Funcția lizozomilor și mitocondriilor. Cum diferă procesele de metabolism energetic din mitocondrii de procesele care au loc în lizozomi? Funcționează într-o cușcă
Citeste si
Lizozomi. Mitocondriile. plastide
1. Care este structura și funcțiile ATP?
2. Ce tipuri de plastide cunoașteți?
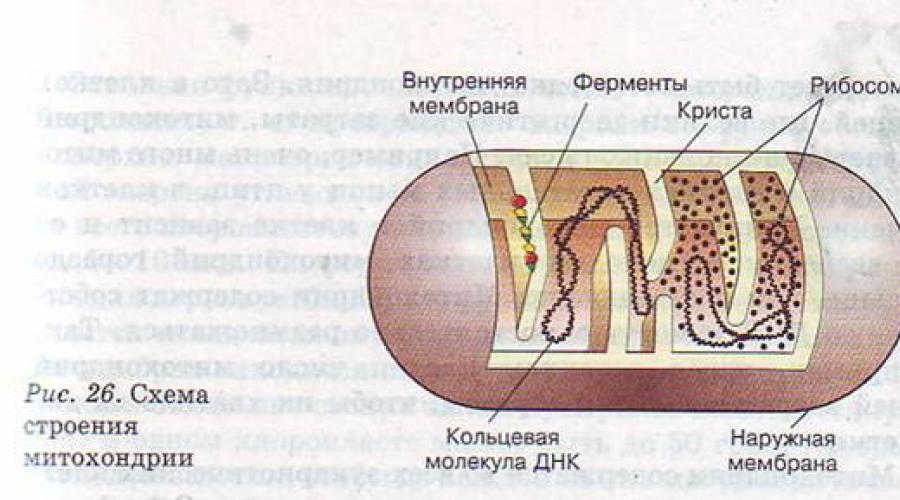
Când diverși nutrienți intră în celulă prin fagocitoză sau pinocitoză, aceștia trebuie digerați. în care veverite trebuie să se descompună în aminoacizi individuali, polizaharide - în molecule de glucoză sau fructoză, lipide- la glicerol și acizi grași. Pentru ca digestia intracelulară să devină posibilă, vezicula fagocitară sau pinocitară trebuie să fuzioneze cu lizozomul (Fig. 25). Lizozomul este o veziculă mică, cu un diametru de doar 0,5-1,0 microni, care conține un set mare de enzime care pot distruge substanțele alimentare. Un lizozom poate conține 30-50 de enzime diferite.

Structuri celulare permanente, organe celulare care asigură îndeplinirea unor funcții specifice în procesul vieții celulare - stocarea și transmiterea informațiilor genetice, transferul de substanțe, sinteza și transformarea substanțelor și energiei, diviziunea, mișcarea etc.
La organele (organele) ale celulelor eucariote raporta:
- cromozomi;
- membrana celulara;
- mitocondriile;
- complexul Golgi;
- reticul endoplasmatic;
- ribozomi;
- microtubuli;
- microfilamente;
- lizozomi.
În celulele animale, există și centrioli, microfibrile și în celulele vegetale - plastide specifice numai acestora.
Uneori, nucleul în ansamblu se referă și la organelele celulelor eucariote.
procariote lipsiți de majoritatea organitelor, au doar o membrană celulară și ribozomi care diferă de ribozomii citoplasmatici ai celulelor eucariote.
Celulele eucariote specializate pot avea structuri complexe bazate pe organite universale, cum ar fi microtubuli și centrioli, principalele componente ale flagelilor și cililor. Microfibrilele stau la baza tono- și neurofibrilelor. Structurile speciale ale organismelor unicelulare, cum ar fi flagelii și cilii (construite în același mod ca în celulele multicelulare), îndeplinesc funcția de organe de mișcare.
Mai des în literatura modernă, termenii " organele " Și " organele sunt folosite ca sinonime.
Structuri comune celulelor animale și vegetale
| Reprezentare schematică | Structura | Funcții |
|
| Membrana plasmatica (plasmalema, membrana celulara) | Două straturi de lipide (dublu strat) între două straturi de proteine | Bariera selectiv permeabilă care reglează schimbul dintre celulă și mediu |
|
| Miez | Cel mai mare organel închis într-o înveliș de două membrane, a pătruns porii nucleari. Conține cromatina- în această formă, cromozomii nerăușiți sunt în interfază. Conține și o structură numită nucleol | Cromozomii conțin ADN - substanța eredității.ADN-ul este format din gene care reglează toate tipurile de activitate celulară. Diviziunea nucleară stă la baza reproducerii celulelor și, prin urmare, a procesului de reproducere. Ribozomii se formează în nucleol |
|
| Reticul endoplasmatic (RE) | Sistem de pungi cu membrană aplatizată - cisterne- sub formă de tuburi și plăci. Formează un întreg integral cu membrana exterioară a învelișului nuclear | Dacă suprafața ER este acoperită cu ribozomi, atunci se numește grungy.Proteina sintetizată pe ribozomi este transportată de-a lungul cisternelor unui astfel de RE. Neted ER(fără ribozomi) servește ca loc pentru sinteza lipidelor și steroizilor |
|
| Ribozomi | Organele foarte mici, formate din două subparticule - mari și mici. Conțin proteine și ARN în proporții aproximativ egale. Ribozomii găsiți în mitocondrii (precum și în cloroplaste - în plante) sunt și mai mici. | Locul sintezei proteinelor unde diferite molecule care interacționează sunt ținute în poziția corectă. Ribozomii sunt asociați cu RE sau se află liber în citoplasmă. Se pot forma mulți ribozomi polizom (poliribozom), în care sunt înșirate pe o singură catenă de ARN mesager |
|
| Mitocondriile | Mitocondria este înconjurată de o teacă de două membrane, membrana interioară formează pliuri ( cristae). Conține o matrice care conține un număr mic de ribozomi, o moleculă circulară de ADN și granule de fosfat | În timpul respirației aerobe, fosforilarea oxidativă și transferul de electroni au loc în crestae, iar enzimele implicate în ciclul Krebs și oxidarea acizilor grași lucrează în matrice. |
|
| aparate Golgi | Teanc de pungi cu membrană turtite - cisterne. La un capăt, stivele pungii sunt formate continuu, iar la celălalt capăt sunt strânse sub formă de bule. Stivele pot exista ca dictiozomi discreti, ca în celulele vegetale, sau pot forma o rețea spațială, ca în multe celule animale. | Multe materiale celulare, cum ar fi enzimele din ER, suferă modificări în cisterne și sunt transportate în vezicule. Aparatul Golgi este implicat în procesul de secreție, iar în el se formează lizozomi. |
|
| Lizozomi | Un sac membranar sferic simplu (membrană unică) umplut cu enzime digestive (hidrolitice). Conținutul pare a fi omogen | Îndeplinește multe funcții, întotdeauna asociate cu degradarea oricăror structuri sau molecule |
|
| microcorp | Organela nu are o formă sferică corectă, înconjurată de o singură membrană. Conținutul este granulat, dar ocazional conține un cristaloid sau un grup de filamente. | Toți microcorpii conțin catalază, o enzimă care catalizează descompunerea peroxidului de hidrogen. Toate sunt asociate cu reacții oxidative. |
|
| Peretele celular, lamina medie, plasmodesmele |
|||
| perete celular | Peretele celular rigid care înconjoară celula este format din microfibrile de celuloză scufundate într-o matrice, care include și alte polizaharide complexe, și anume hemiceluloze și substanțe pectinice. În unele celule, pereții celulari suferă o îngroșare secundară | Oferă suport mecanic și protecție. Datorită acesteia, apare presiunea turgenței, care contribuie la întărirea funcției de sprijin. Previne ruptura osmotică a celulei. Apa și sărurile minerale se deplasează de-a lungul peretelui celular. Diverse modificări, cum ar fi impregnarea cu lignină, asigură funcții specializate |
|
| placa de mijloc | Strat subțire de pectine (pectate de calciu și magneziu) | Ține celulele împreună |
|
| plasmodesmă | Un filament citoplasmatic subțire care conectează citoplasma a două celule învecinate printr-un por subțire din peretele celular. Porul este căptușit cu o membrană plasmatică Un desmotubul trece prin por, adesea conectat la ambele capete la ER | Combinați protoplastele celulelor învecinate într-un singur sistem continuu - symplast prin care substanţele sunt transportate între aceste celule |
|
| cloroplast | O plastidă mare, care conține clorofilă, în care are loc fotosinteza. Cloroplastul este înconjurat de o membrană dublă și umplut cu gelatinos stroma. Stroma conține un sistem de membrane stive, sau boabe. Poate conține și amidon. În plus, stroma conține ribozomi, o moleculă circulară de ADN și picături de ulei. | În acest organel are loc fotosinteza, adică sinteza zaharurilor și a altor substanțe din CO 2 și apă datorită energiei luminoase captate de clorofilă.Energia luminii este transformată în substanțe chimice. |
|
| Vacuola centrală mare | Un sac alcătuit dintr-o singură membrană numită tonoplast. Vacuola conține seva celulară - o soluție concentrată de diferite substanțe, cum ar fi saruri minerale, zaharuri, pigmenți, acizi organici și enzime. În celulele mature, vacuolele sunt de obicei mari | Aici sunt depozitate diferite substanțe, inclusiv produsele finale ale metabolismului. Proprietățile osmotice ale celulei depind în mare măsură de conținutul vacuolei. Uneori, vacuola acționează ca un lizozom |
|
Caracteristici comparative ale ARN-ului și ADN-ului
| semne | ARN | ADN |
| Locația în celulă | Nucleu, ribozomi, citoplasmă, mitocondrii, cloroplaste | Nucleu, mitocondrii, cloroplaste |
| Locație în miez | nucleol | Cromozomii |
| Structura macromoleculei | Un singur lanț de polinucleotide | Polimer liniar dublu, neramificat, înfăşurat într-o spirală dreaptă |
| Monomerii | Ribonucleotide | Dezoxiribonucleotide |
| Compoziția nucleotidei | Baza azotata (purina - adenina, guanina, pirimidina - uracil, citozina); riboză (glucide): reziduu de acid fosforic | Baza azotata (purina - adenina, guanina, pirimidina - timina, citozina); dezoxiriboză (carbohidrat): un reziduu de acid fosforic |
| Tipuri de nucleotide | Alenil (A), guanil (G), uridil (U), citidil (C) | Alenil (A), guanil (G), timidil (T), citidil (C) |
| Proprietăți | Nu este capabil să se autodubleze. Labil | Capabil de autodublare conform principiului complementarității (reduplicare): A-T, T-A, G-C, C-G Stabil |
| Funcții | Informațional (ARNm) - transmite codul informațiilor ereditare despre structura primară a moleculei proteice; ribozomal (ARNr) - face parte din ribozom; transport (ARNt) - transportă aminoacizi la ribozomi; ARN mitocondrial și plastid - fac parte din ribozomii acestor organite | Baza chimică a materialului genetic cromozomial (genă); Sinteza ADN, sinteza ARN, informații despre structura proteinelor |
Conținutul articolului
CELULA, unitate elementară a vieții. Celula este delimitată de alte celule sau de mediul extern printr-o membrană specială și are un nucleu sau echivalentul acestuia, în care se concentrează cea mai mare parte a informațiilor chimice care controlează ereditatea. Citologia se ocupă de studiul structurii celulei, iar fiziologia se ocupă de funcționare. Știința care studiază celulele țesuturilor se numește histologie.
Există organisme unicelulare, al căror corp este format în întregime dintr-o celulă. Acest grup include bacterii și protisti (protozoare și alge unicelulare). Uneori sunt numite și acelulare, dar termenul unicelular este folosit mai des. Adevăratele animale multicelulare (Metazoa) și plantele (Metaphyta) conțin multe celule.
Marea majoritate a țesuturilor sunt formate din celule, dar există câteva excepții. Corpul mucegaiurilor slime (mixomicete), de exemplu, este format dintr-o substanță omogenă, necelulară, cu numeroși nuclei. Unele țesuturi animale, în special mușchiul inimii, sunt organizate într-un mod similar. Corpul vegetativ (talul) ciupercilor este format din filamente microscopice – hife, adesea segmentate; fiecare astfel de fir poate fi considerat echivalentul unei cuști, deși de o formă atipică.
Unele structuri ale corpului care nu sunt implicate în metabolism, cum ar fi scoici, perle sau baza minerală a oaselor, sunt formate nu din celule, ci din produsele lor de secreție. Altele, cum ar fi lemnul, scoarța, coarnele, părul și strat exterior piele - nu de origine secretorie, ci formata din celule moarte.
Organismele mici, cum ar fi rotiferele, constau doar din câteva sute de celule. Spre comparație: în corpul uman, există cca. 10 14 celule, în el în fiecare secundă 3 milioane de eritrocite mor și sunt înlocuite cu altele noi, iar aceasta este doar o zece milione din numărul total de celule ale corpului.
De obicei, dimensiunile celulelor vegetale și animale variază între 5 și 20 de microni în diametru. O celulă bacteriană tipică este mult mai mică - aprox. 2 µm, iar cel mai mic cunoscut este 0,2 µm.
Unele celule cu viață liberă, cum ar fi protozoarele, cum ar fi foraminiferele, pot avea câțiva centimetri lungime; au întotdeauna multe nuclee. Celulele fibrelor subțiri de plante ating o lungime de un metru, iar procesele celulelor nervoase ajung la câțiva metri la animalele mari. Cu o astfel de lungime, volumul acestor celule este mic, iar suprafața este foarte mare.
Cele mai mari celule sunt ouă de păsări nefertilizate umplute cu gălbenuș. cel mai mare ou(și, în consecință, cea mai mare celulă) a aparținut unei păsări uriașe dispărute - epiornis ( Aepyornis). Se presupune că gălbenușul său a cântărit aprox. 3,5 kg. Cel mai mare ou din speciile vii aparține struțului, gălbenușul său cântărește cca. 0,5 kg.
De regulă, celulele animalelor și plantelor mari sunt doar puțin mai mari decât celulele organismelor mici. Un elefant este mai mare decât un șoarece, nu pentru că celulele sale sunt mai mari, ci în principal pentru că celulele în sine sunt mult mai mari. Există grupuri de animale, precum rotifere și nematode, în care numărul de celule din organism rămâne constant. Astfel, deși speciile mari de nematozi au un număr mai mare de celule decât cele mici, principala diferență de dimensiune se datorează în acest caz dimensiunii celulelor mari.
Într-un anumit tip de celulă, dimensiunile lor depind de obicei de ploidie, adică. asupra numărului de seturi de cromozomi prezente în nucleu. Celulele tetraploide (cu patru seturi de cromozomi) sunt de 2 ori mai mari ca volum decât celulele diploide (cu un set dublu de cromozomi). Ploidia unei plante poate fi crescută prin injectarea în ea a preparatului pe bază de plante colchicină. Deoarece plantele expuse au celule mai mari, ele sunt și mai mari. Cu toate acestea, acest fenomen poate fi observat doar la poliploidele de origine recentă. La plantele poliploide vechi din punct de vedere evolutiv, dimensiunile celulelor sunt supuse „reglării inverse” către valori normale, în ciuda creșterii numărului de cromozomi.
STRUCTURA UNEI CELULE
La un moment dat, celula a fost considerată ca o picătură mai mult sau mai puțin omogenă de materie organică, care a fost numită protoplasmă sau substanță vie. Acest termen a devenit învechit după ce a devenit clar că celula constă din multe structuri clar separate, numite organele celulare („organe mici”).
Compoziție chimică.
De obicei, 70-80% din masa celulară este apă, în care sunt dizolvate diferite săruri și substanțe cu greutate moleculară mică. compusi organici. Cele mai caracteristice componente ale unei celule sunt proteinele și acizii nucleici. Unele proteine sunt componente structurale ale celulei, altele sunt enzime, de exemplu. catalizatori care determină viteza și direcția fluxului în celule reacții chimice. Acizii nucleici servesc ca purtători informații ereditare, care se realizează în procesul de sinteză intracelulară a proteinelor.
Celulele conțin adesea o anumită cantitate de substanțe de rezervă care servesc drept rezervă alimentară. Celulele vegetale stochează în primul rând amidonul, forma polimerică a carbohidraților. În celulele ficatului și mușchilor, este depozitat un alt polimer carbohidrați, glicogenul. Grăsimea este, de asemenea, printre alimentele stocate în mod obișnuit, deși unele grăsimi îndeplinesc o funcție diferită, și anume, ele servesc ca cele mai importante componente structurale. Proteinele din celule (cu excepția celulelor semințe) nu sunt de obicei stocate.
Nu este posibil să descriem compoziția tipică a unei celule, în primul rând pentru că există diferențe mari în cantitatea de alimente și apă depozitate. Celulele hepatice conțin, de exemplu, 70% apă, 17% proteine, 5% grăsimi, 2% carbohidrați și 0,1% acizi nucleici; restul de 6% sunt săruri și compuși organici cu greutate moleculară mică, în special aminoacizi. Celulele vegetale conțin de obicei mai puține proteine, mult mai mulți carbohidrați și ceva mai multă apă; excepția sunt celulele care se află în stare de repaus. O celulă de repaus a unui bob de grâu, care este o sursă de nutrienți pentru embrion, conține cca. 12% proteine (în principal proteine stocate), 2% grăsimi și 72% carbohidrați. Cantitatea de apă atinge un nivel normal (70–80%) abia la începutul germinării cerealelor.
Principalele părți ale celulei.
Unele celule, mai ales vegetale și bacteriene, au un perete celular exterior. La plantele superioare, constă din celuloză. Peretele înconjoară celula însăși, protejând-o de influențele mecanice. Celulele, în special cele bacteriene, pot secreta și substanțe mucoase, formând astfel o capsulă în jurul lor, care, la fel ca peretele celular, îndeplinește o funcție de protecție.
Cu distrugerea pereților celulari este asociată moartea multor bacterii sub acțiunea penicilinei. Faptul este că în interiorul celulei bacteriene concentrația de săruri și compuși cu molecul scăzut este foarte mare și, prin urmare, în absența unui perete de armare, afluxul de apă în celulă cauzat de presiunea osmotică poate duce la ruperea acesteia. Penicilina, care previne formarea peretelui său în timpul creșterii celulare, duce doar la ruperea (liza) celulei.
Pereții și capsulele celulare nu sunt implicate în metabolism și pot fi adesea detașați fără a ucide celula. Astfel, ele pot fi considerate părți auxiliare externe ale celulei. În celulele animale, pereții celulari și capsulele sunt de obicei absente.
Celula în sine este formată din trei părți principale. Sub perete celular, dacă este prezentă, este membrana celulară. Membrana înconjoară un material heterogen numit citoplasmă. Un nucleu rotund sau oval este scufundat în citoplasmă. Mai jos luăm în considerare mai detaliat structura și funcțiile acestor părți ale celulei.
MEMBRANA CELULARA
Membrana celulară este o parte foarte importantă a celulei. Ține împreună toate componentele celulare și delimitează mediul intern și cel extern. În plus, pliurile modificate ale membranei celulare formează multe dintre organelele celulei.
Membrana celulară este un strat dublu de molecule (strat bimolecular sau dublu). Practic, acestea sunt molecule de fosfolipide și alte substanțe apropiate acestora. Moleculele de lipide au o natură duală, manifestată în modul în care se comportă în raport cu apa. Capetele moleculelor sunt hidrofile, adică. au afinitate pentru apă, iar cozile lor de hidrocarburi sunt hidrofobe. Prin urmare, atunci când sunt amestecate cu apă, lipidele formează o peliculă pe suprafața sa, asemănătoare unei pelicule de ulei; în același timp, toate moleculele lor sunt orientate în același mod: capetele moleculelor sunt în apă, iar cozile de hidrocarburi sunt deasupra suprafeței acesteia.
Există două astfel de straturi în membrana celulară, iar în fiecare dintre ele capetele moleculelor sunt întoarse spre exterior, iar cozile sunt întoarse în interiorul membranei, una la alta, astfel încât să nu atingă apa. Grosimea acestei membrane este de cca. 7 nm. În plus față de principalele componente lipidice, conține molecule mari de proteine care sunt capabile să „plutească” în stratul dublu lipidic și sunt situate astfel încât una dintre părțile lor să fie întoarsă în interiorul celulei, iar cealaltă să fie în contact cu mediul extern. Unele proteine sunt localizate numai pe exteriorul sau doar pe suprafața interioară a membranei sau sunt doar parțial scufundate în stratul dublu lipidic.
Funcția principală a membranei celulare este de a regla transportul de substanțe în și în afara celulei. Deoarece membrana este similară fizic cu uleiul într-o oarecare măsură, substanțele solubile în ulei sau solvenți organici, cum ar fi eterul, trec ușor prin ea. Același lucru este valabil și pentru gaze precum oxigenul și dioxidul de carbon. În același timp, membrana este practic impermeabilă la majoritatea substanțelor solubile în apă, în special la zaharuri și săruri. Datorită acestor proprietăți, este capabil să mențină în interiorul celulei un mediu chimic care diferă de exterior. De exemplu, în sânge, concentrația ionilor de sodiu este mare, iar ionii de potasiu sunt scăzute, în timp ce în lichidul intracelular, acești ioni sunt prezenți în raport opus. O situație similară este tipică pentru mulți alți compuși chimici.
Evident, celula, însă, nu poate fi complet izolată de mediu inconjurator, deoarece trebuie să primească substanțele necesare metabolismului și să scape de produsele sale finale. În plus, stratul dublu lipidic nu este complet impermeabil chiar și pentru substanțele solubile în apă, ci așa-numitele „straturi” care îl pătrund. Proteinele „formatoare de canale” creează pori, sau canale care se pot deschide și închide (în funcție de modificarea conformației proteinei) și în stare deschisă conduc anumiți ioni (Na + , K + , Ca 2+) de-a lungul gradientului de concentrație. În consecință, diferența de concentrații în interiorul celulei și în exterior nu poate fi menținută numai datorită permeabilității scăzute a membranei. De fapt, conține proteine care îndeplinesc funcția de „pompă” moleculară: transportă anumite substanțe atât în celulă, cât și în afara ei, lucrând împotriva gradientului de concentrație. Ca rezultat, atunci când concentrația, de exemplu, de aminoacizi este mare în interiorul celulei și scăzută în exterior, aminoacizii pot fi încă transferați din exterior în interior. Un astfel de transfer se numește transport activ, iar energia furnizată de metabolism este cheltuită pentru el. Pompele cu membrană sunt foarte specifice: fiecare dintre ele este capabilă să transporte fie numai ioni ai unui anumit metal, fie un aminoacid, fie zahăr. Canalele ionice membranare sunt de asemenea specifice.
O astfel de permeabilitate selectivă este foarte importantă din punct de vedere fiziologic, iar absența ei este prima dovadă a morții celulare. Acest lucru poate fi ușor ilustrat cu exemplul sfeclei. Dacă o rădăcină vie de sfeclă este scufundată în apă rece, își păstrează pigmentul; dacă sfecla este fiartă, atunci celulele mor, devin ușor permeabile și pierd pigmentul, care înroșește apa.
Moleculele mari, cum ar fi celulele proteice, pot „înghiți”. Sub influența anumitor proteine, dacă acestea sunt prezente în fluidul din jurul celulei, are loc o invaginare în membrana celulară, care apoi se închide, formând o bulă - o mică vacuolă care conține apă și molecule de proteine; după aceea, membrana din jurul vacuolei se rupe, iar conținutul intră în celulă. Acest proces se numește pinocitoză (literal „băutură celulară”) sau endocitoză.
Particulele mai mari, cum ar fi particulele alimentare, pot fi absorbite într-un mod similar în timpul așa-numitului. fagocitoză. De regulă, vacuola formată în timpul fagocitozei este mai mare, iar hrana este digerată de enzimele lizozomilor din interiorul vacuolei până când membrana care o înconjoară se rupe. Acest tip de nutriție este tipic pentru protozoare, de exemplu, pentru amibe care mănâncă bacterii. Cu toate acestea, capacitatea de fagocitoză este caracteristică atât celulelor intestinale ale animalelor inferioare, cât și fagocitelor, unul dintre tipurile de globule albe (leucocite) ale vertebratelor. În acest din urmă caz, sensul acestui proces nu este în alimentația fagocitelor în sine, ci în distrugerea bacteriilor, virușilor și a altor materiale străine dăunătoare organismului.
Funcțiile vacuolelor pot fi diferite. De exemplu, protozoarele care trăiesc în apa dulce, experimentează un aflux osmotic constant de apă, deoarece concentrația de săruri în interiorul celulei este mult mai mare decât în exterior. Ei sunt capabili să secrete apă într-o vacuolă specială de excreție (contractilă), care își împinge periodic conținutul.
În celulele vegetale, există adesea o vacuola centrală mare care ocupă aproape întreaga celulă; citoplasma formează doar un strat foarte subțire între peretele celular și vacuola. Una dintre funcțiile unei astfel de vacuole este acumularea de apă, care permite celulei să crească rapid în dimensiune. Această abilitate este necesară în special într-un moment în care țesuturile plantelor cresc și formează structuri fibroase.
În țesuturi, în locurile de joncțiune strânsă a celulelor, membranele lor conțin numeroși pori formați din proteine care pătrund în membrană - așa-numitele. conexiuni. Porii celulelor adiacente sunt aranjați unul față de celălalt, astfel încât substanțele cu greutate moleculară mică să se poată muta de la celulă la celulă - acest sistem de comunicare chimică coordonează activitatea lor vitală. Un exemplu de astfel de coordonare este diviziunea mai mult sau mai puțin sincronă a celulelor învecinate observată în multe țesuturi.
CITOPLLASMA
În citoplasmă există membrane interne asemănătoare cu cele exterioare și formând organele de diferite tipuri. Aceste membrane pot fi considerate ca pliuri ale membranei exterioare; uneori membranele interioare formează un întreg integral cu cea exterioară, dar adesea pliul interior este împletit și contactul cu membrana exterioară este întrerupt. Cu toate acestea, chiar dacă contactul este menținut, membranele interioare și exterioare nu sunt întotdeauna identice din punct de vedere chimic. În special, compoziția proteinelor membranare din diferite organele celulare diferă.
Reticulul endoplasmatic.
O rețea de tubuli și vezicule se extinde de la suprafața celulei până la nucleu. Această rețea se numește reticul endoplasmatic. S-a remarcat adesea că tubii se deschid pe suprafața celulei, iar reticulul endoplasmatic joacă astfel rolul unui aparat de microcirculație prin care Mediul extern poate interacționa direct cu tot conținutul celulei. O astfel de interacțiune a fost găsită în unele celule, în special în celulele musculare, dar nu este încă clar dacă este universală. În orice caz, transportul unui număr de substanțe prin acești tubuli de la o parte a celulei la alta are loc efectiv.
Corpuri minuscule numite ribozomi acoperă suprafața reticulului endoplasmatic, în special în apropierea nucleului. Diametrul ribozomului aprox. 15 nm, sunt jumătate proteine, jumătate acizi ribonucleici. Funcția lor principală este sinteza proteinelor; matricea (informații) ARN-ul și aminoacizii asociați cu ARN-ul de transfer sunt atașați de suprafața lor. Zonele reticulului acoperite cu ribozomi se numesc reticul endoplasmatic aspru, iar cele fără ele se numesc netede. Pe lângă ribozomi, de reticulul endoplasmatic sunt adsorbite sau atașate în alt mod diferite enzime, inclusiv sisteme enzimatice care asigură utilizarea oxigenului pentru formarea sterolilor și pentru neutralizarea anumitor otrăvuri. În condiții nefavorabile, reticulul endoplasmatic degenerează rapid și, prin urmare, starea sa servește ca un indicator sensibil al sănătății celulelor.
Aparate Golgi.
Aparatul Golgi (complexul Golgi) este o parte specializată a reticulului endoplasmatic, constând din saci de membrană plate stivuite. Este implicat în secreția de proteine de către celulă (în ea are loc împachetarea proteinelor secretate în granule) și de aceea este dezvoltat în special în celulele care îndeplinesc o funcție secretorie. Funcțiile importante ale aparatului Golgi includ, de asemenea, atașarea grupurilor de carbohidrați la proteine și utilizarea acestor proteine pentru a construi membrana celulară și membrana lizozomului. La unele alge, fibrele de celuloză sunt sintetizate în aparatul Golgi.
Lizozomi
sunt vezicule mici înconjurate de o singură membrană. Ele înmuguresc din aparatul Golgi și posibil din reticulul endoplasmatic. Lizozomii conțin o varietate de enzime care descompun molecule mari, în special proteine. Datorită acțiunii lor distructive, aceste enzime sunt, parcă, „blocate” în lizozomi și sunt eliberate doar la nevoie. Deci, în timpul digestiei intracelulare, enzimele sunt eliberate din lizozomi în vacuolele digestive. Lizozomii sunt, de asemenea, necesari pentru distrugerea celulelor; de exemplu, în timpul transformării unui mormoloc într-o broască adultă, eliberarea enzimelor lizozomale asigură distrugerea celulelor cozii. În acest caz, acest lucru este normal și benefic pentru organism, dar uneori o astfel de distrugere a celulelor este patologică. De exemplu, atunci când praful de azbest este inhalat, acesta poate pătrunde în celulele plămânilor, iar apoi lizozomii se rupe, celulele sunt distruse și se dezvoltă boala pulmonară.
Mitocondriile și cloroplastele.
Mitocondriile sunt formațiuni relativ mari asemănătoare sacului, cu o structură destul de complexă. Ele constau dintr-o matrice înconjurată de o membrană interioară, un spațiu intermembranar și o membrană exterioară. Membrana interioară este pliată în pliuri numite cristae. Acumulările de proteine sunt localizate pe cresta. Multe dintre ele sunt enzime care catalizează oxidarea produselor de descompunere a carbohidraților; altele catalizează reacţiile de sinteză şi oxidare a grăsimilor. Enzimele auxiliare implicate în aceste procese sunt dizolvate în matricea mitocondrială.
În mitocondrii are loc oxidarea substanțelor organice, cuplată cu sinteza adenozin trifosfat (ATP). Descompunerea ATP cu formarea de adenozin difosfat (ADP) este însoțită de eliberarea de energie, care este cheltuită pe diferite procese de viață, de exemplu, pentru sinteza proteinelor și acizilor nucleici, transportul de substanțe în și din exterior. celula, transmiterea impulsurilor nervoase sau contractie musculara. Prin urmare, mitocondriile sunt stații energetice care procesează „combustibil” – grăsimi și carbohidrați – într-o formă de energie care poate fi folosită de celulă și, prin urmare, de organism în ansamblu.
Celulele vegetale conțin și mitocondrii, dar principala sursă de energie pentru celulele lor este lumina. Energia luminoasă este folosită de aceste celule pentru a forma ATP și a sintetiza carbohidrați din dioxid de carbon și apă. Clorofila, un pigment care acumulează energie luminoasă, se găsește în cloroplaste. Cloroplastele, ca și mitocondriile, au o membrană interioară și exterioară. Din excrescențele membranei interioare în procesul de dezvoltare a cloroplastelor, așa-numitele. membrane tilacoide; acestea din urmă formează pungi turtite, adunate în grămezi ca o coloană de monede; aceste stive, numite grana, conțin clorofilă. Pe lângă clorofilă, cloroplastele conțin toate celelalte componente necesare fotosintezei.
Unele cloroplaste specializate nu efectuează fotosinteza, ci îndeplinesc alte funcții, de exemplu, asigură depozitarea amidonului sau a pigmenților.
autonomie relativă.
În unele privințe, mitocondriile și cloroplastele se comportă ca organisme autonome. De exemplu, la fel ca și celulele în sine, care apar numai din celule, mitocondriile și cloroplastele se formează numai din mitocondriile și cloroplastele preexistente. Acest lucru a fost demonstrat în experimente pe celule vegetale, în care formarea de cloroplaste a fost inhibată de antibioticul streptomicina, și pe celulele de drojdie, unde formarea mitocondriilor a fost inhibată de alte medicamente. După asemenea influențe, celulele nu au restaurat niciodată organelele lipsă. Motivul este că mitocondriile și cloroplastele conțin o anumită cantitate din propriul lor material genetic (ADN) care codifică o parte din structura lor. Dacă acest ADN se pierde, ceea ce se întâmplă atunci când formarea de organele este suprimată, atunci structura nu poate fi recreată. Ambele tipuri de organite au propriul sistem de sinteză a proteinelor (ribozomi și ARN-uri de transfer), care este oarecum diferit de sistemul principal de sinteză a proteinelor din celule; se știe, de exemplu, că sistemul de sinteză a proteinelor al organelelor poate fi suprimat de antibiotice, în timp ce acestea nu afectează sistemul principal.
ADN-ul organic este responsabil pentru cea mai mare parte a moștenirii extracromozomiale sau citoplasmatice. Ereditatea extracromozomială nu respectă legile mendeliane, deoarece în timpul diviziunii celulare, ADN-ul organelelor este transmis celulelor fiice într-un mod diferit decât cromozomii. Studiul mutațiilor care apar în ADN-ul organitelor și ADN-ul cromozomilor a arătat că ADN-ul organelelor este responsabil doar pentru o mică parte a structurii organitelor; majoritatea proteinelor lor sunt codificate în gene situate pe cromozomi.
Autonomia genetică parțială a organitelor luate în considerare și caracteristicile sistemelor lor de sinteză a proteinelor au servit drept bază pentru presupunerea că mitocondriile și cloroplastele provin din bacterii simbiotice care s-au instalat în celule în urmă cu 1-2 miliarde de ani. Exemplu modern O astfel de simbioză poate fi servită de mici alge fotosintetice care trăiesc în interiorul celulelor unor corali și moluște. Algele oferă gazdelor lor oxigen și de la ele primesc nutrienți.
structuri fibrilare.
Citoplasma unei celule este un fluid vâscos, astfel încât tensiunea de suprafață s-ar aștepta ca celula să fie sferică, cu excepția cazului în care celulele sunt strâns împachetate. Cu toate acestea, acest lucru nu este de obicei observat. Multe protozoare au tegumente dense sau membrane care dau celulei o formă specifică, nesferică. Cu toate acestea, chiar și fără membrană, celulele pot menține o formă nesferică datorită faptului că citoplasma este structurată cu numeroase fibre paralele, destul de rigide. Acestea din urmă sunt formate din microtubuli goali, care constau din unități proteice organizate în spirală.
Unele protozoare formează pseudopodi - excrescențe citoplasmatice lungi și subțiri cu care captează hrana. Pseudopodiile își păstrează forma datorită rigidității microtubulilor. Dacă presiunea hidrostatică crește la aproximativ 100 de atmosfere, microtubulii se dezintegrează și celula capătă o formă de picătură. Când presiunea revine la normal, microtubulii reasamblați și celula formează pseudopodii. Multe alte celule reacționează în mod similar la modificările presiunii, ceea ce confirmă participarea microtubulilor la menținerea formei celulei. Asamblarea și dezintegrarea microtubulilor, necesare pentru ca celula să își schimbe rapid forma, are loc și în absența modificărilor de presiune.
De asemenea, microtubulii formează structuri fibrilare care servesc ca organe de mișcare a celulelor. Unele celule au excrescențe asemănătoare unor bici numite flageli sau cili - bătaia lor asigură mișcarea celulei în apă. Dacă celula este imobilă, aceste structuri conduc apa, particulele de alimente și alte particule către sau departe de celulă. Flagelii sunt relativ mari și, de obicei, celula are doar unul, uneori mai mulți flageli. Cilii sunt mult mai mici și acoperă întreaga suprafață a celulei. Deși aceste structuri sunt caracteristice în principal protozoarelor, ele pot fi prezente și în forme foarte organizate. În corpul uman, toate căile respiratorii sunt căptușite cu cili. Particulele mici care intră în ele sunt de obicei prinse de mucusul de pe suprafața celulei, iar cilii le mută afară împreună cu mucusul, protejând astfel plămânii. Celulele germinale masculine ale majorității animalelor și ale unor plante inferioare se mișcă cu ajutorul unui flagel.
Există și alte tipuri de mișcare celulară. Una dintre ele este mișcarea amoeboid. Amoeba, precum și unele celule ale organismelor multicelulare, „curg” din loc în loc, adică. se mișcă datorită curentului conținutului celulei. Un curent constant de materie există și în interiorul celulelor vegetale, dar nu implică mișcarea celulei în ansamblu. Cel mai studiat tip de mișcare celulară este contracția celulelor musculare; se realizează prin alunecarea fibrilelor (fire de proteine) unele față de altele, ceea ce duce la scurtarea celulei.
CORE
Nucleul este înconjurat de o membrană dublă. Un spațiu foarte îngust (aproximativ 40 nm) între două membrane se numește perinuclear. Membranele nucleului trec în membranele reticulului endoplasmatic, iar spațiul perinuclear se deschide în reticular. De obicei, membrana nucleară are pori foarte îngusti. Aparent, prin ele sunt transferate molecule mari, cum ar fi ARN-ul mesager, care este sintetizat pe ADN și apoi intră în citoplasmă.
Partea principală a materialului genetic este localizată în cromozomii nucleului celular. Cromozomii constau din lanțuri lungi de ADN dublu catenar, la care sunt atașate proteine de bază (adică, alcaline). Uneori, cromozomii au mai multe fire identice de ADN situate unul lângă celălalt - astfel de cromozomi se numesc politen (multifilamentos). Numărul de cromozomi din diferite specii nu este același. Celulele diploide ale corpului uman conțin 46 de cromozomi sau 23 de perechi.
Într-o celulă care nu se divide, cromozomii sunt atașați în unul sau mai multe puncte de membrana nucleară. În starea normală nespiralizată, cromozomii sunt atât de subțiri încât nu sunt vizibili la microscopul cu lumină. La anumite loci (zone) ale unuia sau mai multor cromozomi se formează un corp dens prezent în nucleele majorității celulelor - așa-numitul. nucleol. În nucleol, ARN-ul este sintetizat și acumulat, care este folosit pentru a construi ribozomi, precum și alte tipuri de ARN.
DIVIZIUNE CELULARA
Deși toate celulele provin din diviziunea celulei precedente, nu toate continuă să se dividă. De exemplu, celulele nervoase din creier, odată ce au apărut, nu se mai divid. Numărul lor scade treptat; țesutul cerebral deteriorat nu se poate recupera prin regenerare. Dacă celulele continuă să se dividă, atunci ele sunt caracterizate de un ciclu celular format din două etape principale: interfaza și mitoza.
Interfaza în sine constă din trei faze: G 1 , S și G 2 . Mai jos este durata lor, tipică pentru celulele vegetale și animale.
G 1 (4–8 ore). Această fază începe imediat după nașterea celulei. În timpul fazei G 1, celula, cu excepția cromozomilor (care nu se modifică), își mărește masa. Dacă celula nu se divide în continuare, rămâne în această fază.
S (6–9 ore). Masa celulei continuă să crească și are loc dublarea (duplicarea) ADN-ului cromozomial. Cu toate acestea, cromozomii rămân unici ca structură, deși au dublat în masă, deoarece cele două copii ale fiecărui cromozom (cromatide) sunt încă conectate între ele pe toată lungimea lor.
G2. Masa celulei continuă să crească până când este de aproximativ dublu față de masa inițială, apoi apare mitoza.
După ce cromozomii s-au dublat, fiecare dintre celulele fiice trebuie să primească un set complet de cromozomi. Simpla diviziune celulară nu poate realiza acest lucru - acest rezultat este obținut printr-un proces numit mitoză. Fără a intra în detalii, începutul acestui proces ar trebui considerat alinierea cromozomilor în planul ecuatorial al celulei. Apoi fiecare cromozom se împarte longitudinal în două cromatide, care încep să diverge în direcții opuse, devenind cromozomi independenți. Ca urmare, la cele două capete ale celulei se află pe întregul set de cromozomi. Apoi celula se împarte în două, iar fiecare celulă fiică primește un set complet de cromozomi.
Următoarea este o descriere a mitozei într-o celulă animală tipică. De obicei, este împărțit în patru etape.
I. Profaza. O structură celulară specială - centriolul - se dublează (uneori această dublare are loc în perioada S a interfazei), iar cei doi centrioli încep să diverge către polii opuși ai nucleului. Membrana nucleară este distrusă; în același timp, proteinele speciale se combină (se agrega), formând microtubuli sub formă de filamente. Centriolii, acum situati la polii opuși ai celulei, au un efect de organizare asupra microtubulilor, care, ca urmare, se aliniază radial, formând o structură care seamănă cu o floare de aster („stea”). Alte fire de microtubuli se întind de la un centriol la altul, formând așa-numitul. ax de diviziune. În acest moment, cromozomii sunt într-o stare spiralizată, asemănătoare cu un izvor. Sunt clar vizibile la microscop optic, mai ales după colorare. În profază, cromozomii se divid, dar cromatidele rămân încă legate în perechi în zona centromerului, un organel cromozomial similar ca funcție cu centriolul. Centromerii au, de asemenea, un efect de organizare asupra firelor fusului, care acum se întind de la centriol la centromer și de la acesta la un alt centriol.
II. Metafaza. Cromozomii, până în acest punct aranjați aleatoriu, încep să se miște, ca și cum ar fi trase de firele fusului atașate de centromerii lor și se aliniază treptat într-un plan într-o anumită poziție și la o distanță egală de ambii poli. Aflat în același plan, centromerii împreună cu cromozomii formează așa-numitul. placa ecuatorială. Centromerii care leagă perechile de cromatide se divid, după care cromozomii surori sunt complet separați.
III. Anafaza. Cromozomii fiecărei perechi se mișcă în direcții opuse spre poli, parcă ar fi târâti de firele fusului. În acest caz, se formează fire între centromerii cromozomilor perechi.
IV. Telofază. De îndată ce cromozomii se apropie de polii opuși, celula însăși începe să se dividă de-a lungul planului în care se afla placa ecuatorială. Ca rezultat, se formează două celule. Fibrele fusului se descompun, cromozomii se desfășoară și devin invizibili, iar în jurul lor se formează o membrană nucleară. Celulele revin la faza G 1 a interfazei. Întregul proces de mitoză durează aproximativ o oră.
Detaliile mitozei variază oarecum în diferite tipuri de celule. Într-o celulă tipică de plantă, se formează un fus, dar nu există centrioli. La ciuperci, mitoza are loc în interiorul nucleului, fără dezintegrarea prealabilă a membranei nucleare.
Diviziunea celulei în sine, numită citokineză, nu este strict legată de mitoză. Uneori una sau mai multe mitoze trec fără diviziune celulară; ca urmare, se formează celule multinucleate, adesea găsite în alge. Dacă nucleul este îndepărtat din oul de arici de mare prin micromanipulare, apoi axul continuă să se formeze și oul continuă să se dividă. Aceasta arată că prezența cromozomilor nu este o condiție necesară pentru diviziunea celulară.
Reproducerea prin mitoză se numește reproducere asexuată, reproducere vegetativă sau clonare. Cele mai multe ale lui aspect important- genetică: la o astfel de reproducere, nu există divergență a factorilor ereditari la descendenți. Celulele fiice rezultate sunt genetic exact aceleași cu cel părinte. Mitoza este singurul mod de auto-reproducere la speciile care nu au reproducere sexuală, cum ar fi multe organisme unicelulare. Cu toate acestea, chiar și la speciile care se reproduc sexual, celulele corpului se divid prin mitoză și provin dintr-o singură celulă, ovulul fertilizat, și, prin urmare, sunt toate identice genetic. Plantele superioare se pot reproduce asexuat (folosind mitoza) prin răsaduri și mustăți (un exemplu celebru este căpșunile).
Reproducerea sexuală a organismelor se realizează cu ajutorul celulelor specializate, așa-numitele. gameți - ovule (ouă) și spermatozoizi (spermatozoizi). Gameții fuzionează pentru a forma o singură celulă, zigotul. Fiecare gamet este haploid, adică. are un set de cromozomi. În cadrul setului, toți cromozomii sunt diferiți, dar fiecărui cromozom al oului îi corespunde unul dintre cromozomii spermatozoizilor. Prin urmare, zigotul conține deja o pereche de astfel de cromozomi corespunzători unul altuia, care se numesc omologi. Cromozomii omologi sunt similari deoarece au aceleasi gene sau variantele lor (alele) care determina caracteristici specifice. De exemplu, unul dintre cromozomii perechi poate avea o genă care codifică pentru grupa sanguină A, iar celălalt o variantă a acesteia care codifică pentru grupa sanguină B. Cromozomii zigot proveniți din ovul sunt materni, iar cei proveniți din spermatozoizi sunt patern.
Ca rezultat al diviziunilor mitotice multiple, fie un organism multicelular, fie numeroase celule cu viață liberă apar din zigotul format, așa cum se întâmplă la protozoarele cu reproducere sexuală și la algele unicelulare.
În timpul formării gameților, setul diploid de cromozomi pe care îl avea zigotul ar trebui redus la jumătate (redus). Dacă acest lucru nu s-ar întâmpla, atunci în fiecare generație fuziunea gameților ar duce la o dublare a setului de cromozomi. Reducerea la numărul haploid de cromozomi are loc ca urmare a diviziunii de reducere - așa-numita. meioza, care este o variantă a mitozei.
scindare si recombinare.
O caracteristică a meiozei este că, în timpul diviziunii celulare, placa ecuatorială este formată din perechi de cromozomi omologi și nu cromozomi individuali dublați, ca în mitoză. Cromozomii perechi, fiecare dintre care au rămas singuri, diverg către polii opuși ai celulei, celula se divide și, ca urmare, celulele fiice primesc o jumătate de set de cromozomi în comparație cu zigotul.
De exemplu, să presupunem că setul haploid este format din doi cromozomi. În zigot (și, în consecință, în toate celulele organismului care produce gameți), sunt prezenți cromozomii materni A și B și paterni A „și B”. În timpul meiozei, ele se pot separa după cum urmează:
Cel mai important lucru din acest exemplu este faptul că atunci când cromozomii diverg, setul matern și patern inițial nu este neapărat format, dar recombinarea genelor este posibilă, ca în gameții AB „și A” B din diagrama de mai sus.
Să presupunem acum că perechea de cromozomi AA” conține două alele - AȘi b- o genă care determină grupele sanguine A și B. În mod similar, o pereche de cromozomi BB „conține alele mȘi n o altă genă care determină grupele sanguine M și N. Separarea acestor alele se poate face după cum urmează:
Evident, gameții rezultați pot conține oricare dintre următoarele combinații de alele a două gene: a.m, bn, bm sau un.
Dacă există mai mulți cromozomi, atunci perechile de alele se vor împărți independent în același mod. Aceasta înseamnă că aceiași zigoți pot produce gameți cu diferite combinații de alele genice și pot da naștere la genotipuri diferite la descendenți.
diviziune meiotică.
Ambele exemple ilustrează principiul meiozei. De fapt, meioza este un proces mult mai complex, deoarece implică două diviziuni succesive. Principalul lucru în meioză este că cromozomii sunt duplicați o singură dată, în timp ce celula se divide de două ori, rezultând o reducere a numărului de cromozomi și setul diploid devine haploid.
În timpul profezei primei diviziuni, cromozomii omologi se conjugă, adică se unesc în perechi. Ca rezultat al acestui proces foarte precis, fiecare genă este opusă omologul său pe celălalt cromozom. Ambii cromozomi se dublează apoi, dar cromatidele rămân conectate între ele printr-un centromer comun.
În metafază, cele patru cromatide unite se aliniază pentru a forma placa ecuatorială, ca și cum ar fi un singur cromozom duplicat. Spre deosebire de ceea ce se întâmplă în timpul mitozei, centromerii nu se divid. Ca rezultat, fiecare celulă fiică primește o pereche de cromatide încă conectate prin cetromer. În timpul celei de-a doua diviziuni, cromozomii, deja individuali, se aliniază din nou, formând, ca în mitoză, o placă ecuatorială, dar dublarea lor nu are loc în timpul acestei diviziuni. Centromerii se divid apoi și fiecare celulă fiică primește o cromatidă.
diviziunea citoplasmei.
Ca urmare a două diviziuni meiotice ale unei celule diploide, se formează patru celule. În timpul formării celulelor germinale masculine, se obțin patru spermatozoizi de aproximativ aceeași dimensiune. În timpul formării ouălor, diviziunea citoplasmei are loc foarte neuniform: o celulă rămâne mare, în timp ce celelalte trei sunt atât de mici încât sunt aproape în întregime ocupate de nucleu. Aceste celule mici, așa-zisul corpii polari, servesc doar pentru a acomoda excesul de cromozomi format ca urmare a meiozei. Partea principală a citoplasmei necesară zigotului rămâne într-o celulă - oul.
conjugarea și trecerea.
În timpul conjugării, cromatidele cromozomilor omologi se pot rupe și apoi se pot alătura într-o nouă ordine, schimbând secțiuni după cum urmează:

Acest schimb de secțiuni de cromozomi omologi se numește crossing over (încrucișare). După cum se arată mai sus, încrucișarea duce la apariția de noi combinații de alele de gene legate. Deci, dacă cromozomii originali ar avea combinații ABȘi ab, apoi după trecerea lor vor conține AbȘi aB. Acest mecanism pentru apariția de noi combinații de gene completează efectul sortării independente a cromozomilor care are loc în timpul meiozei. Diferența este că încrucișarea separă genele de pe același cromozom, în timp ce sortarea independentă separă doar genele de pe cromozomi diferiți.
ALTERNATIE DE GENERATIE
CELULELE PRIMITIVE: PROCARIOTE
Toate cele de mai sus se aplică celulelor plantelor, animalelor, protozoarelor și algelor unicelulare, numite colectiv eucariote. Eucariotele au evoluat dintr-o formă mai simplă, procariotele, care acum sunt bacterii, inclusiv arheobacterii și cianobacterii (cele din urmă erau numite anterior alge albastru-verzi). În comparație cu celulele eucariote, celulele procariote sunt mai mici și au mai puține organele celulare. Au o membrană celulară, dar fără reticul endoplasmatic, iar ribozomii plutesc liber în citoplasmă. Mitocondriile sunt absente, dar enzimele oxidative sunt de obicei atașate de membrana celulară, care devine astfel echivalentul mitocondriilor. Procariotele sunt, de asemenea, lipsite de cloroplaste, iar clorofila, dacă este prezentă, este prezentă sub formă de granule foarte mici.
Procariotele nu au un nucleu închis în membrană, deși locația ADN-ului poate fi identificată prin densitatea sa optică. Echivalentul unui cromozom este o catenă de ADN, de obicei circulară, cu un număr mult mai mic de proteine atașate. Un lanț de ADN la un punct este atașat de membrana celulară. Mitoza este absentă la procariote. Este înlocuit de următorul proces: ADN-ul se dublează, după care membrana celulară începe să crească între punctele de atașare adiacente a două copii ale moleculei de ADN, care ca urmare diverg treptat. Celula se împarte în cele din urmă între punctele de atașare ale moleculelor de ADN, formând două celule, fiecare cu propria copie a ADN-ului.
DIFERENȚIAREA CELULUI
Plantele și animalele pluricelulare au evoluat din organisme unicelulare ale căror celule au rămas împreună după diviziune, formând o colonie. Inițial, toate celulele au fost identice, dar evoluția ulterioară a dat naștere diferențierii. În primul rând, celulele somatice (adică celulele corpului) și celulele germinale s-au diferențiat. În plus, diferențierea a devenit mai complicată - au apărut tot mai multe tipuri de celule diferite. Ontogenia - dezvoltarea individuală a unui organism multicelular - se repetă în in termeni generali acest proces evolutiv (filogeneza).
Fiziologic, celulele se diferențiază parțial prin întărirea uneia sau a alteia caracteristici comune tuturor celulelor. De exemplu, funcția contractilă este crescută în celulele musculare, ceea ce poate fi rezultatul unei îmbunătățiri a mecanismului care efectuează mișcări amiboide sau alte tipuri în celulele mai puțin specializate. Un exemplu similar sunt celulele rădăcinii cu pereți subțiri cu procesele lor, așa-numitele. firele de păr de rădăcină, care servesc la absorbția sărurilor și a apei; într-o măsură sau alta, această funcție este inerentă oricăror celule. Uneori, specializarea este asociată cu dobândirea de noi structuri și funcții - un exemplu este dezvoltarea unui organ locomotor (flagel) în spermatozoizi.
Diferențierea la nivel celular sau tisular a fost studiată în detaliu. Știm, de exemplu, că uneori se procedează autonom, adică. un tip de celulă se poate transforma în altul, indiferent de tipul de celule din care aparțin vecinii. Cu toate acestea, așa-numitul. inducția embrionară este un fenomen în care un tip de țesut stimulează celulele de alt tip să se diferențieze într-o direcție dată.
În cazul general, diferențierea este ireversibilă, adică. celulele foarte diferențiate nu se pot transforma într-un alt tip de celulă. Cu toate acestea, acest lucru nu este întotdeauna cazul, mai ales în celulele vegetale.
Diferențele de structură și funcție sunt determinate în cele din urmă de ce tipuri de proteine sunt sintetizate în celulă. Deoarece sinteza proteinelor este controlată de gene, iar setul de gene din toate celulele corpului este același, diferențierea ar trebui să depindă de activarea sau inactivarea anumitor gene în tipuri variate celule. Reglarea activității genelor are loc la nivelul transcripției, adică. formarea ARN mesager folosind ADN-ul ca șablon. Numai genele transcrise produc proteine. Proteinele sintetizate pot bloca transcripția, dar uneori o activează. De asemenea, deoarece proteinele sunt produse ale genelor, unele gene pot controla transcrierea altor gene. Hormonii, în special hormonii steroizi, sunt, de asemenea, implicați în reglarea transcripției. Genele foarte active pot fi duplicate (dublate) de multe ori pentru a produce mai mult ARN mesager.
Dezvoltarea tumorilor maligne a fost adesea privită ca un caz special de diferențiere celulară. Cu toate acestea, apariția celulelor maligne este rezultatul unei modificări în structura ADN-ului (mutație), și nu al proceselor de transcripție și translație în proteina ADN-ului normal.
METODE DE STUDIU CELULUI
Microscop cu lumină.
În studiul formei și structurii celulelor, primul instrument a fost microscopul cu lumină. Rezoluția sa este limitată de dimensiuni comparabile cu lungimea de undă a luminii (0,4–0,7 µm pentru lumina vizibila). Cu toate acestea, multe elemente ale structurii celulare au dimensiuni mult mai mici.
O altă dificultate este că majoritatea componentelor celulare sunt transparente și indicele lor de refracție este aproape același cu cel al apei. Pentru a îmbunătăți vizibilitatea, sunt adesea folosiți coloranți care au afinități diferite pentru diferite componente celulare. Colorarea este, de asemenea, folosită pentru a studia chimia celulei. De exemplu, unii coloranți se leagă predominant la acizii nucleici și, prin urmare, dezvăluie localizarea lor în celulă. O mică parte din coloranți - se numesc intravitali - poate fi folosită pentru colorarea celulelor vii, dar de obicei celulele trebuie să fie prefixate (folosind substanțe care coagulează proteina) și abia apoi pot fi colorate. Cm. HISTOLOGIE.
Înainte de testare, celulele sau bucățile de țesut sunt de obicei încorporate în parafină sau plastic și apoi tăiate în secțiuni foarte subțiri folosind un microtom. Această metodă este utilizată pe scară largă în laboratoarele clinice pentru a detecta celulele tumorale. Pe lângă microscopia luminoasă convențională, au fost dezvoltate și alte metode optice pentru studiul celulelor: microscopia cu fluorescență, microscopia cu contrast de fază, spectroscopie și analiza prin difracție de raze X.
Microscop electronic.
Microscopul electronic are o rezoluție de cca. 1–2 nm. Acest lucru este suficient pentru studiul moleculelor mari de proteine. De obicei, este necesar să colorați și să contrastați obiectul cu săruri sau metale metalice. Din acest motiv, precum și pentru că obiectele sunt examinate în vid, doar celulele moarte pot fi studiate cu un microscop electronic.
Autoradiografie.
Dacă în mediu se adaugă un izotop radioactiv absorbit de celule în timpul metabolismului, atunci localizarea sa intracelulară poate fi detectată folosind autoradiografie. În această metodă, secțiunile subțiri de celule sunt plasate pe film. Filmul se întunecă sub acele locuri în care există izotopi radioactivi.
Centrifugarea.
Pentru studiul biochimic al componentelor celulare, celulele trebuie distruse - mecanic, chimic sau prin ultrasunete. Componentele eliberate sunt în suspensie în lichid și pot fi izolate și purificate prin centrifugare (cel mai adesea într-un gradient de densitate). De obicei, astfel de componente purificate păstrează o activitate biochimică ridicată.
Culturi celulare.
Unele țesuturi pot fi împărțite în celule individuale, astfel încât celulele să rămână în viață și să fie adesea capabile să se reproducă. Acest fapt confirmă în cele din urmă ideea unei celule ca unitate a vieții. Un burete, un organism multicelular primitiv, poate fi împărțit în celule prin frecarea printr-o sită. După un timp, aceste celule se recombină și formează un burete. Țesuturile embrionare animale pot fi făcute să se disocieze folosind enzime sau alte mijloace care slăbesc legăturile dintre celule.
Embriologul american R. Harrison (1879–1959) a fost primul care a arătat că celulele embrionare și chiar unele mature pot crește și se pot multiplica în afara corpului într-un mediu adecvat. Această tehnică, numită cultură celulară, a fost perfecționată de biologul francez A. Carrel (1873-1959). Celulele vegetale pot fi cultivate și în cultură, dar, în comparație cu celulele animale, ele formează grupuri mai mari și sunt mai puternic atașate unele de altele, astfel încât țesutul se formează în timpul creșterii culturii, mai degrabă decât celulele individuale. În cultura celulară, o plantă adultă întreagă, cum ar fi un morcov, poate fi cultivată dintr-o singură celulă.
Microchirurgie.
Cu ajutorul unui micromanipulator, părți individuale ale celulei pot fi îndepărtate, adăugate sau modificate într-un fel. O celulă mare de amibe poate fi împărțită în trei componente principale - membrana celulară, citoplasmă și nucleu, iar apoi aceste componente pot fi reasamblate și se obține o celulă vie. În acest fel, se pot obține celule artificiale, formate din componente ale diferitelor tipuri de amibe.
Având în vedere că este posibilă sintetizarea artificială a unor componente celulare, experimentele privind asamblarea celulelor artificiale pot fi primul pas către crearea de noi forme de viață în laborator. Întrucât fiecare organism se dezvoltă dintr-o singură celulă, metoda de obținere a celulelor artificiale permite în principiu construcția unor organisme de un anumit tip, dacă în același timp se utilizează componente ușor diferite de cele găsite în celulele existente în prezent. În realitate, totuși, nu este necesară sinteza completă a tuturor componentelor celulare. Structura majorității, dacă nu a tuturor componentelor unei celule este determinată de acizii nucleici. Astfel, problema creării de noi organisme se reduce la sinteza de noi tipuri de acizi nucleici și înlocuirea acestora cu acizi nucleici naturali în anumite celule.
Fuziunea celulară.
Un alt tip de celule artificiale poate fi obținut prin fuziunea celulelor de același tip sau de diferite tipuri. Pentru a realiza fuziunea, celulele sunt expuse la enzime virale; în acest caz, suprafețele exterioare ale două celule se lipesc împreună, iar membrana dintre ele se prăbușește și se formează o celulă în care două seturi de cromozomi sunt închise într-un nucleu. Puteți îmbina celule de diferite tipuri sau în diferite stadii de diviziune. Folosind această metodă, a fost posibil să se obțină celule hibride de șoarece și pui, om și șoarece, om și broască râioasă. Astfel de celule sunt hibride doar inițial și după numeroase diviziunile celulare pierde majoritatea cromozomilor fie ai unei specii, fie a celeilalte specii. Produsul final devine, de exemplu, în esență o celulă de șoarece, în care genele umane sunt absente sau prezente doar în cantități mici. Un interes deosebit este fuziunea celulelor normale și maligne. În unele cazuri, hibrizii devin maligni, în altele nu; ambele proprietăți pot apărea atât ca dominante, cât și ca recesive. Acest rezultat nu este neașteptat, deoarece malignitatea poate fi cauzată de diverși factori și are un mecanism complex.
Literatură:
Ham A, Cormac D. Histologie, vol. 1. M., 1982
Alberts B., Bray D., Lewis J., Raff M., Roberts K., Watson J. Biologia moleculară a celulei, vol. 1. M., 1994
1. Structura similară a celulelor vegetale și animale - dovadă
a) relația lor
B) origine comună a organismelor din toate regnurile
c) originea plantelor din animale
D) complicarea organismelor în procesul de evoluție
D) unitatea lumii organice
E) diversitatea organismelor
Răspuns
2. Ce funcții îndeplinește complexul Golgi?
A) sintetizează substanţe organice din anorganice
B) descompune biopolimerii în monomeri
B) acumulează proteine, lipide, carbohidrați sintetizati în celulă
D) asigură ambalarea și îndepărtarea substanțelor din celulă
D) oxidează substanțele organice la anorganice
E) participă la formarea lizozomilor
Răspuns
3. Stabiliți o corespondență între trăsătura organismului și grupul pentru care este caracteristic: 1-procariote, 2-virusuri.
A) structura celulară a corpului
B) prezența propriului metabolism
C) inserarea propriului ADN în ADN-ul celulei gazdă
D) constă din acid nucleicși învelișul proteic
D) reproducerea prin împărțire în două
E) capacitatea de a inversa transcrierea
Răspuns
A1 B1 C2 D2 E1 E2
4. Autotrofii includ
A) plante cu spori
B) ciuperci
B) alge unicelulare
D) bacterii chimiotrofe
d) virusuri
E) majoritatea protozoarelor
Răspuns
5. Setați secvența proceselor care au loc în timpul meiozei
A) localizarea perechilor de cromozomi omologi în planul ecuatorial
B) conjugarea, încrucișarea cromozomilor omologi
C) localizarea în planul ecuatorului și divergența cromozomilor surori
D) formarea a patru nuclei haploizi
D) divergenţa cromozomilor omologi
Răspuns
5a. Care sunt caracteristicile meiozei?
A) prezența a două diviziuni consecutive
B) formarea a două celule cu aceleași informații ereditare
C) divergența cromozomilor omologi în diferite celule
D) formarea celulelor fiice diploide
E) lipsa interfazei înainte de prima divizie
E) conjugarea și încrucișarea cromozomilor
Răspuns
6. Stabiliți o corespondență între caracteristicile gametogenezei și tipul acesteia: 1-ovogeneză, 2-spermatogeneză
A) se formează o celulă sexuală mare
B) se formează celule de ghidare
C) se formează mulți gameți mici
D) nutrienții sunt stocați într-una dintre cele patru celule
D) se formează gameți mobili
Răspuns
A1 B1 C2 D1 D2
7. Stabiliți succesiunea proceselor care au loc într-o celulă cu cromozomi în interfază și mitoză ulterioară
A) localizarea cromozomilor în planul ecuatorial
B) Replicarea ADN-ului și formarea cromozomilor cu două cromatide
B) spiralizarea cromozomilor
D) divergența cromozomilor surori la polii celulei
Răspuns
7+. Ce structuri celulare suferă cele mai mari modificări în timpul mitozei?
A) miezul
B) citoplasmă
B) ribozomi
D) lizozomi
D) centrul celular
E) cromozomi
Răspuns
8. Stabiliți o corespondență între structura organoidului și tipul acestuia: 1-centru celular, 2-ribozom
A) este format din doi cilindri dispuși perpendicular
B) este format din două subunități
B) formată din microtubuli
D) contine proteine care asigura miscarea cromozomilor
D) conține proteine și acid nucleic
Răspuns
A1 B2 C1 D1 D2
9. Setați secvența proceselor care au loc în timpul fagocitozei
A) intrarea monomerilor în citoplasmă
B) captarea nutrienților de către membrana celulară
C) hidroliza polimerilor la monomeri
D) formarea unei vezicule fagocitare în interiorul celulei
D) fuziunea veziculei fagocitare cu lizozomul
Răspuns
Răspuns
12. Principalele prevederi ale teoriei celulare ne permit să tragem concluzii despre
A) influența mediului asupra fitnessului
B) relația dintre organismele
D) dezvoltarea organismelor de la simplu la complex
E) posibilitatea generării spontane a vieții din materie neînsuflețită
Răspuns
12+. Principalele prevederi ale teoriei celulare ne permit să tragem concluzii despre
A) migrarea biogenă a atomilor
B) relația dintre organismele
C) originea plantelor și animalelor dintr-un strămoș comun
D) apariția vieții pe Pământ în urmă cu aproximativ 4,5 miliarde de ani
D) structura similară a celulelor tuturor organismelor
E) relația dintre natura animată și cea neînsuflețită
Răspuns
12++. Care sunt prevederile teoriei celulare?
A) se formează celule noi ca urmare a diviziunii celulei mamă
B) Celulele sexuale conțin un set haploid de cromozomi
C) Celulele sunt similare din punct de vedere chimic
D) Celulă - o unitate de dezvoltare a tuturor organismelor
D) Celulele țesuturilor tuturor plantelor și animalelor au aceeași structură.
E) Toate celulele conțin molecule de ADN.
Răspuns
13. Asemănarea celulelor fungice și animale este că au
A) o înveliș de substanță asemănătoare chitinei
B) glicogenul ca carbohidrat de stocare
B) miez în formă
D) vacuole cu seva celulară
d) mitocondriile
E) plastide
A) formează țesuturi și organe
b) participa la procesul de fertilizare
B) întotdeauna haploid
D) au un set diploid de cromozomi
D) se formează în timpul meiozei
E) se împarte prin mitoză
Răspuns
A1 B2 C2 D1 E2 E1
17. Prin ce sunt diferite mitocondriile de lizozomi?
A) au membrane exterioare și interioare
B) au numeroase excrescente - cristae
C) participa la procesele de eliberare a energiei
D) în ele acidul piruvic este oxidat în dioxid de carbon și apă
D) în ele, biopolimerii sunt descompuși în monomeri
E) sunt implicate în metabolism
Răspuns