Какие женские гормоны выделяются гипоталамусом. Гормоны гипоталамуса статины, либерины и гипофиза тропные
Читайте также
Важнейшая роль в регуляции гормональных функций гипофиза и периферических эндокринных желез принадлежит гипоталамусу.
Это своеобразный нейроэндокринный орган, обеспечивающий единство нервных и эндокринных процессов в организме. Гипоталамус имеет нервные связи с лимбической системой, ретикулярной формацией, корой головного мозга, гипофизом. Через гипофиз он связан с другими эндокринными железами. Нервные клетки гипоталамуса образуют нейрогормоны, или рилизинг-гормоны (рилизинг-факторы).
Рилизинг-гормоны гипоталамуса, регулирующие тропные функции аденогипофиза, по своей химической природе относятся к полипептидам. Местом их выработки являются соответствующие ядра гипоталамуса. К рилизинг-гормонам гипоталамуса относятся соматолиберин и соматостатин, пролактолиберин и пролактостатин, люлиберин, фоллиберин, кортиколиберин, тиролиберин, меланолиберин, меланостатин и др. Перечисленные рилизинг-гормоны имеют высокую биологическую активность и лишены видовой специфичности. Многие из них (ТРГ, Гн-РГ, ГР-РГ) синтезированы химическим путем. Механизм, посредством которого рилизинг-гормоны гипоталамуса стимулируют или тормозят инкрецию гормонов аденогипофиза, связан с изменением проницаемости клеточных мембран, однако пока еще достаточно не изучен. Полагают, что они ускоряют освобождение гормонов гипофиза и участвуют в их биосинтезе.
В связи с высокой устойчивостью к действию протеолитических ферментов, высокой активностью и отсутствием видовой специфичности рилизинг-гормоны, очевидно, получат широкое применение в животноводстве и ветеринарии.
Соматолиберин , или рилизинг-гормон, стимулирующий гормон роста (ГР-РГ), и соматостатин , или рилизинг-гормон, ингибирующий гормон роста (ГР-ИГ), вырабатываются в основном в области срединного возвышения и медиобазальной части гипоталамуса. Синтезированы аналоги ГР-РГ и ГР-ИГ. Действие рилизинг-гормонов на соматотропные клетки гипофиза осуществляется с участием цАМФ и цГМФ. Изменение соотношения цАМФ и цГМФ в клетках гипофиза обеспечивает регуляцию инкреции гормона роста.
Пролактолиберин , или пролактинстимулирующий гормон (ПСГ), и пролактостатин , или пролактинингибирующий гормон (ПИГ), вырабатываются в медиальной преоптической и некоторых других областях гипоталамуса. Эти рилизинг-гормоны соответственно стимулируют и тормозят иикрецию пролактина аденогипофизом. Физиологическими стимуляторами инкреции пролактина являются акт сосания и изменение концентрации эстрогенов в крови. Основным стимулятором инкреции ПИГ является дофамин.
Гонадолиберин , или гонадотропин-рилизинг-гормон (Гн-РГ), содержит в себе люлиберин и фоллиберин. Он образуется, в основном, медиобазальной частью гипоталамуса. В 1971 г. осуществлен синтез Гн-РГ. Этот гормон регулирует инкрецию лютеинизирующего гормона (ЛГ) и фолликулостимулирующего гормона (ФСГ) в гипофизе. Полагают, что выделение ЛГ или ФСГ обусловлено влиянием половых стероидов на гипоталамус и гипофиз. Повышение уровня эстрогенов в крови повышает чувствительность гипофиза к Гн-РГ и сопровождается освобождением ЛГ. Стимуляция выделения ЛГ осуществляется эстрогенами в меньших концентрациях и проявляется раньше, чем стимуляция выделения ФСГ.
В настоящее время существуют две гипотезы, объясняющие механизм действия Гн-РГ. Согласно первой Гн-РГ изменяет проницаемость клеточных мембран гонадотропов, вызывает их деполяризацию и повышает концентрацию ионов кальция, которые активируют выход гонадотропинов. Вторая гипотеза придает главное значение Гн-РГ в активации аденилциклазной системы и увеличении концентрации цАМФ в цитозоле клеток аденогипофиза. Допускается возможность объединения этих гипотез.
Кортиколиберин , или кортикотропин-рилизинг-гормон (КРГ),- гормон, стимулирующий инкрецию адренокортикотропного гормона (АКТГ) гипофизом. Основным местом синтеза КРГ является медиобазальная область гипоталамуса. КРГ выделяется из гипоталамуса с определенной цикличностью путем периодических выбросов. Механизм действия КРГ на клетки гипофиза, инкретирующие АКТГ, изучен недостаточно.
Тиролиберин , или тиротропин-рилизинг-гормон (ТРГ), - выделен из гипоталамуса овец, свиней. Для выделения одного миллиграмма чистого ТРГ потребовалось 300 000 гипоталамусов овец. ТРГ имеет одинаковую структуру у всех животных. Местом образования ТРГ является преоптическая область гипоталамуса.
ТРГ и тиреоидные гормоны, поступающие в кровь, регулируют тиротропную функцию гипофиза. ТРГ действует на рецепторы мембран тиротропных клеток при определенной концентрации Са ++ и Na + в экстрацеллюлярной жидкости.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
И гипоталамуса оказывают исключительное влияние на весь организм человека. Они координируют рост, развитие, половое созревание и все виды обмена веществ. Гормоны гипоталамуса, выделение которых контролируется гипофизом, регулируют множество жизненно важных функций организма. Давайте рассмотрим эту железу с анатомической точки зрения.
Гормоны гипоталамуса и его строение
Гипофиз, центральный орган представляет собой небольшое округлое образование, состоящее из двух частей. Гипоталамус же расположен над гипофизом в так называемом Его еще называют подбугорьем. Вес железы - до пяти граммов. Однако это маленькое образование играет значительную роль для нашего тела, регулируя температурный баланс, обмен веществ (как белков, жиров и углеводов, так и минералов), функции щитовидки, яичников и надпочечников. Состоит железа из трех отделов, имеет гипофизарную ножку. Основную ее массу составляют нейросекреторные и группирующиеся в ядра (которых более 30).
Рилизинг-гормоны
Кортиколиберин воздействует на переднюю часть гипофиза. Этот нейропептид регулирует ряд (реакции активизации, способность к ориентировке). Этот гормон усиливает тревогу, страх, напряжение. Длительное его воздействие на организм приводит к хроническому стрессу, депрессии, истощению, бессоннице. Такие гормоны гипоталамуса, как упомянутый кортиколиберин, являются веществами пептидной природы. Это части белковых молекул. Всего нейрогормонов 7, их называют еще либеринами. Их воздействие на гипофиз дает начало синтезу тропных гормонов - соматотропина, гонадотропина и тиреотропина. Помимо них нейросекреторные клетки, находящиеся в гипоталамусе, вырабатывают и другие вещества, влияющие на гипофиз. Это - статины, угнетающие секрецию перечисленных тропных гормонов. Все они влияют на рост, развитие, взаимодействие эндокринной системы с нервной. Стимуляторами рилизинг-гормонов могут являться катехоламины. Однако это пока только гипотеза.

Окситоцин
Синтезируясь в гипоталамусе, это вещество затем поступает в гипофиз (заднюю его долю) и выделяется в кровь. Максимальная концентрация окситоцина связана с ощущением эмоциональной близости - у матерей при контакте с новорожденным ребенком, у мужчин при привязанности и сексуальных контактах. Если этот гормон вырабатывается в недостаточном количестве, тогда невозможна оптимальная родовая деятельность, высока опасность выкидыша.
Вазопрессин
Невозможно перечислять гормоны гипоталамуса и не упомянуть Его функциями являются повышение артериального давления, поддержание водного баланса, координация усвоения калия в организме. Секреция вазопрессина увеличивается при тошноте, стрессе, боли, гипогликемии. Чтобы снизить ее, следует употреблять много (курага, помидоры). Недостаток вазопрессина приводит к развитию несахарного диабета.
Препараты гормонов гипоталамуса
Лекарственные средства «Гонадорелин» и «Леупролид» используют в терапии задержки полового созревания, при крипторхизме и гипогонадизме. А также при поликистозе яичников, эндометриозе.
Рилизинг-гормоны являются нейрогормонами человека, которые синтезируют ядра гипоталамуса. Они угнетают (статины) или стимулируют (либерины) выработку тропных гормонов гипофиза. Активизируется работа желез внутренней секреции, и происходит регуляция выделения ими гормонов. Высшие отделы ЦНС и находятся в тесной связи из-за рилизинг-гормонов.
Функции гипоталамуса
Одно из важных составляющих эндокринной системы, отвечающее за производство гормонов, - гипоталамус. Вырабатываемые гипоталамусом вещества представляют собой гормоны, участвующие в метаболических процессах организма.
В гипоталамусе имеются нервные клетки, обеспечивающие выработку необходимых веществ, нужных организму для нормального функционирования. Эти клетки именуются нейросекреторными. Их задача - прием импульсов, которые передают разные участки нервной системы. Выделение элементов происходит через аксовазальные синапсы.
Продуцируемые гипоталамусом рилизинг-гормоны или, как их по-другому называют, статины и либерины, крайне необходимы для нормальной работы гипофиза. По своей химической природе они являются пептидами. Благодаря химическим и нервным импульсам происходит их синтез, транспортируются они в гипофиз кровью по гипоталамо-гипофизарной системе.
Классификация гормонов
Рассмотрим наиболее известные рилизинг-гормоны:
- Ингибирующие секреторную - речь идет о соматостатине, меланостатине, пролактостатине.
- Стимулирующие - речь идет о меланолиберине, пролактолиберине, фоллиберине, люлиберине, соматолиберине, тиролиберине, гонадолиберине и кортиколиберине.
Перечисленные вещества, точнее некоторые из них, могут быть спродуцированы другими органами, не только гипоталамусом (например, поджелудочной железой).
Статины и либерины
Функционирование гипофиза напрямую зависит от них. Также они оказывают влияние на функционирование периферических эндокринных желез:
- щитовидной железы;
- яичников у девушек;
- яичек у мужского пола.

Статины и либерины, которые наиболее известны:
- дофамин;
- гонадолиберин (люлиберин, фоллиберин);
- мелоностатин;
- соматостатин;
- тиреолиберин.
Секрецию лютеинизирующего и фолликулостимулирующего гормонов гипофиза обеспечивают гонадолиберины.
На активность андрогенов у мужчин также влияют гонадолиберины, способствуют увеличению активности сперматозоидов и уровню либидо.
А у женщин нейрогормоны отвечают за менструальный цикл, причем количество гормонов меняется в зависимости он фазы цикла.
Недостаточная выработка рилизинг-гормонов нередко становится причиной бесплодия и импотенции.
Характеристика гормонов
Гормон кортиколиберин, отвечающий за чувство тревоги, вырабатывается гипоталамусом. Это еще один важный рилизинг-фактор, действующий совместно с гормонами гипофиза и оказывающий воздействие на работу надпочечников. Люди с недостатком этого гормона часто страдают гипертонией и надпочечниковой недостаточностью.
Гонадолиберин - гормон, усиливающий выработку гонадотропинов, - также является продуктом гипоталамуса. Его еще называют гонадотропин-рилизинг-гормон.
Нормальное функционирование половых органов не может обходиться без гонадолиберина. Именно этот гормон отвечает за естественное протекание менструального цикла у женщин. С его участием проходит процесс созревания и выхода яйцеклетки. Этот гормон отвечает за либидо (половое влечение). При недостаточном производстве гипоталамусом этого гормона у женщин часто развивается бесплодие. Какие еще бывают рилизинг-гормоны?

Соматолиберин
Наиболее бурно выделяется в детском и подростковом возрасте. Его основным свойством является нормализация процессов роста органов и систем организма. От его выработки зависит полноценное развитие и формирование ребенка. Недостаточное производство этого гормона гипоталамуса может привести к
Пролактолиберин
Его выработка наиболее активно происходит в период вынашивания плода и на протяжении всего периода вскармливания ребенка матерью. Этот рилизинг-фактор нормализует выработку пролактина, формирующего протоки молочных желез.
Пролактостатин
Пролактостатин относится к подклассу статинов, вырабатываемых гипоталамусом, отвечает за угнетение пролактина.
К пролактостатинам относятся:
· дофамин;
· соматостатин;
· меланостатин.
Их основное действие направлено на подавление тропных гормонов гипофиза и гипоталамуса.
Меланотропин-рилизинг-гормон
На процесс выработки меланина и разделение пигментных клеток оказывает влияние меланолиберин. Также он воздействует на элементы ПРД гипофиза.
Оказывает влияние на нейрофизиологическое поведение человека. Применяется для снятия депрессивных состояний и лечения паркинсонизма.
Тиреотропин-рилизинг-гормон (ТРГ)
К тиреотропин-рилизинг-гормонам гипоталамуса также относится тиролиберин. Он способствует продуцированию тиреотропных гормонов аденогипофиза.
В незначительной степени оказывает воздействие на процесс выработки пролактина. Тиролиберин обеспечивает повышение концентрации тироксина в крови.
ЦНС оказывает большое влияние на процессы продуцирования гормонов. За выработку нейрогормонов отвечают нейросекреторные клетки регуляционной системы.
Основные функции либеринов

Это рилизинг-гормоны гипоталамуса. Выполняют Гонадолиберины нормализуют функционирование половой сферы женщин и мужчин.
Они отвечают за воспроизводство фолликулостимулирующих гормонов и оказывают воздействие на работу яичек и яичников.
Такой компонент, как люлиберин, оказывает обособляющее действие на овуляцию, формируя возможность зачатия плода.
У женщин, равнодушных к интимной жизни, люлиберин и фоллиберин вырабатываются в недостаточном количестве.
Существуют еще рилизинг-факторы, относящиеся к средней доле гипоталамуса, но их соединения с элементами гипофиза и аденогипофиза не изучены.
Агонисты рилизинг-гормона: препараты
Как уже было отмечено, данные гормоны вырабатывает гипоталамус. Когда требуется простимулировать яичники, к примеру перед процедурой ЭКО, применяют агонисты или аналоги рилизинг-гормонов. То есть они оказывают на организм то же действие, что и собственный гормон.
Но велика вероятность развития побочных реакций со стороны женского организма. Так происходит по причине снижения уровня эстрогенов. К наиболее частым явлениям относят:
- головную боль;
- излишнюю потливость;
- приливы;
- сухость во влагалище;
- перепады настроения;
- депрессивные состояния.
Препараты применяются следующие:


Нами рассмотрены препараты-агонисты рилизинг-гормона.
Антагонисты
Поскольку при приеме агонистов рилизинг-гормона чрезвычайно повышается эстрадиол, может произойти резкий выброс лютеинизирующего гормона. Это приводит к преждевременной овуляции и гибели яйцеклеток. Чтобы этого не произошло, применяют антагонисты рилизинг-гормона. В результате их действия гипофиз снова можно стимулировать. не проявляет себя, а ведь так часто случалось из-за длительного использования агонистов ГнРг. Вводят через пять дней с момента начала применения фолликулостимулирующего гормона.
Чтобы терапия проходила успешно, все назначения препаратов должен проводить только специалист.
Гипоталамус служит местом непосредственного взаимодействия высших отделов ЦНС и эндокринной системы. Природа связей, существующих между ЦНС и эндокринной системой, стала проясняться в последние десятилетия, когда из гипоталамуса были выделены первые гуморальные факторы, оказавшиеся гормональными веществами с чрезвычайно высокой биологической активностью. Потребовалось немало труда и экспериментального мастерства, чтобы доказать, что эти вещества образуются в нервных клетках гипоталамуса, откуда по системе портальных капилляров достигают гипофиза и регулируют секрецию гипофизарных гормонов, точнее их освобождение. Эти вещества получили сначала наименование нейрогормонов, а затем рилизинг-факторов (от англ.release – освобождать), или либеринов . Вещества с противоположным действием, т.е. угнетающие освобождение гипофизарных гормонов, стали называть ингибирующими факторами, или статинами . Таким образом, гормонам гипоталамуса принадлежит ключевая роль в физиологической системе гормональной регуляции многосторонних биологических функций отдельных органов, тканей и целостного организма.
Анатомическое строение
Гипоталамус - главный подкорковый центр регуляции вегетативно-висцеральных и эндокринных функций.
Он занимает вентральную часть промежуточного мозга и располагается ниже подбугорной борозды, sulcus hypothalamicus . Делится на зрительную и обонятельную части, pars optica et pars olfactoria .
В настоящее время в гипоталамусе описано 32-48 ядер, являющимися высшими вегетативными центрами, регулирующими все виды обмена веществ, терморегуляцию и т. д.
- Различают три основные области скопления групп нервных клеток гипоталамуса:
- переднюю;
- промежуточную;
- заднюю.
Некоторые ядра обладают нейросекреторной функцией (супраоптические, паравентрикулярные, дугообразные и вентромедиальные).
Гистологическое строение
Гипоталамус - участок промежуточного мозга, содержащий особые нейросекреторные ядра , клетки которых вырабатывают и секретируют в кровь нейрогормоны . Эти клетки получают афферентные импульсы из других частей нервной системы, а их аксоны оканчиваются на кровеносных сосудах (аксо-вазальные синапсы) .
Нейросекреторные клетки - отростчатой формы, с крупным везикулярным ядром, хорошо заметным ядрышком и базофильной цитоплазмой, содержащей развитую грЭПС и крупный комплекс Гольджи, от которого отделяются нейросекреторные гранулы (рис. 1). Гранулы транспортируются со скоростью около 1-4 мм/ч по аксону вдоль центрального пучка микротрубочек и микрофиламентов, а местами накапливаются в больших количествах, растягивая аксон. Самые крупные из таких участков хорошо видны под световым микроскопом и называются накопительными нейросекреторными тельцами (Херринга) ; в них сосредоточено до 60% всего нейросекрета, лишь около 30% находится в области терминалей . Терминали (аксо-вазальные синапсы) характеризуются присутствием, помимо гранул, многочисленных светлых пузырьков (осуществляют возврат мембраны после экзоцитоза).
Рис. 1. Ультраструктурная организация нейросекреторных клеток. А - аксон, ABC - аксо-вазапьный синапс, КАП - капилляр, ННСТ - накопительное нейросекреторное тельце, НСГ - нейросекреторные гранулы.
Нейросекреторные ядра гипоталамуса в зависимости от размеров клеток и их функциональных особенностей разделяют на крупно- и мелкоклеточные .
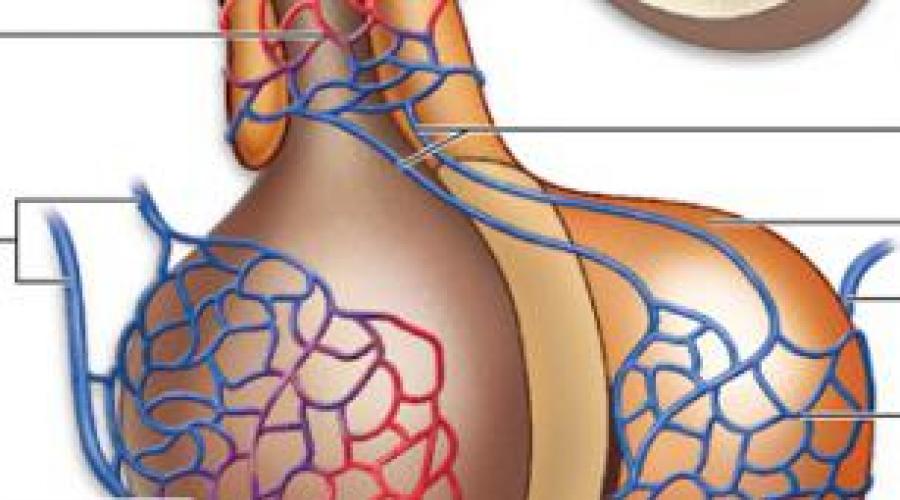
1. Крупноклеточные ядра образованы клеточными телами, которые в 2-3 раза крупнее, чем в других отделах гипоталамуса; к ним относятся супраоптическое (СОЯ) и паравентрикулярное (ПВЯ) ядра. СОЯ в 3-4 раза объемнее ПВЯ; ПВЯ лишь в центральных участках построено по типу крупноклеточного, а в периферических отделах образовано мелкими нейросекреторными клетками. Аксоны клеток СОЯ и ПВЯ покидают гипоталамус и, в составе гипоталамо-гипофизарного тракта пересекая гемато-энцефалическип барьер , проникают в заднюю долю гипофиза, где образуют терминали на капиллярах (рис. 2). Крупноклеточные ядра секретируют антидиуретический гормон (АДГ) или вазопрессин (ВП) , и окситоцин . Эти гормоны вырабатываются разными клетками. У человека АДГ образуется, главным образом, в СОЯ, а окситоцин - в ПВЯ. АДГ и окситоцин синтезируются в грЭПС (в виде крупной молекулы прогормона ) и переносятся в комплекс Гольджи, где упаковываются в гранулы. Процессинг продукта (завершается лишь в ходе транспорта гранул в аксоне) приводит к освобождению активного гормона и нейрофизина - белка с неясной функцией (ранее считали переносчиком нейрогормонов).

Рис. 2. Схема строения гипоталамо-гипофизарной нейросекреторной системы. ПДГ - передняя доля гипофиза, ПРДГ - промежуточная доля гипофиза, ЗДГ - задняя доля гипофиза, ККН - крупноклеточные нейроны, МКН - мелкоклеточные нейроны ПЗН - перекрест зрительных нервов, ГЭБ - гемато-энцефалический барьер, ПКС - первичная капиллярная сеть, ВКС - вторичная капиллярная сеть.
2. Мелкоклеточные ядра вырабатывают ряд гипофизотропных факторов, которые усиливают (рилизинг факторы или либерины ) или угнетают (ингабирующие факторы или статины ) выработку гормонов клетками передней доли, попадая к ним по воротной системе сосудов . Аксоны нейросекреторных клеток этих ядер образуют терминалы на первичной капиллярной сети в срединном возвышении (нейрогемальной контактной зоне). Эта сеть далее собирается в воротные вены , проникающие в переднюю долю гипофиза и распадающиеся на вторичную сеть капилляров между тяжами железистых клеток - аденоцитов (см. рис. 2).
Физиология гипоталамуса
Основные связи между нервной и эндокринной системами регуляции осуществляются посредством взаимодействия гипоталамуса и гипофиза (рис. 3).
Нервные импульсы, приходящие в гипоталамус, активируют секрецию так называемых рилизинг-факторов (либеринов и статинов): тиреолиберина, соматолиберина, пролактолиберина, гонадолиберина и кортиколиберина, а также соматостатина и пролактостатина. Мишенью для либеринов и статинов, секретируемых гипоталамусом, является гипофиз. Каждый из либеринов взаимодействует с определенной популяцией клеток гипофиза и вызывает в них синтез соответствующих тропинов : тиреотропина, соматотропного гормона (соматотропин - гормон роста), пролактина, гонадотропного гормона, (гонадотропины - лютеинизирующий и фолликулостимулирующий), а также адренокортикотропного гормона (АКТГ, кортикотропин). Статины оказывают на гипофиз влияние, противоположное действию либеринов, - подавляют секрецию тропинов. Тропины, секретируемые гипофизом, поступают в общий кровоток и, попадая на соответствующие железы, активируют в них секреторные процессы.

Рис 3 . Регуляция активности эндокринных желез центральной нервной системой при участии гипоталамуса и гипофиза. ТЛ - тиреолиберин; СЛ - соматалиберин; СС - соматостатин; ПЛ - пролактолиберин; ГЛ - гонадолиберин; КЛ - кортиколиберин; ТТГ - тиреотропный гормон; СТГ - соматотропный гормон (гормон роста); Пр - пролактин; ФСГ - фолликулостимулирующий гормон; ЛГ - лютеиниэирующий гормон; АКТГ - адренокортикотропный гормон. Сплошными стрелками обозначено активирующее, пунктирными - ингибирующее влияние.
Регуляция деятельности гипофиза и гипоталамуса, кроме сигналов, идущих «сверху вниз», осуществляется гормонами «исполнительных» желез (рис. 3). Эти «обратные» сигналы поступают в гипоталамус и затем передаются в гипофиз, что приводит к изменению секреции соответствующих тропинов. После удаления или атрофии эндокринной железы стимулируется секреция соответствующего тропного гормона; при гиперфункции железы секреция соответствующего тропина подавляется.

Рис.4 1 - медленно развивающееся и продолжительное ингибирование секреции гормонов и нейромедиаторов, а также изменение поведения и формирование памяти; 2 - быстро развивающееся, но продолжительное ингибирование; 3 - кратковременное ингибирование.
Обратные связи не только позволяют регулировать концентрацию гормонов в крови, но и участвуют в дифференцировке гипоталамуса в онтогенезе. Образование половых гормонов в женском организме происходит циклически, что объясняется циклической секрецией гонадотропных гормонов. Синтез этих гормонов контролируется гипоталамусом, образующим рилизинг-фактор этих тропинов (гонадолиберин). Если самке пересадить гипофиз самца, то пересаженный гипофиз начинает функционировать циклично. Половая дифференцировка гипоталамуса происходит под действием андрогенов. Если самца лишить половых желез, продуцирующих андрогены, то гипоталамус будет дифференцироваться по женскому типу.
В железах внутренней секреции иннервированы, как правило, только сосуды, а эндокринные клетки изменяют свою биосинтетическую и секреторную активность лишь под действием метаболитов, кофакторов и гормонов, причем не только гипофизарных. Так, ангиотензин II стимулирует синтез и секрецию альдостерона. Отметим также, что некоторые гормоны гипоталамуса и гипофиза могут образовываться не только в этих тканях. Например, соматостатин (гормон гипоталамуса, ингибирующий образование и секрецию гормона роста) обнаружен также в поджелудочной железе, где он подавляет секрецию инсулина и глюкагона.
Большинство нервных и гуморальных путей регуляции сходится на уровне гипоталамуса и благодаря этому в организме образуется единая нейроэндокринная регуляторная система. К клеткам гипоталамуса подходят аксоны нейронов, расположенных в коре больших полушарий и подкорковых образованиях. Эти аксоны секретируют различные нейромедиаторы, оказывающие на секреторную активность гипоталамуса как активирующее, так и тормозное влияние. Поступающие из мозга нервные импульсы гипоталамус «превращает» в эндокринные стимулы, которые могут быть усилены или ослаблены в зависимости от гуморальных сигналов, поступающих в гипоталамус от желез и тканей, подчиненных ему.
Тропины, образующиеся в гипофизе, не только регулируют деятельность подчиненных желез, но и выполняют самостоятельные эндокринные функции. Например, пролактин оказывает лактогенное действие, а также тормозит процессы дифференцировки клеток, повышает чувствительность половых желез к гонадотропинам, стимулирует родительский инстинкт. Кортикотропин является не только стимулятором стероцдогенеза, но и активатором липолиза в жировой ткани, а также важнейшим участником процесса превращения в мозге кратковременной памяти в долговременную. Гормон роста может стимулировать активность иммунной системы, обмен липидов, сахаров и т. д.
В задней доле гипофиза (нейрогипофиз) депонируются антидиуретический гормон (вазопрессин) и окситоцин (см. рис. 2). Первый вызывает задержку воды в организме и повышает тонус сосудов, второй стимулирует сокращение матки при родах и секрецию молока. Оба гормона синтезируются в гипоталамусе, затем транспортируются по аксонам в заднюю долю гипофиза, где депонируются и потом секретируются в кровь.
Характер процессов, протекающих в ЦНС, во многом определяется состоянием эндокринной регуляции. Так, андрогены и эстрогены формируют половой инстинкт, многие поведенческие реакции. Очевидно, что нейроны, точно так же как и другие клетки нашего организма, находятся под контролем гуморальной системы регуляции. Нервная система, эволюционно более поздняя, имеет как управляющие, так и подчиненные связи с эндокринной системой. Эти две регуляторные системы дополняют друг друга, образуют функционально единый механизм, что обеспечивает высокую эффективность нейрогуморальной регуляции, ставит ее во главе систем, согласующих все процессы жизнедеятельности в многоклеточном организме.
Гормоны гипоталамуса
К настоящему времени в гипоталамусе открыто 7 стимуляторов и 3 ингибитора секреции гормонов гипофиза, а именно: кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин,пролактолиберин, меланолиберин, соматостатин, пролактостатин и меланостатин . В чистом виде выделено 5 гормонов, для которых установлена первичная структура, подтвержденная химическим синтезом.
Большие трудности при получении гормонов гипоталамуса в чистом виде объясняются чрезвычайно низким содержанием их в исходной ткани.
По химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми олигопептидами необычного строения, хотя точный аминокислотный состав и первичная структура выяснены не для всех. Приводим полученные к настоящему времени данные о химической природе шести из известных 10 гормонов гипоталамуса.
(Пиро-Глу–Гис–Про–NH 2):
Тиролиберин представлен трипептидом , состоящим из пироглутаминовой (циклической) кислоты, гистидина и пролинамида, соединенных пептидными связями. В отличие от классических пептидов он не содержит свободных NH 2 - и СООН-групп у N- и С-концевых аминокислот.
Обеспечивает высвобождение тиротропного гормона (ТТГ) из передней доли гипофиза.
2. Гонадолиберин
является декапептидом, состоящим из 10 аминокислот последовательности:
Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2
Концевая С-аминокислота представлена глицинамидом.
Обеспечивает высвобождение ФСГ, ЛГ и пролактина
Является циклическим тетрадекапептидом (состоит из 14 аминокислотных остатков) :
Отличается этот гормон от двух предыдущих, помимо циклической структуры, тем, что не содержит на N-конце пироглутаминовой кислоты: дисульфидная связь образуется между двумя остатками цистеина в 3-м и 14-м положениях. Следует отметить, что синтетический линейный аналог соматостатина также наделен аналогичной биологической активностью, что свидетельствует о несущественности дисульфидного мостика природного гормона. Помимо гипоталамуса, соматостатин продуцируется нейронами центральной и периферической нервных систем, а также синтезируется в S-клетках панкреатических островков (островков Лангерганса) в поджелудочной железе и клетках кишечника. Он оказывает широкий спектр биологического действия; в частности, показано ингибирующее действие на синтез гормона роста в аденогипофизе, а также прямое тормозящее действие его на биосинтез инсулина и глюкагона в β- и α-клетках островков Лангерганса.
4. Соматолиберин. Он представлен 44 аминокислотными остатками с полностью раскрытой последовательностью. Биологической активностью соматолиберина наделен, кроме того, химически синтезированный декапептид:
Н-Вал–Гис–Лей–Сер–Ала–Глу–Глн–Лиз–Глу–Ала-ОН.
Этот декапептид стимулирует синтез и секрецию гормона роста гипофиза соматотропина.
5. Меланолиберин , химическая структура которого аналогична структуре открытого кольца гормона окситоцина (без трипептидной боковой цепи), имеет следующее строение:
Н-Цис–Тир–Иле–Глн–Асн–Цис-ОН.
6. Меланостатин (меланотропинингибирующий фактор) представленили трипептидом: Пиро-Глу–Лей–Гли-NН 2 , или пентапептидом со следующей последовательностью:
Пиро-Глу–Гис–Фен–Aрг–Гли–NН2.
Необходимо отметить, что меланолиберин оказывает стимулирующее действие, а меланостатин, напротив, ингибирующее действие на синтез и секрецию меланотропина в передней доле гипофиза.
Он выделяет гормоны, управляющие эндокринной системой. Секреторная активность проявляется через нейроны гипоталамуса. Вообще, можно сказать, что все нервные клетки выделяют гормоны. Они способны вырабатывать ацетилхолин, норадреналин и дофамин, которые работают в организме как медиаторы, то есть принимают участие в передаче различных нервных импульсов.
В гипоталамусе выделяются супраоптическое и паравентрикулярное ядра. В них секретируются, ответственно, вазопрессин и окситоцин. Данные гормоны совместно с белком-носителем через ножку гипофиза поступают к задней доле гипофиза, а она имеет с гипоталамусом общее неврогенное происхождение, но является при этом депо, где данные гормоны только накапливаются, но они там не продуцируются.
Какие гормоны выделяет гипоталамус?
Другими отделами гипоталамуса вырабатываются гипофизотропные гормоны (их часто также называют рилизинг-факторами). Они контролируют выделение гормонов передней доли гипофиза. Данная часть гипофиза не принадлежит эмбриологически мозгу, и при этом не имеет непосредственной иннервации со стороны гипоталамуса.
Она связана с гипоталамусом сетью сосудов, которая проходит по ножке гипофиза. Рилизинг-гормоны поступают по кровеносным сосудам в переднюю долю гипофиза, при этом регулируя синтез и выделение различных гипофизарных гормонов. Регуляция таких гормонов осуществляется стимулирующими, и одновременно различными ингибирующими гормонами гипоталамуса.
Но в отношении одних групп гипофизарных гормонов большее значение имеет регуляция их со стороны стимулирующих рилизинг-гормонов, а другой - воздействие ингибирующих гормонов гипоталамуса. При этом к первой группе гормонов относят АКТГ, ТТГ (тиреотропин), СТГ (гормон роста), ФСГ и ЛГ. Каждый из них регулируется соответствующими гипоталамическими рилизинг-гормонами.
На данный момент времени расшифрованы структуры ТТГ-РГ (то есть тиреотропин-рилизинг-гормона), который оказался трипептидом, а также СТГ-РГ, АКТГ-РГ и ЛГ-РГ, которые имеют структуру декапептидов.
С помощью синтетического ТТГ-РГ при условии внутривенного введения у здорового человека можно значительное повысить концентрацию в крови тиреотропина. МСГ и пролактин регулируются с помощью преимущественно ингибирующих гипоталамических факторов, соответственно МИФ и ПИФ. Потому в случае перерезки ножки гипофиза, тогда, когда устраняется воздействие гипоталамуса, секреция пролактина и МСГ увеличивается, а секреция других гипофизарных гормонов в то же время резко снижается.
Что еще может гипоталамус?
Помимо нейросекреторной активности, некоторые скопления гипоталамических нейронов играют также роль нейрогенных центров, которые регулируют некоторые основные функции организма. В частности, именно в гипоталамусе находится центр жажды. При этом нейрофизиологические данные показывают, что чувство жажды проявляется как гипоталамические сигналы в ответ на повышение уровня осмотического кровяного давления (сгущение крови), которое воспринимается осморецепторами гипоталамического супраоптического ядра.
В результате этого влияния, которое меняет электрические свойства мембран у осморецепторов, повышается секреция гормона вазопрессина, и в результате достигается задержка в организме воды.
Вместе с тем появляется ощущение жажды, которое направлено в итоге на восстановление осмотического давления. Рецепторы, которые расположены в разных отделах сосудистого ложа, одновременно с этим также воспринимают изменения объёма в организме циркулирующей крови.Информация поступает и в гипоталамус, и одновременно в ренин-ангиотензиновую систему. Это наряду с влиянием на гипоталамус ангиотензина оказывает регулирующее воздействие через почки.
Помимо центра жажды, в гипоталамусе находятся терморецепторы, которые воспринимают изменения в температуре крови. При этом имеются отдельные нейроны, которые реагируют на понижение и на повышение температуры (происходит гипоталамическая терморегуляция).
Важно упомянуть то, что серотонин и катехоламины, оказывая влияние на гипоталамический центр терморегуляции, могут менять температуру тела.
Гипоталамическая регуляция аппетита у людей связана в первую очередь с латеральным и вентромедиальным отделами гипоталамуса. Они работают соответственно как «центр аппетита» (голода) и «центр насыщения».
Раньше считалось, что в организме действует энергетически-температурный, липостатический и осмотический механизмы регуляции активности данных центров, а теперь считается, что регуляцию процессов аппетита и насыщения регулирует глюкостатический механизм.
При этом главную роль играет в первую очередь не только абсолютный уровень глюкозы в том или ином отделе гипоталамуса, где находятся глюкорецепторы, но интенсивность утилизации глюкозы в данных рецепторах.
Нужно подчеркнуть, что при гипогликемии, к примеру, в случае избытка в организме инсулина стимуляция аппетита также осуществляется за счёт того, что активизируются вторичные поведенческие реакции.
Ещё более важным является то, что нетолько то, в каком состоянии находится центр аппетита, но также регуляция секреции СТГ, которая имеет ключевое значение в обеспечении организма энергетическими субстратами, имеет связь с процессом утилизации глюкозы. Также возможно и то, что гипоталамус получает информацию о том, насколько интенсивно утилизируется глюкоза, на периферии, в первую очередь в печени.
Также с деятельностью гипоталамуса связана регуляция сна и бодрствования. Но здесь, также как и в отношении регуляции эмоциональных проявлений, гипоталамус больше проявляет себя уже как составная часть ретикулярной формации, контролирующей данные проявления.
Гипоталамус играет значимую роль также в процессах регулирования сердечно-сосудистой системы. Роль гипоталамических нарушений, например, повышения активности сосудорегулирующих центров в дальнейшем развитии гипертонической болезни является несомненной. То же самое можно сказать о регуляции вегетативных функций организма.
Она хоть и осуществляется разными отделами цнс, но при этом доминирующее воздействие оказывает гипоталамус. Характерно, что признаки симпатической активации, которая возникает при раздражении гипоталамуса, далее распространяются уже и на сердечно-сосудистую систему, и на функциональное состояние всего организма.
Гипофизотропная часть гипоталамуса и действие на организм гипоталамических нейронов в гипоталамических центрах находятся под контролем нейромедиаторов,образующихся преимущественно в самом гипоталамусе. Нервные окончания нейронов гипоталамуса отличаются специализацией в секреции гормонов дофамина, норадреналина и серотонина.
Адренергические нейроны повышают секрецию различных рилизинг-гормонов и вследствие этого секрецию АКТГ, гонадотропных гормонов, пролактина и СТГ и подавляют секрецию ингибирующих гормонов гипоталамуса.
Поэтому резерпин и аминазин, способные блокировать адренергическую передачу импульсов, влияют на снижение секреции гонадотропинов. АКТГ и СТГ, напротив, увеличивают секрецию гонадотропинов в результате подавления секреции ПИФ. Приэтом ДОФА, являясь предшественником норадреналина и дофамина, повышает концентрацию в головном мозге катехоламинов и потому тормозит секрецию гормона пролактина, но при этом повышает продукцию гонадотропинов, СТГ, ТТГ.
Но нужно отметить, что данные показали то, что норадреналинпродуцирующие и дофаминпродуцирующие нейроны, невзирая на свою адренергическую природу, в гипоталамусе часто обладают отдельными, специфическими функциями. Так,норадреналинпродуцирующие нейроны осуществляют контроль также секреции вазопрессина и окситоцина. Серотонинпродуцирующие нейроны точно так же связаны с механизмами, контролирующими секрецию АКТГ и гонадотропинов, при этом в мозге концентрация серотонина снижает продукцию гонадотропинов, например, ЛГ.
Именно этим объясняется то, что имипрамин, который блокирует транспорт серотонина, влияет на изменение эстрального цикла, а -этил-триптамин, активирующий серотониновые рецепторы, снижает секрецию гормона АКТГ. Мелатонин и некоторые другие метоксииндолы влияют на гипоталамус, воздействуя на уровне серотонинпродуцирующих нейронов, при этом вызывая снижение секреции МСГ, гонадотропинов, снижение функции щитовидки и стимулируют «центр сна».