniquement des ions d'un certain métal, soit un acide aminé, soit un sucre. Les canaux ioniques membranaires sont également spécifiques.
Une telle perméabilité sélective est physiologiquement très importante et son absence est la première preuve de mort cellulaire. Ceci est facile à illustrer avec l’exemple des betteraves. Si une racine de betterave vivante est immergée dans l’eau froide, elle conserve son pigment ; si les betteraves sont bouillies, les cellules meurent, deviennent facilement perméables et perdent leur pigment, ce qui rend l'eau rouge.
La cellule peut « avaler » de grosses molécules comme des protéines. Sous l'influence de certaines protéines, si elles sont présentes dans le liquide entourant la cellule, une invagination se produit dans la membrane cellulaire, qui se ferme alors, formant une vésicule - une petite vacuole contenant des molécules d'eau et de protéines ; Après cela, la membrane autour de la vacuole se rompt et le contenu pénètre dans la cellule. Ce processus est appelé pinocytose (littéralement « boire la cellule ») ou endocytose.
Les particules plus grosses, telles que les particules alimentaires, peuvent être absorbées de la même manière lors de ce qu'on appelle. phagocytose. En règle générale, la vacuole formée lors de la phagocytose est plus grande et la nourriture est digérée par les enzymes lysosomales à l'intérieur de la vacuole avant la rupture de la membrane environnante. Ce type de nutrition est typique des protozoaires, comme les amibes, qui se nourrissent de bactéries. Cependant, la capacité de phagocytose est caractéristique à la fois des cellules intestinales des animaux inférieurs et des phagocytes, l'un des types de globules blancs (leucocytes) des vertébrés. Dans ce dernier cas, le sens de ce processus ne réside pas dans la nutrition des phagocytes eux-mêmes, mais dans leur destruction des bactéries, virus et autres corps étrangers nocifs pour l'organisme.
Les fonctions des vacuoles peuvent être différentes. Par exemple, les protozoaires vivant dans eau fraiche, subissent un afflux osmotique constant d’eau, car la concentration de sels à l’intérieur de la cellule est beaucoup plus élevée qu’à l’extérieur. Ils sont capables de sécréter de l'eau dans une vacuole excrétrice (contractile) spéciale, qui expulse périodiquement son contenu.
Les cellules végétales ont souvent une grande vacuole centrale occupant presque toute la cellule ; le cytoplasme ne forme qu'une très fine couche entre la paroi cellulaire et la vacuole. L’une des fonctions d’une telle vacuole est l’accumulation d’eau, permettant à la cellule d’augmenter rapidement en taille. Cette capacité est particulièrement nécessaire pendant la période où les tissus végétaux se développent et forment des structures fibreuses.
Dans les tissus, aux endroits où les cellules sont étroitement liées, leurs membranes contiennent de nombreux pores formés par des protéines qui pénètrent dans la membrane - ce qu'on appelle. connexions. Les pores des cellules adjacentes sont situés l'un en face de l'autre, de sorte que les substances de faible poids moléculaire peuvent passer de cellule en cellule - ce système de communication chimique coordonne leur activité vitale. Un exemple d’une telle coordination est la division plus ou moins synchrone de cellules voisines observée dans de nombreux tissus.
CYTOPLASME
Le cytoplasme contient des membranes internes similaires à la membrane externe et forment des organites de différents types. Ces membranes peuvent être considérées comme des plis de la membrane externe ; parfois les membranes internes sont solidaires de la membrane externe, mais souvent le pli interne est délacé et le contact avec la membrane externe est interrompu. Cependant, même si le contact est maintenu, les membranes interne et externe ne sont pas toujours chimiquement identiques. En particulier, la composition des protéines membranaires diffère selon les organites cellulaires.
Réticulum endoplasmique.
Un réseau de membranes internes constitué de tubules et de vésicules s'étend de la surface cellulaire jusqu'au noyau. Ce réseau est appelé réticulum endoplasmique. Il a souvent été constaté que les tubules s'ouvrent à la surface de la cellule, et le réticulum endoplasmique joue ainsi le rôle de microvascularisation à travers laquelle environnement externe peut interagir directement avec tout le contenu de la cellule. Cette interaction a été constatée dans certaines cellules, notamment les cellules musculaires, mais il n’est pas encore clair si elle est universelle. Dans tous les cas, un certain nombre de substances sont effectivement transportées à travers ces tubules d'une partie de la cellule à une autre.
De minuscules corps appelés ribosomes recouvrent la surface du réticulum endoplasmique, en particulier près du noyau. Diamètre des ribosomes env. 15 nm, ils sont constitués pour moitié de protéines, pour moitié d’acides ribonucléiques. Leur fonction principale est la synthèse des protéines ; L'ARN messager et les acides aminés associés à l'ARN de transfert sont attachés à leur surface. Les zones du réticulum recouvertes de ribosomes sont appelées réticulum endoplasmique rugueux, et celles qui en sont dépourvues sont appelées lisses. En plus des ribosomes, diverses enzymes sont adsorbées sur le réticulum endoplasmique ou y sont fixées d'une autre manière, notamment des systèmes enzymatiques qui assurent l'utilisation de l'oxygène pour la formation de stérols et pour la neutralisation de certains poisons. Dans des conditions défavorables, le réticulum endoplasmique dégénère rapidement et son état sert donc d'indicateur sensible de la santé cellulaire.
Appareil de Golgi.
L'appareil de Golgi (complexe de Golgi) est une partie spécialisée du réticulum endoplasmique, constituée de sacs membranaires plats empilés. Il est impliqué dans la sécrétion de protéines par la cellule (il y a un emballage des protéines sécrétées dans des granules) et est donc particulièrement développé dans les cellules qui remplissent une fonction sécrétoire. Les fonctions importantes de l'appareil de Golgi incluent également la fixation de groupes glucidiques aux protéines et l'utilisation de ces protéines pour construire la membrane cellulaire et la membrane du lysosome. Chez certaines algues, les fibres de cellulose sont synthétisées dans l'appareil de Golgi.
Lysosomes
- Ce sont de petites bulles entourées d'une seule membrane. Ils bourgeonnent à partir de l'appareil de Golgi et éventuellement du réticulum endoplasmique. Les lysosomes contiennent diverses enzymes qui décomposent les grosses molécules, en particulier les protéines. En raison de leur action destructrice, ces enzymes sont en quelque sorte « enfermées » dans les lysosomes et ne sont libérées qu’en cas de besoin. Ainsi, lors de la digestion intracellulaire, les enzymes sont libérées des lysosomes dans les vacuoles digestives. Les lysosomes sont également nécessaires à la destruction des cellules ; par exemple, lors de la transformation d'un têtard en grenouille adulte, la libération d'enzymes lysosomales assure la destruction des cellules de la queue. Dans ce cas, cela est normal et bénéfique pour l’organisme, mais parfois une telle destruction cellulaire est pathologique. Par exemple, lorsque la poussière d’amiante est inhalée, elle peut pénétrer dans les cellules pulmonaires, ce qui entraîne la rupture des lysosomes, la destruction des cellules et l’apparition d’une maladie pulmonaire.
Mitochondries et chloroplastes.
Les mitochondries sont des structures en forme de sac relativement grandes avec une structure plutôt complexe. Ils sont constitués d'une matrice entourée d'une membrane interne, d'un espace intermembranaire et d'une membrane externe. La membrane interne est pliée en plis appelés crêtes. Des amas de protéines sont situés sur les crêtes. Beaucoup d’entre eux sont des enzymes qui catalysent l’oxydation des produits de dégradation des glucides ; d'autres catalysent les réactions de synthèse et d'oxydation des graisses. Les enzymes auxiliaires impliquées dans ces processus sont dissoutes dans la matrice mitochondriale.
L'oxydation des substances organiques se produit dans les mitochondries, couplée à la synthèse d'adénosine triphosphate (ATP). La dégradation de l'ATP pour former l'adénosine diphosphate (ADP) s'accompagne de la libération d'énergie, qui est dépensée pour divers processus vitaux, par exemple pour la synthèse de protéines et d'acides nucléiques, le transport de substances dans et hors de la cellule, la transmission de l'influx nerveux ou contraction musculaire. Les mitochondries sont donc des stations énergétiques qui transforment le « carburant » – les graisses et les glucides – en une forme d’énergie utilisable par la cellule, et donc par l’organisme dans son ensemble.
Les cellules végétales contiennent également des mitochondries, mais la principale source d’énergie de leurs cellules est la lumière. L'énergie lumineuse est utilisée par ces cellules pour produire de l'ATP et synthétiser des glucides à partir du dioxyde de carbone et de l'eau. La chlorophylle, un pigment qui accumule l'énergie lumineuse, se trouve dans les chloroplastes. Les chloroplastes, comme les mitochondries, possèdent des membranes internes et externes. Des excroissances de la membrane interne lors du développement des chloroplastes naissent ce qu'on appelle les chloroplastes. membranes thylakoïdes; ces derniers forment des sacs aplatis, rassemblés en piles comme une colonne de pièces de monnaie ; ces piles, appelées grana, contiennent de la chlorophylle. En plus de la chlorophylle, les chloroplastes contiennent tous les autres composants nécessaires à la photosynthèse.
Certains chloroplastes spécialisés n'effectuent pas la photosynthèse, mais ont d'autres fonctions, comme le stockage de l'amidon ou des pigments.
Autonomie relative.
À certains égards, les mitochondries et les chloroplastes se comportent comme des organismes autonomes. Par exemple, tout comme les cellules elles-mêmes, qui proviennent uniquement de cellules, les mitochondries et les chloroplastes sont formés uniquement à partir de mitochondries et de chloroplastes préexistants. Cela a été démontré dans des expériences sur des cellules végétales, dans lesquelles la formation de chloroplastes a été supprimée par l'antibiotique streptomycine, et sur des cellules de levure, où la formation de mitochondries a été supprimée par d'autres médicaments. Après de tels effets, les cellules n’ont jamais restauré les organites manquants. La raison en est que les mitochondries et les chloroplastes contiennent une certaine quantité de leur propre matériel génétique (ADN) qui code une partie de leur structure. Si cet ADN est perdu, ce qui se produit lorsque la formation des organites est supprimée, la structure ne peut pas être recréée. Les deux types d'organites possèdent leur propre système de synthèse de protéines (ribosomes et ARN de transfert), qui est quelque peu différent du système de synthèse de protéines principal de la cellule ; on sait, par exemple, que le système de synthèse des protéines des organites peut être supprimé à l'aide d'antibiotiques, alors qu'ils n'ont aucun effet sur le système principal.
L'ADN des organelles est responsable de l'essentiel de l'hérédité extrachromosomique ou cytoplasmique. L'hérédité extrachromosomique n'obéit pas aux lois mendéliennes, puisque lorsqu'une cellule se divise, l'ADN des organites est transmis aux cellules filles d'une manière différente de celle des chromosomes. L'étude des mutations qui se produisent dans l'ADN des organites et dans l'ADN chromosomique a montré que l'ADN des organites n'est responsable que d'une petite partie de la structure des organites ; la plupart de leurs protéines sont codées dans des gènes situés sur les chromosomes.
L'autonomie génétique partielle des organites considérés et les caractéristiques de leurs systèmes de synthèse des protéines ont servi de base à l'hypothèse selon laquelle les mitochondries et les chloroplastes provenaient de bactéries symbiotiques qui se sont installées dans les cellules il y a 1 à 2 milliards d'années. Un exemple moderne Une telle symbiose peut servir de petites algues photosynthétiques qui vivent à l'intérieur des cellules de certains coraux et mollusques. Les algues fournissent de l'oxygène à leurs hôtes et en reçoivent des nutriments.
Structures fibrillaires.
Le cytoplasme d'une cellule est un fluide visqueux, donc la tension superficielle donnerait à penser que la cellule est sphérique à moins que les cellules ne soient étroitement emballées. Toutefois, cela n’est généralement pas observé. De nombreux protozoaires ont des téguments ou des membranes denses qui donnent à la cellule une forme spécifique non sphérique. Néanmoins, même sans membrane, les cellules peuvent conserver une forme non sphérique du fait que le cytoplasme est structuré à l'aide de nombreuses fibres parallèles plutôt rigides. Ces derniers sont formés de microtubules creux, constitués d’unités protéiques organisées en spirale.
Certains protozoaires forment des pseudopodes – de longues et fines projections cytoplasmiques avec lesquelles ils capturent la nourriture. Les pseudopodes conservent leur forme en raison de la rigidité des microtubules. Si la pression hydrostatique augmente jusqu'à environ 100 atmosphères, les microtubules se désintègrent et la cellule prend la forme d'une goutte. Lorsque la pression revient à la normale, les microtubules se réassemblent et la cellule forme des pseudopodes. De nombreuses autres cellules réagissent de manière similaire aux changements de pression, ce qui confirme la participation des microtubules au maintien de la forme cellulaire. L’assemblage et la désintégration des microtubules, nécessaires au changement rapide de forme d’une cellule, se produisent même en l’absence de changement de pression.
Les microtubules forment également des structures fibrillaires qui servent d'organes de mouvement cellulaire. Certaines cellules ont des projections en forme de fouet appelées flagelles ou cils : leurs battements assurent le mouvement de la cellule dans l'eau. Si la cellule est immobile, ces structures poussent l’eau, les particules de nourriture et d’autres particules vers ou loin de la cellule. Les flagelles sont relativement grands et la cellule ne possède généralement qu'un, parfois plusieurs, flagelles. Les cils sont beaucoup plus petits et couvrent toute la surface de la cellule. Bien que ces structures soient caractéristiques principalement des plus simples, elles peuvent également se présenter sous des formes très organisées. Dans le corps humain, toutes les voies respiratoires sont tapissées de cils. Les petites particules qui y pénètrent sont généralement capturées par le mucus à la surface des cellules et les cils les poussent avec le mucus, protégeant ainsi les poumons. Les cellules reproductrices mâles de la plupart des animaux et de certaines plantes inférieures se déplacent à l'aide d'un flagelle.
Il existe d'autres types de mouvements cellulaires. L’un d’eux est le mouvement amiboïde. L'amibe, ainsi que certaines cellules d'organismes multicellulaires, « circulent » d'un endroit à l'autre, c'est-à-dire se déplacer en raison du courant du contenu de la cellule. Un courant constant de matière existe également à l’intérieur des cellules végétales, mais il n’entraîne pas de mouvement de la cellule dans son ensemble. Le type de mouvement cellulaire le plus étudié est la contraction des cellules musculaires ; elle est réalisée par glissement des fibrilles (fils protéiques) les unes par rapport aux autres, ce qui conduit à un raccourcissement de la cellule.
CŒUR
Le noyau est entouré d'une double membrane. L'espace très étroit (environ 40 nm) entre deux membranes est appelé périnucléaire. Les membranes nucléaires passent dans les membranes du réticulum endoplasmique et l'espace périnucléaire s'ouvre dans l'espace réticulaire. Généralement, la membrane nucléaire possède des pores très étroits. Apparemment, de grosses molécules y sont transportées, comme l'ARN messager, qui est synthétisé sur l'ADN puis pénètre dans le cytoplasme.
La majeure partie du matériel génétique se trouve dans les chromosomes du noyau cellulaire. Les chromosomes sont constitués de longues chaînes d’ADN double brin, auxquelles sont attachées des protéines basiques (c’est-à-dire alcalines). Parfois, les chromosomes ont plusieurs brins d'ADN identiques côte à côte - ces chromosomes sont appelés polytènes (multibrins). Le nombre de chromosomes varie selon les espèces. Les cellules diploïdes du corps humain contiennent 46 chromosomes, soit 23 paires.
Dans une cellule qui ne se divise pas, les chromosomes sont attachés en un ou plusieurs points à la membrane nucléaire. Dans leur état normal déroulé, les chromosomes sont si fins qu’ils ne sont pas visibles au microscope optique. Au niveau de certains locus (sections) d'un ou plusieurs chromosomes, un corps dense se forme, présent dans les noyaux de la plupart des cellules - ce qu'on appelle. nucléole. Dans les nucléoles, se produisent la synthèse et l’accumulation d’ARN utilisé pour construire les ribosomes, ainsi que d’autres types d’ARN.
LA DIVISION CELLULAIRE
Bien que toutes les cellules soient issues de la division d’une cellule précédente, toutes ne continuent pas à se diviser. Par exemple, les cellules nerveuses du cerveau, une fois formées, ne se divisent pas. Leur nombre diminue progressivement ; Le tissu cérébral endommagé n’est pas capable de récupérer par régénération. Si les cellules continuent de se diviser, elles sont alors caractérisées par un cycle cellulaire composé de deux étapes principales : l'interphase et la mitose.
L'interphase elle-même se compose de trois phases : G 1, S et G 2. Vous trouverez ci-dessous leur durée, typique des cellules végétales et animales.
G 1 (4 à 8 heures). Cette phase commence immédiatement après la naissance de la cellule. Durant la phase G 1, la cellule, à l'exception des chromosomes (qui ne changent pas), augmente sa masse. Si la cellule ne se divise pas davantage, elle reste dans cette phase.
S (6 à 9 heures). La masse cellulaire continue d'augmenter et un doublement (duplication) de l'ADN chromosomique se produit. Cependant, les chromosomes restent de structure unique, bien que doublés en masse, puisque deux copies de chaque chromosome (chromatides) sont toujours reliées entre elles sur toute leur longueur.
G2. La masse de la cellule continue d'augmenter jusqu'à atteindre environ le double de sa masse d'origine, puis la mitose se produit.
Une fois les chromosomes dupliqués, chacune des cellules filles devrait recevoir un ensemble complet de chromosomes. La simple division cellulaire ne peut pas y parvenir – ce résultat est obtenu grâce à un processus appelé mitose. Sans entrer dans les détails, le début de ce processus doit être considéré comme l'alignement des chromosomes dans le plan équatorial de la cellule. Ensuite, chaque chromosome se divise longitudinalement en deux chromatides, qui commencent à diverger dans des directions opposées, devenant ainsi des chromosomes indépendants. En conséquence, un ensemble complet de chromosomes se trouve aux deux extrémités de la cellule. La cellule se divise ensuite en deux et chaque cellule fille reçoit un ensemble complet de chromosomes.
Ce qui suit est une description de la mitose dans une cellule animale typique. Elle est généralement divisée en quatre étapes.
I. Prophase. Une structure cellulaire spéciale - le centriole - double (ce doublement se produit parfois pendant la période S de l'interphase) et les deux centrioles commencent à diverger vers les pôles opposés du noyau. La membrane nucléaire est détruite ; en même temps, des protéines spéciales se combinent (s'agrègent), formant des microtubules sous forme de fils. Les centrioles, désormais situés aux pôles opposés de la cellule, ont un effet organisateur sur les microtubules, qui s'alignent ainsi radialement, formant une structure rappelant l'apparence d'une fleur d'aster (« étoile »). D'autres fils de microtubules s'étendent d'un centriole à l'autre, formant ce qu'on appelle. fuseau de fission. À ce moment-là, les chromosomes sont en spirale, ressemblant à un ressort. Ils sont clairement visibles au microscope optique, surtout après coloration. En prophase, les chromosomes sont divisés, mais les chromatides restent attachées par paires dans la zone du centromère - un organite chromosomique dont la fonction est similaire à celle du centriole. Les centrioles ont également un effet organisateur sur les filaments du fuseau, qui s'étendent désormais d'un centriole à l'autre et de celui-ci à un autre centriole.
II. Métaphase. Les chromosomes, jusqu'à présent disposés de manière aléatoire, commencent à se déplacer, comme s'ils étaient tirés par des fils de fuseau attachés à leurs centromères, et s'alignent progressivement dans le même plan dans une certaine position et à égale distance des deux pôles. Les centromères situés dans le même plan avec les chromosomes forment ce qu'on appelle. plaque équatoriale. Les centromères reliant les paires de chromatides se divisent, après quoi les chromosomes frères sont complètement séparés.
III. Anaphase. Les chromosomes de chaque paire se déplacent dans des directions opposées vers les pôles, comme s'ils étaient entraînés par des fils de fuseau. Dans ce cas, des fils se forment également entre les centromères des chromosomes appariés.
IV. Télophase. Dès que les chromosomes se rapprochent des pôles opposés, la cellule elle-même commence à se diviser le long du plan dans lequel se trouvait la plaque équatoriale. En conséquence, deux cellules se forment. Les fils du fuseau sont détruits, les chromosomes se déroulent et deviennent invisibles, et une membrane nucléaire se forme autour d'eux. Les cellules reviennent à la phase G1 de l'interphase. L'ensemble du processus de mitose prend environ une heure.
Les détails de la mitose varient quelque peu selon les différents types de cellules. Une cellule végétale typique forme un fuseau mais manque de centrioles. Chez les champignons, la mitose se produit à l’intérieur du noyau, sans désintégration préalable de la membrane nucléaire.
La division cellulaire elle-même, appelée cytokinèse, n'a pas de lien strict avec la mitose. Parfois, une ou plusieurs mitoses se produisent sans division cellulaire ; En conséquence, des cellules multinucléées se forment, que l'on trouve souvent dans les algues. Si le noyau est retiré d’un œuf d’oursin par micromanipulation, le fuseau continue de se former et l’œuf continue de se diviser. Cela montre que la présence de chromosomes n’est pas une condition nécessaire à la division cellulaire.
La reproduction par mitose est appelée reproduction asexuée, reproduction végétative ou clonage. C'est le plus aspect important– génétique : avec une telle reproduction, il n'y a pas de divergence des facteurs héréditaires chez la progéniture. Les cellules filles qui en résultent sont génétiquement exactement les mêmes que la cellule mère. La mitose est le seul moyen d'auto-reproduction chez les espèces qui n'ont pas de reproduction sexuée, comme de nombreux organismes unicellulaires. Cependant, même chez les espèces à reproduction sexuée, les cellules du corps se divisent par mitose et proviennent d’une seule cellule, l’œuf fécondé, et sont donc toutes génétiquement identiques. Les plantes supérieures peuvent se reproduire de manière asexuée (par mitose) par semis et vrilles (un exemple bien connu est celui des fraises).
La reproduction sexuée des organismes est réalisée à l'aide de cellules spécialisées, appelées. gamètes - ovocytes (ovules) et spermatozoïdes (sperme). Les gamètes fusionnent pour former une seule cellule : un zygote. Chaque gamète est haploïde, c'est-à-dire possède un ensemble de chromosomes. Au sein de l'ensemble, tous les chromosomes sont différents, mais chaque chromosome de l'ovule correspond à l'un des chromosomes du spermatozoïde. Le zygote contient donc déjà une paire de chromosomes correspondant les uns aux autres, appelés homologues. Les chromosomes homologues sont similaires car ils possèdent les mêmes gènes ou leurs variantes (allèles) qui déterminent des caractéristiques spécifiques. Par exemple, l'un des chromosomes appariés peut avoir un gène codant pour le groupe sanguin A et l'autre peut avoir une variante codant pour le groupe sanguin B. Les chromosomes du zygote provenant de l'ovule sont maternels et ceux provenant du sperme sont paternels.
À la suite de divisions mitotiques répétées, un organisme multicellulaire ou de nombreuses cellules libres naissent du zygote résultant, comme cela se produit chez les protozoaires à reproduction sexuée et dans les algues unicellulaires.
Lorsque les gamètes se forment, l’ensemble diploïde de chromosomes présents dans le zygote doit être réduit de moitié. Si cela ne se produisait pas, la fusion des gamètes à chaque génération entraînerait un doublement de l'ensemble des chromosomes. La réduction du nombre haploïde de chromosomes se produit à la suite d'une division de réduction - ce qu'on appelle. la méiose, qui est une variante de la mitose.
Clivage et recombinaison.
La particularité de la méiose est que lors de la division cellulaire, la plaque équatoriale est formée de paires de chromosomes homologues, et non de chromosomes individuels dupliqués, comme dans la mitose. Les chromosomes appariés, dont chacun reste unique, divergent vers les pôles opposés de la cellule, la cellule se divise et, par conséquent, les cellules filles reçoivent la moitié du jeu de chromosomes par rapport au zygote.
Par exemple, supposons que l’ensemble haploïde soit constitué de deux chromosomes. Chez le zygote (et par conséquent dans toutes les cellules de l'organisme qui produit des gamètes), les chromosomes maternels A et B et les chromosomes paternels A" et B" sont présents. Durant la méiose, ils peuvent se diviser comme suit :
La chose la plus importante dans cet exemple est le fait que lorsque les chromosomes divergent, l'ensemble maternel et paternel d'origine n'est pas nécessairement formé, mais la recombinaison des gènes est possible, comme dans les gamètes AB" et A"B dans le diagramme ci-dessus.
Supposons maintenant que la paire de chromosomes AA" contienne deux allèles : un Et b– un gène qui détermine les groupes sanguins A et B. De même, la paire de chromosomes « BB » contient des allèles m Et n un autre gène qui détermine les groupes sanguins M et N. La séparation de ces allèles peut se dérouler comme suit :
Évidemment, les gamètes résultants peuvent contenir l’une des combinaisons suivantes d’allèles des deux gènes : suis, milliard, bm ou un.
S’il y a plus de chromosomes, alors les paires d’allèles se sépareront indépendamment selon le même principe. Cela signifie que les mêmes zygotes peuvent produire des gamètes avec différentes combinaisons d’allèles génétiques et donner naissance à différents génotypes chez la progéniture.
Division méiotique.
Les deux exemples illustrent le principe de la méiose. En fait, la méiose est un processus beaucoup plus complexe, puisqu’elle implique deux divisions successives. L'essentiel dans la méiose est que les chromosomes ne sont doublés qu'une seule fois, tandis que la cellule se divise deux fois, ce qui réduit le nombre de chromosomes et l'ensemble diploïde se transforme en un ensemble haploïde.
Lors de la prophase de la première division, les chromosomes homologues se conjuguent, c'est-à-dire qu'ils se réunissent par paires. Grâce à ce processus très précis, chaque gène se retrouve face à son homologue sur un autre chromosome. Les deux chromosomes doublent alors, mais les chromatides restent reliées entre elles par un centromère commun.
En métaphase, les quatre chromatides connectées s’alignent pour former une plaque équatoriale, comme s’il s’agissait d’un chromosome dupliqué. Contrairement à ce qui se passe lors de la mitose, les centromères ne se divisent pas. De ce fait, chaque cellule fille reçoit une paire de chromatides toujours reliées par le centromère. Lors de la deuxième division, les chromosomes, déjà individuels, s'alignent à nouveau, formant, comme lors de la mitose, une plaque équatoriale, mais leur dédoublement ne se produit pas lors de cette division. Les centromères se divisent alors et chaque cellule fille reçoit une chromatide.
Division cytoplasmique.
À la suite de deux divisions méiotiques d'une cellule diploïde, quatre cellules se forment. Lorsque les cellules reproductrices mâles se forment, quatre spermatozoïdes d’approximativement la même taille sont obtenus. Lors de la formation des œufs, la division du cytoplasme se produit de manière très inégale : une cellule reste grande, tandis que les trois autres sont si petites qu'elles sont presque entièrement occupées par le noyau. Ces petites cellules, soi-disant les corps polaires servent uniquement à accueillir les chromosomes excédentaires formés à la suite de la méiose. La majeure partie du cytoplasme nécessaire au zygote reste dans une cellule : l'œuf.
Conjugaison et croisement.
Lors de la conjugaison, les chromatides des chromosomes homologues peuvent se briser puis se rejoindre dans un nouvel ordre, échangeant des sections comme suit :

Cet échange de sections de chromosomes homologues est appelé croisement. Comme indiqué ci-dessus, le croisement conduit à l’émergence de nouvelles combinaisons d’allèles de gènes liés. Donc, si les chromosomes originaux avaient des combinaisons UN B Et un B, puis après avoir traversé, ils contiendront Un B Et un B. Ce mécanisme d'émergence de nouvelles combinaisons de gènes complète l'effet du tri indépendant des chromosomes qui se produit pendant la méiose. La différence est que le croisement sépare les gènes sur le même chromosome, tandis que le tri indépendant ne sépare que les gènes sur des chromosomes différents.
GÉNÉRATIONS ALTERNÉES
CELLULES PRIMITIVES : PROCARYOTES
Tout ce qui précède s’applique aux cellules des plantes, des animaux, des protozoaires et des algues unicellulaires, collectivement appelées eucaryotes. Les eucaryotes ont évolué à partir d'une forme plus simple, les procaryotes, qui sont désormais représentés par des bactéries, notamment des archéobactéries et des cyanobactéries (ces dernières autrefois appelées algues bleu-vert). Comparées aux cellules eucaryotes, les cellules procaryotes sont plus petites et possèdent moins d’organites cellulaires. Ils possèdent une membrane cellulaire mais sont dépourvus de réticulum endoplasmique et les ribosomes flottent librement dans le cytoplasme. Les mitochondries sont absentes, mais les enzymes oxydatives sont généralement fixées à la membrane cellulaire, qui devient ainsi l'équivalent des mitochondries. Les procaryotes manquent également de chloroplastes et la chlorophylle, si elle est présente, se présente sous la forme de très petits granules.
Les procaryotes n'ont pas de noyau entouré d'une membrane, bien que l'emplacement de l'ADN puisse être identifié par sa densité optique. L’équivalent d’un chromosome est un brin d’ADN, généralement circulaire, auquel sont attachées beaucoup moins de protéines. La chaîne d'ADN est attachée à la membrane cellulaire en un point. Il n’y a pas de mitose chez les procaryotes. Il est remplacé par le processus suivant : l'ADN se double, après quoi la membrane cellulaire commence à se développer entre les points d'attache adjacents de deux copies de la molécule d'ADN, qui divergent donc progressivement. La cellule finit par se diviser entre les points d’attache des molécules d’ADN, formant deux cellules, chacune possédant sa propre copie de l’ADN.
DIFFÉRENCIATION CELLULAIRE
Les plantes et les animaux multicellulaires ont évolué à partir d’organismes unicellulaires dont les cellules, après division, sont restées ensemble pour former une colonie. Initialement, toutes les cellules étaient identiques, mais une évolution ultérieure a donné lieu à une différenciation. Tout d’abord, les cellules somatiques (c’est-à-dire les cellules du corps) et les cellules germinales se différencient. La différenciation ultérieure est devenue plus compliquée - de plus en plus de types de cellules différents sont apparus. L'ontogenèse - le développement individuel d'un organisme multicellulaire - se répète dans Plan général ce processus évolutif (phylogénie).
Physiologiquement, les cellules se différencient en partie en améliorant l'une ou l'autre caractéristique commune à toutes les cellules. Par exemple, la fonction contractile est améliorée dans les cellules musculaires, ce qui peut être le résultat d'une amélioration du mécanisme qui effectue les mouvements amiboïdes ou d'autres types de cellules moins spécialisées. Un exemple similaire est celui des cellules racinaires à parois minces avec leurs processus, ce qu'on appelle. les poils absorbants, qui servent à absorber les sels et l’eau ; à un degré ou à un autre, cette fonction est inhérente à toutes les cellules. Parfois, la spécialisation est associée à l’acquisition de nouvelles structures et fonctions – un exemple est le développement d’un organe locomoteur (flagellum) dans le sperme.
La différenciation au niveau cellulaire ou tissulaire a été étudiée de manière assez détaillée. Nous savons par exemple que cela se produit parfois de manière autonome, c'est-à-dire un type de cellule peut se transformer en un autre quel que soit le type de cellules des cellules voisines. Cependant, ce qu'on appelle L'induction embryonnaire est un phénomène dans lequel un type de tissu stimule la différenciation des cellules d'un autre type dans une direction donnée.
Dans le cas général, la différenciation est irréversible, c'est-à-dire les cellules hautement différenciées ne peuvent pas se transformer en un autre type de cellule. Cependant, ce n’est pas toujours le cas, notamment dans les cellules végétales.
Les différences de structure et de fonction sont finalement déterminées par les types de protéines synthétisées dans la cellule. Puisque la synthèse des protéines est contrôlée par les gènes et que l'ensemble des gènes dans toutes les cellules du corps est le même, la différenciation doit dépendre de l'activation ou de l'inactivation de certains gènes dans divers types cellules. La régulation de l'activité des gènes se produit au niveau de la transcription, c'est-à-dire formation d’ARN messager en utilisant l’ADN comme modèle. Seuls les gènes transcrits produisent des protéines. Les protéines synthétisées peuvent bloquer la transcription, mais parfois aussi l'activer. De plus, comme les protéines sont le produit de gènes, certains gènes peuvent contrôler la transcription d’autres gènes. Les hormones, notamment les stéroïdes, interviennent également dans la régulation de la transcription. Les gènes très actifs peuvent être dupliqués (doublés) plusieurs fois pour produire davantage d’ARN messager.
Le développement de tumeurs malignes a souvent été considéré comme un cas particulier de différenciation cellulaire. Cependant, l’apparition de cellules malignes est le résultat de modifications de la structure de l’ADN (mutation) et non des processus de transcription et de traduction en protéines de l’ADN normal.
MÉTHODES D'ÉTUDE DES CELLULES
Microscope optique.
Dans l’étude de la forme et de la structure des cellules, le premier outil était le microscope optique. Sa résolution est limitée par des dimensions comparables à la longueur d'onde de la lumière (0,4 à 0,7 μm pour lumière visible). Cependant, de nombreux éléments de la structure cellulaire sont de taille beaucoup plus petite.
Une autre difficulté est que la plupart des composants cellulaires sont transparents et ont un indice de réfraction presque identique à celui de l’eau. Pour améliorer la visibilité, des colorants ayant des affinités différentes pour différents composants cellulaires sont souvent utilisés. La coloration est également utilisée pour étudier la chimie cellulaire. Par exemple, certains colorants se lient préférentiellement aux acides nucléiques et révèlent ainsi leur localisation dans la cellule. Une petite proportion de colorants, appelés colorants intravitales, peut être utilisée pour colorer des cellules vivantes, mais généralement les cellules doivent d'abord être fixées (à l'aide de substances coagulantes protéiques) avant de pouvoir être colorées. Cm. HISTOLOGIE.
Avant les tests, les cellules ou les morceaux de tissus sont généralement incorporés dans de la paraffine ou du plastique, puis coupés en sections très fines à l'aide d'un microtome. Cette méthode est largement utilisée dans les laboratoires cliniques pour identifier les cellules tumorales. En plus de la microscopie optique conventionnelle, d'autres méthodes optiques d'étude des cellules ont été développées : la microscopie à fluorescence, la microscopie à contraste de phase, la spectroscopie et l'analyse par diffraction des rayons X.
Microscope électronique.
Un microscope électronique a une résolution d'env. 1 à 2 nm. Ceci est suffisant pour étudier les grosses molécules de protéines. Il est généralement nécessaire de colorer et de contraster l'objet avec des sels métalliques ou des métaux. Pour cette raison, et aussi parce que les objets sont examinés sous vide, seules les cellules tuées peuvent être étudiées au microscope électronique.
Autoradiographie.
Si un isotope radioactif absorbé par les cellules au cours du métabolisme est ajouté au milieu, sa localisation intracellulaire peut alors être détectée par autoradiographie. Avec cette méthode, de fines sections de cellules sont placées sur un film. Le film s'assombrit aux endroits où se trouvent les isotopes radioactifs.
Centrifugation.
Pour l'étude biochimique des composants cellulaires, les cellules doivent être détruites - mécaniquement, chimiquement ou par ultrasons. Les composants libérés sont en suspension dans le liquide et peuvent être isolés et purifiés par centrifugation (le plus souvent selon un gradient de densité). Généralement, ces composants purifiés conservent une activité biochimique élevée.
Cultures cellulaires.
Certains tissus peuvent être divisés en cellules individuelles afin que celles-ci restent vivantes et soient souvent capables de se reproduire. Ce fait confirme définitivement l'idée de la cellule comme unité de vie. Une éponge, un organisme multicellulaire primitif, peut être séparée en cellules en la frottant à travers un tamis. Après un certain temps, ces cellules se reconnectent et forment une éponge. Les tissus embryonnaires animaux peuvent être dissociés à l’aide d’enzymes ou d’autres moyens qui affaiblissent les liens entre les cellules.
L'embryologiste américain R. Harrison (1879-1959) fut le premier à montrer que les cellules embryonnaires et même certaines cellules matures pouvaient croître et se multiplier en dehors du corps dans un environnement approprié. Cette technique, appelée culture cellulaire, a été mise au point par le biologiste français A. Carrel (1873-1959). Les cellules végétales peuvent également être cultivées, mais comparées aux cellules animales, elles forment des amas plus gros et sont plus fermement attachées les unes aux autres, de sorte que des tissus se forment au fur et à mesure de la croissance de la culture, plutôt que des cellules individuelles. En culture cellulaire, une plante adulte entière, comme une carotte, peut être cultivée à partir d’une seule cellule.
Microchirurgie.
À l’aide d’un micromanipulateur, des parties individuelles de la cellule peuvent être supprimées, ajoutées ou modifiées d’une manière ou d’une autre. Une grande cellule d'amibe peut être divisée en trois composants principaux : la membrane cellulaire, le cytoplasme et le noyau, puis ces composants peuvent être réassemblés pour former une cellule vivante. De cette manière, des cellules artificielles constituées de composants de différents types d’amibes peuvent être obtenues.
Si l'on tient compte du fait qu'il semble possible de synthétiser artificiellement certains composants cellulaires, alors les expériences d'assemblage de cellules artificielles pourraient être la première étape vers la création de nouvelles formes de vie en laboratoire. Puisque chaque organisme se développe à partir d'une seule cellule, la méthode de production de cellules artificielles permet en principe de construire des organismes d'un type donné, tout en utilisant des composants légèrement différents de ceux trouvés dans les cellules existantes. En réalité, cependant, une synthèse complète de tous les composants cellulaires n’est pas requise. La structure de la plupart, sinon de la totalité, des composants d’une cellule est déterminée par les acides nucléiques. Ainsi, le problème de la création de nouveaux organismes se résume à la synthèse de nouveaux types d'acides nucléiques et à leur remplacement par des acides nucléiques naturels dans certaines cellules.
Fusion cellulaire.
Un autre type de cellules artificielles peut être obtenu en fusionnant des cellules de la même espèce ou d'espèces différentes. Pour réaliser la fusion, les cellules sont exposées à des enzymes virales ; dans ce cas, les surfaces externes de deux cellules sont collées ensemble, la membrane entre elles est détruite et une cellule se forme dans laquelle deux ensembles de chromosomes sont enfermés dans un noyau. Il est possible de fusionner des cellules de différents types ou à différents stades de division. Grâce à cette méthode, il a été possible d’obtenir des cellules hybrides d’une souris et d’un poulet, d’un humain et d’une souris, et d’un humain et d’un crapaud. De telles cellules ne sont hybrides qu'au début, et après de nombreuses la division cellulaire perdre la plupart des chromosomes d’une espèce ou de l’autre. Le produit final devient, par exemple, essentiellement une cellule de souris avec peu ou pas de gènes humains présents. La fusion de cellules normales et malignes est particulièrement intéressante. Dans certains cas, les hybrides deviennent malins, dans d'autres non, c'est-à-dire les deux propriétés peuvent se manifester à la fois comme dominantes et récessives. Ce résultat n’est pas inattendu, car la malignité peut être causée par divers facteurs et possède un mécanisme complexe.
Littérature:
Ham A., Cormack D. Histologie, tome 1. M., 1982
Alberts B., Bray D., Lewis J., Raff M., Roberts K., Watson J. Biologie cellulaire moléculaire, tome 1. M., 1994
1. Structure similaire des cellules végétales et animales - preuve
A) leur relation
B) origine commune des organismes de tous les règnes
B) l'origine des plantes à partir d'animaux
D) complications des organismes en cours d'évolution
D) unité du monde organique
E) diversité des organismes
Répondre
2. Quelles fonctions le complexe de Golgi remplit-il ?
A) synthétise des substances organiques à partir de substances inorganiques
B) décompose les biopolymères en monomères
C) accumule des protéines, des lipides, des glucides synthétisés dans la cellule
D) assure le conditionnement et le retrait des substances de la cellule
D) oxyde les substances organiques en substances inorganiques
E) participe à la formation des lysosomes
Répondre
3. Établir une correspondance entre la caractéristique d'un organisme et le groupe pour lequel il est caractéristique : 1-procaryotes, 2-virus.
A) structure cellulaire du corps
B) la présence de son propre métabolisme
C) intégration de son propre ADN dans l’ADN de la cellule hôte
D) se compose de acide nucléique et coque protéique
D) reproduction par division en deux
E) la capacité d'inverser la transcription
Répondre
A1 B1 C2 D2 D1 E2
4. Les autotrophes incluent
A) plantes à spores
B) moules
B) algues unicellulaires
D) bactéries chimiotrophes
D) virus
E) la plupart des protozoaires
Répondre
5. Établir la séquence des processus se produisant pendant la méiose
A) disposition des paires de chromosomes homologues dans le plan équatorial
B) conjugaison, croisement de chromosomes homologues
B) localisation dans le plan équatorial et divergence des chromosomes frères
D) formation de quatre noyaux haploïdes
D) divergence des chromosomes homologues
Répondre
5a. Quels signes caractérisent la méiose ?
A) la présence de deux divisions se succédant
B) la formation de deux cellules avec la même information héréditaire
B) divergence des chromosomes homologues dans différentes cellules
D) formation de cellules filles diploïdes
D) absence d'interphase avant la première division
E) conjugaison et croisement des chromosomes
Répondre
6. Établir une correspondance entre les caractéristiques de la gamétogenèse et son type : 1-ovogenèse, 2-spermatogenèse
A) une grande cellule sexuelle est formée
B) des cellules guides sont formées
C) de nombreux petits gamètes se forment
D) les nutriments sont stockés dans l'une des quatre cellules
D) des gamètes mobiles se forment
Répondre
A1 B1 C2 D1 D2
7. Établir la séquence des processus se produisant dans une cellule avec des chromosomes en interphase et en mitose ultérieure
A) disposition des chromosomes dans le plan équatorial
B) Réplication de l'ADN et formation de chromosomes bichromatides
B) spiralisation des chromosomes
D) divergence des chromosomes frères vers les pôles cellulaires
Répondre
7+. Quelles structures cellulaires subissent les plus grands changements pendant la mitose ?
Un noyau
B) cytoplasme
B) ribosomes
D) lysosomes
D) centre cellulaire
E) chromosomes
Répondre
8. Établir une correspondance entre la structure de l'organite et son type : 1 centre cellulaire, 2-ribosome
A) se compose de deux cylindres situés perpendiculairement
B) se compose de deux sous-unités
B) formé de microtubules
D) contient des protéines qui assurent le mouvement des chromosomes
D) contient des protéines et de l'acide nucléique
Répondre
A1 B2 C1 D1 D2
9. Établir la séquence des processus se produisant pendant la phagocytose
A) entrée de monomères dans le cytoplasme
B) capture des nutriments par la membrane cellulaire
B) hydrolyse des polymères en monomères
D) formation d'une vésicule phagocytotique à l'intérieur de la cellule
D) fusion de la vésicule phagocytaire avec le lysosome
Répondre
Répondre
12. Les principes de base de la théorie cellulaire nous permettent de tirer des conclusions sur
A) l'influence de l'environnement sur la condition physique
B) parenté des organismes
D) le développement des organismes du simple au complexe
E) la possibilité d'une génération spontanée de vie à partir de matière inanimée
Répondre
12+. Les principes de base de la théorie cellulaire nous permettent de tirer des conclusions sur
A) migration biogénique des atomes
B) parenté des organismes
C) l'origine des plantes et des animaux d'un ancêtre commun
D) l'apparition de la vie sur Terre il y a environ 4,5 milliards d'années
D) structure similaire des cellules de tous les organismes
E) relations entre la nature vivante et inanimée
Répondre
12++. Quelles dispositions la théorie cellulaire contient-elle ?
A) De nouvelles cellules se forment à la suite de la division de la cellule mère
B) Les cellules sexuelles contiennent un ensemble haploïde de chromosomes
B) Les cellules ont une composition chimique similaire
D) La cellule est l'unité de développement de tous les organismes
D) Les cellules tissulaires de toutes les plantes et de tous les animaux ont une structure identique
E) Toutes les cellules contiennent des molécules d'ADN
Répondre
13. La similitude entre les cellules fongiques et animales est qu'elles ont
A) une coquille de substance semblable à la chitine
B) le glycogène comme glucide de stockage
B) noyau formé
D) vacuoles avec sève cellulaire
D) mitochondries
E) plastes
A) former des tissus et des organes
B) participer au processus de fécondation
B) toujours haploïde
D) avoir un ensemble diploïde de chromosomes
D) se forment au cours du processus de méiose
E) diviser par mitose
Répondre
A1 B2 C2 D1 D2 E1
17. En quoi les mitochondries diffèrent-elles des lysosomes ?
A) avoir des membranes externes et internes
B) avoir de nombreuses excroissances - crêtes
C) participer aux processus de libération d'énergie
D) en eux, l'acide pyruvique est oxydé en dioxyde de carbone et en eau
D) en eux, les biopolymères sont décomposés en monomères
E) participer au métabolisme
Répondre