Клеточный митоз. Митотическое деление
Читайте также
Учебник для 10-11 классов
Раздел II. Размножение и развитие организмов
Глава V. Размножение организмов
Каждую секунду на Земле гибнет от старости, болезней и хищников астрономическое количество живых существ, и только благодаря размножению, этому универсальному свойству организмов, жизнь на Земле не прекращается.
Может показаться, что процессы размножения у живых существ очень разнообразны, однако все их можно свести к двум формам: бесполому и половому. У некоторых организмов встречаются разные формы размножения. Например, многие растения могут размножаться черенками, отводками, клубнями (бесполое размножение) и семенами (половое).
При половом размножении каждый организм развивается из одной клетки, образующейся от слияния двух половых клеток - мужской и женской.
В основе размножения и индивидуального развития организма лежит процесс деления клеток.
§ 20. Деление клетки. Митоз
Способность к делению - важнейшее свойство клеток. Без деления невозможно представить себе увеличение числа одноклеточных существ, развитие сложного многоклеточного организма из одной оплодотворенной яйцеклетки, возобновление клеток, тканей и даже органов, утраченных в процессе жизнедеятельности организма.
Деление клеток осуществляется поэтапно. На каждом этапе деления происходят определенные процессы. Они приводят к удвоению генетического материала (синтезу ДНК) и его распределению между дочерними клетками. Период жизни клетки от одного деления до следующего называют клеточным циклом.
Подготовка к делению. Эукариотические организмы, состоящие из клеток, имеющих ядра, начинают подготовку к делению на определенном этапе клеточного цикла, в интерфазе.
Именно в период интерфазы в клетке происходит процесс биосинтеза белка, удваиваются хромосомы. Вдоль исходной хромосомы из имеющихся в клетке химических соединений синтезируется ее точная копия, удваивается молекула ДНК. Удвоенная хромосома состоит из двух половинок - хроматид. Каждая из хроматид содержит одну молекулу ДНК.
Интерфаза в клетках растений и животных в среднем продолжается 10-20 ч. Затем наступает процесс деления клетки - митоз.
Во время митоза клетка проходит ряд последовательных фаз, в результате которых каждая дочерняя клетка получает такой же набор хромосом, какой был в материнской клетке.
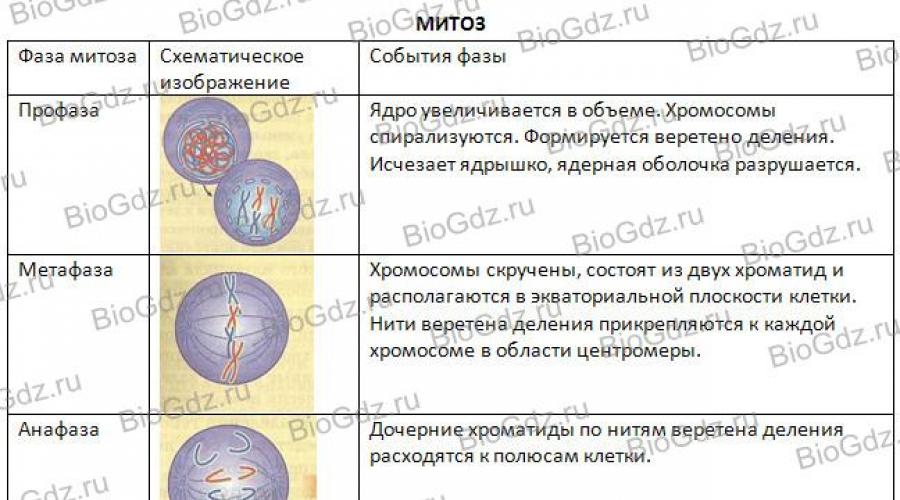
Фазы митоза. Различают следующие четыре фазы митоза: профаза, метафаза, анафаза и телофаза. На рисунке 29 схематически показан ход митоза. В профазе хорошо видны центриоли - образования, находящиеся в клеточном центре и играющие роль в расхождении дочерних хромосом животных. (Напомним, что только у некоторых растений есть центриоли в клеточном центре, который организует расхождение хромосом.) Мы же рассмотрим митоз на примере животной клетки, поскольку присутствие центриоли делает процесс расхождения хромосом более наглядным. Центриоли удваиваются и расходятся к разным полюсам клетки. От центриолей протягиваются микротрубочки, образующие нити веретена деления, которое регулирует расхождение хромосом к полюсам делящейся клетки.
Рис. 29. Схема митоза
В конце профазы ядерная оболочка распадается, ядрышко постепенно исчезает, хромосомы спирализуются и в результате этого укорачиваются и утолщаются, и их уже можно наблюдать в световой микроскоп. Еще лучше они видны на следующей стадии митоза - метафазе.
В метафазе хромосомы располагаются в экваториальной плоскости клетки. При этом хорошо видно, что каждая хромосома, состоящая из двух хроматид, имеет перетяжку - центромеру. Хромосомы своими центромерами прикрепляются к нитям веретена деления. После деления центромеры каждая хроматида становится самостоятельной дочерней хромосомой.
Затем наступает следующая стадия митоза - анафаза, во время которой дочерние хромосомы (хроматиды одной хромосомы) расходятся к разным полюсам клетки.
Следующая стадия деления клетки - телофаза. Она начинается после того, как дочерние хромосомы, состоящие из одной хроматиды, достигли полюсов клетки. На этой стадии хромосомы вновь деспирализуются и приобретают такой же вид, какой они имели до начала деления клетки в интерфазе (длинные тонкие нити). Вокруг них возникает ядерная оболочка, а в ядре формируется ядрышко, в котором синтезируются рибосомы. В процессе деления цитоплазмы все органоиды (митохондрии, комплекс Гольджи, рибосомы и др.) распределяются между дочерними клетками более или менее равномерно.
Таким образом, в результате митоза из одной клетки получаются две, каждая из которых имеет характерное для данного вида организма число и форму хромосом, а следовательно, постоянное количество ДНК.
Весь процесс митоза занимает в среднем 1-2 ч. Продолжительность его несколько различна для разных видов клеток. Зависит она также и от условий внешней среды (температуры, светового режима и других показателей).
Биологическое значение митоза заключается в том, что он обеспечивает постоянство числа хромосом во всех клетках организма. В процессе митоза происходит распределение ДНК хромосом материнской клетки строго поровну между возникающими из нее двумя дочерними клетками. В результате митоза все дочерние клетки получают одну и ту же генетическую информацию.
- Какие изменения в клетке предшествуют делению?
- Когда образуется веретено деления? Какова его роль?
- Охарактеризуйте фазы митоза и кратко расскажите, как происходит этот процесс.
- Что такое хроматида? Когда она становится хромосомой?
- Что такое центромера? Какую роль она выполняет при митозе?
- В чем заключается биологическое значение митоза?
Вспомните из курса ботаники, зоологии, анатомии, физиологии и гигиены человека, как происходит размножение в органическом мире.
1. Дайте определения понятий.
Интерфаза
– фаза подготовки к митотическому делению, когда происходит удвоение ДНК.
Митоз
– это деление, в результате которого происходит строго одинаковое распределение точно скопированных хромосом между дочерними клетками, что обеспечивает образование генетически идентичных клеток.
Жизненный цикл
– период жизни клетки от момента ее возникновения в процессе деления до гибели или конца последующего деления.
2. Чем рост одноклеточных организмов отличается от роста многоклеточных?
Рост одноклеточного организма – это увеличение размеров и усложнение строения отдельной клетки, а рост многоклеточных – это также и активное деление клеток – увеличение их количества.
3. Почему в жизненном цикле клетки обязательно существует интерфаза?
В интерфазе происходит подготовка к делению и удвоение ДНК. Если бы его не происходило, то при каждом делении клетки количество хромосом уменьшалось бы вдвое, и довольно скоро в клетке вообще бы не осталось хромосом.
4. Заполните кластер «Фазы митоза».
5. Используя рисунок 52 в § 3.4, заполните таблицу.

6. Составьте синквейн к термину «митоз».
Митоз
Четырехфазный, равномерный
Делит, распределяет, дробит
Поставляет генетический материал дочерним клеткам
Клеточное деление.
7. Установите соответствие между фазами митотического цикла и событиями, происходящими в них.
Фазы
1. Анафаза
2. Метафаза
3. Интерфаза
4. Телофаза
5. Профаза
События
A. Клетка растет, образуются органоиды, удваивается ДНК.
Б. Хроматиды расходятся и становятся самостоятельными хромосомами.
B. Начинается спирализация хромосом, разрушается ядерная оболочка.
Г. Хромосомы располагаются в экваториальной плоскости клетки. Нити веретена деления присоединяются к центромерам.
Д. Исчезает веретено деления, формируются ядерные оболочки, хромосомы раскручиваются.
8. Почему завершение митоза – деление цитоплазмы происходит по-разному в животных и растительных клетках?
В животных клетках нет клеточной стенки, у них клеточная мембрана впячивается внутрь, и клетка делится путем перетяжки.
В клетках растений мембрана формируется в экваториальной плоскости внутри клетки и, распространяясь к периферии, делит клетку пополам.
9. Почему в митотическом цикле интерфаза занимает гораздо более продолжительное время, чем само деление?
Во время интерфазы клетка усиленно готовится к митозу, в ней идут процессы синтеза, удвоение ДНК, клетка растет, проходит ее жизненный цикл, не включая само деление.
10. Выберите правильный ответ.
Тест 1.
В результате митоза из одной диплоидной клетки образуются:
4) 2 диплоидные клетки.
Тест 2.
Деление центромер и расхождение хроматид к полюсам клетки происходит в:
3) анафазе;
Тест 3.
Жизненный цикл - это:
2) жизнь клетки от деления до конца следующего деления или смерти;
Тест 4.
Какой термин написан с орфографической ошибкой?
4) телафаза.
11. Объясните происхождение и общее значение слова (термина), опираясь на значение корней, его составляющих.

12. Выберите термин и объясните, насколько его современное значение соответствует первоначальному значению его корней.
Выбранный термин – интерфаза.
Соответствие. Термин соответствует, и означает период между фазами митоза, когда происходит подготовка к делению.
13. Сформулируйте и запишите основные идеи § 3.4.
Жизненный цикл – это жизнь клетки от деления до конца следующего деления или смерти. Между делениями клетка подготавливается к нему в период интерфазы. В это время происходит синтез веществ, удвоение ДНК.
Клетка делится митозом. Он состоит из 4 стадий:
Профаза.
Метафаза.
Анафаза.
Телофаза.
Цель митоза: в результате его из 1 материнской клетки образуются 2 дочерние с идентичным набором генов. Количество генетического материала и хромосом при этом остается одинаковым, обеспечивается генетическая стабильность клеток.
Важнейшим компонентом клеточного цикла является митотический (пролиферативный) цикл. Он представляет собой комплекс взаимосвязанных и согласованных явлений во время деления клетки, а также до и после него. Митотический цикл - это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием двух клеток следующей генерации. Кроме этого, в понятие жизненного цикла входят также период выполнения клеткой своих функций и периоды покоя. В это время дальнейшая клеточная судьба неопределенна: клетка может начать делиться (вступает в митоз) либо начать готовиться к выполнению специфических функций.
Основные стадии митоза
1.
Редупликация (самоудвоение) генетической информации материнской клетки и
равномерное распределение ее между дочерними клетками. Это сопровождается
изменениями структуры и морфологии хромосом, в которых сосредоточено более 90%
информации эукариотической клетки.
2.
Митотический цикл состоит из четырех последовательных периодов: пресинтетического (или постмитотического) G1, синтетического S,
постсинтетического (или премитотического) G2 и собственно митоза. Они составляют
автокаталитическую интерфазу (подготовительный период).
Фазы клеточного цикла:
1) пресинтетическая (G1). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки (гистоны, структурные белки, ферменты), РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов (т. е. структур, способных к ауторепродукции). Восстанавливаются черты организации интерфазной клетки после предшествующего деления;
2)
синтетическая (S). Происходит удвоение генетического материала путем
репликации ДНК. Она происходит полуконсервативным способом, когда двойная
спираль молекулы ДНК расходится на две цепи и на каждой из них синтезируется комплементарная цепочка.
В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит
из одной новой и старой цепи ДНК. Количество наследственного материала
удваивается. Кроме этого, продолжается синтез РНК и белков. Также репликации
подвергается небольшая часть митохонд-риальной ДНК (основная же ее часть
реплицируется в G2 период);
3) постсинтетическая (G2). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных).
S и G2 непосредственно связаны с митозом, поэтому их иногда выделяют в отдельный
период - препрофазу.
После этого наступает собственно митоз, который состоит из четырех фаз. Процесс
деления включает в себя несколько последовательных фаз и представляет собой
цикл. Его продолжительность различна и составляет у большинства клеток от 10 до
50 ч. При этом у клеток тела человека продолжительность самого митоза составляет
1-1,5 ч, G2-периода интерфазы - 2-3 ч, S-периода интерфазы - 6-10 ч.
Длительность отдельных стадий различна и варьируется в зависимости от типа
ткани, физиологического состояния организма, внешних факторов. Наиболее
продолжительны стадии сопряженные с процессами внутриклеточного синтеза: профаза
и телофаза. Наиболее быстротечны фазы митоза, в ходе которых происходит движение
хромосом: метафаза и анафаза. Непосредственно процесс расхождения хромосом к
полюсам обычно не превышает 10 минут.
Профаза
К основным событиям профазы относят конденсацию хромосом внутри ядра и
образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе
является характерной, но не обязательной для всех клеток особенностью.
Условно за начало профазы принимается момент возникновения микроскопически
видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение
хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения
сопровождаются повышением активности фосфорилаз, модифицирующих гистоны,
непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается
транскрипционная активность хроматина, инактивируются ядрышковые гены, большая
часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в
ранней профазе остаются спаренными по всей своей длине с помощью
белков-когезинов, однако к началу прометафазы связь между хроматидами
сохраняется лишь в области центромер. К поздней профазе на каждой центромере
сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для
присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено - одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом.
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.
Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
В клетках млекопитающих прометафаза протекает, как правило, в течение 10-20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона - около 30 минут.
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи - к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.
В метафазе, также как и в течение других фаз митоза, продолжается активное
обновление микротрубочек веретена путём интенсивной сборки и деполимеризации
молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных
микротрубочек, происходит постоянная переборка межполюсных микротрубочек,
численность которых в метафазе достигает максимума.
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид,
соединение между которыми сохраняется лишь в центромерных участках. Плечи
хроматид располагаются параллельно друг другу, и становится отчетливо заметной
разделяющая их щель.
Анафаза
Анафаза - самая короткая стадия митоза, которая начинается внезапным
разделением и последующим расхождением сестринских хроматид в направлении
противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью
достигающей 0,5-2 мкм/мин., при этом они часто принимают V-образную форму. Их
движение обусловлено воздействием значительных сил, оценочно 10 дин на
хромосому, что в 10 000 раз превышает усилие, необходимое для простого
продвижения хромосомы через цитоплазму с наблюдаемой скоростью.
Как правило, расхождение хромосом в анафазе состоит из двух относительно
независимых процессов называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным
полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее
удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения
хроматид сопровождается сокращением длины деполимеризующихся кинетохорных
микротрубочек. Причем их распад наблюдается преимущественно в области
кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у
кинетохоров либо в области полюсов деления является необходимым условием для
перемещения сестринских хроматид, так как их движение прекращается при
добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на
микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока
остается неизвестным.
Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от
анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со
стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при
взаимодействии отчасти и создают расталкивающее полюса усилие. Величина
относительного перемещения полюсов при этом, также как и степень перекрывания
полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей
разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие
силы со стороны астральных микротрубочек, которые создаются в результате
взаимодействия с динеино-подобными белками на плазматической мембране клетки.
Последовательность, продолжительность и относительный вклад каждого из двух
процессов, слагающих анафазу, могут быть крайне различны. Так в клетках
млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к
противоположным полюсам и продолжается вплоть до удлинения митотического
веретена в 1,5-2 раза по сравнению с метафазным. В некоторых других клетках
анафаза В начинается только после того как хроматиды достигают полюсов деления.
У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по
сравнению с метафазным. В растительных клетках анафаза В отсутствует.
Телофаза
Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.
Окончание телофазы преимущественно совпадает с разделением тела материнской
клетки - цитокинезом. При этом образуются две или более дочерние клетки.
Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине
анафазы и могут продолжаться после завершения телофазы. Митоз не всегда
сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в
качестве отдельной фазы митотического деления и обычно рассматривается в составе
телофазы.
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и
деление путём образования клеточной пластинки. Плоскость деления клетки
детерминируется положением митотического веретена и проходит под прямым углом к
длинной оси веретена.
При делении поперечной перетяжкой клетки место разделения цитоплазмы
закладывается предварительно ещё в период анафазы, когда в плоскости метафазной
пластинки под мембраной клетки возникает сократимое кольцо из актиновых и
миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца,
образуется борозда деления, которая постепенно углубляется вплоть до полного
разделения клетки. По окончании цитокинеза сократимое кольцо полностью
распадается, а плазматическая мембрана стягивается вокруг остаточного тельца
Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек,
тесно упакованных вместе с материалом плотного матрикса.
Деление путём образования клеточной пластинки начинается с перемещения мелких
ограниченных мембраной пузырьков по направлению к экваториальной плоскости
клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру
- раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата
Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных
микротрубочек веретена деления, образующих цилиндрическую структуру, называемую
фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего
фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых
мембранных пузырьков продолжается рост клеточной пластинки вплоть до её
окончательного слияния с мембраной материнской клетки. После окончательного
разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы
целлюлозы, завершая образование жёсткой клеточной стенки.
Для определения завершения каждой фазы клеточного цикла необходимо наличие в нем контрольных точек. Если клетка «проходит» контрольную точку то она продолжается «двигаться» по клеточному циклу. Если же какие-либо обстоятельства, например повреждение ДНК, мешают клетке пройти через контрольную точку, которую можно сравнить со своего рода контрольным пунктом, то клетка останавливается и другой фазы клеточного цикла не наступает по крайней мере до тех пор, пока не будут устранены препятствия, не позволявшие клетке пройти через контрольный пункт.
Митотическое деление клеток
Митоз (от греч. Mitos - нить), называемый также кариокинезом, или непрямым делением клеток, является универсальным механизмом деления клеток. Митоз следует за G2-периодом и завершает клеточный цикл.
Он длится 1-3 часа и обеспечивает равномерное распределение генетического материала в дочерние клетки. Митоз включает 4 основные фазы: профазу, метафазу, анафазу и телофазу.
Митоз - это один из фундаментальных процессов онтогенеза. Митотическое деление обеспечивает рост многоклеточных эукариот за счёт увеличения популяций клеток тканей.
В результате митотического деления клеток меристем увеличивается количество клеток тканей растений. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений.
На основании морфологических особенностей митоз условно подразделяется на стадии: профазу, прометафазу, метафазу, анафазу, телофазу. Первые описания фаз митоза и установление их последовательности были предприняты в 70-80-х годах XIX века. В конце 1870-х годов немецкий гистолог Вальтер Флемминг для обозначения процесса непрямого деления клетки ввёл термин «митоз».
Продолжительность митоза в среднем составляет 1-2 часа. Митоз клеток животных, как правило, длится 30-60 минут, а растений - 2-3 часа. За 70 лет в теле человека суммарно осуществляется порядка 10 14 клеточных делений .
Первые неполные описания, касающиеся поведения и изменения ядер в делящихся клетках, встречаются в работах учёных начала 1870-х годов.
В работе русского ботаника Руссова, датируемой 1872 годом, отчётливо описаны и изображены метафазные и анафазные пластинки, состоящие из отдельных хромосом.
Годом позже немецкий зоолог Г.А. Шнейдер ещё более отчётливо и последовательно, но, конечно, не совсем полно описал митотическое деление на примере дробящихся яиц прямокишечной турбеллярии Mesostomum. В его работе, в сущности, описаны и проиллюстрированы в правильной последовательности основные фазы митоза: профаза, метафаза, анафаза (ранняя и поздняя). В 1874 году московский ботаник И.Д. Чистяков также наблюдал отдельные фазы клеточного деления в спорах плаунов и хвощей. Несмотря на первые успехи ни Руссову, ни Шнейдеру, ни Чистякову не удалось дать чёткое и последовательное описание митотического деления.
В 1875 году вышли работы, содержащие более детальные описания митозов. О. Бючли дал описание цитологических картин в дробящихся яйцах круглых червей и моллюсков и в сперматогенных клетках насекомых.
Э. Страсбургер исследовал митотическое деление в клетках зелёной водоросли спирогиры, в материнских клетках пыльцы лука и в материнских споровых клетках плауна. Ссылаясь на работу О. Бючли и основываясь на собственных исследованиях, Э. Страсбургер обратил внимание на единство процессов клеточного деления в растительных и животных клетках.
К концу 1878 - началу 1879 года появились подробные работы Шлейхера и В. Флемминга. В своей работе в 1879 году Шлейхер предложил термин «кариокинез» для обозначения сложных процессов клеточного деления, подразумевая перемещения составных частей ядра. Вальтер Флемминг впервые для обозначения непрямого деления клетки ввёл термин «митоз», который впоследствии стал общепринятым. Также Флеммингу принадлежит окончательная формулировка определения митоза как циклического процесса, завершающегося разделением хромосом между дочерними клетками .
В 1880 г. О.В. Баранецкий установил спиральное строение хромосом. В ходе дальнейших исследований были развиты представления о спирализации и деспирализации хромосом во время митотического цикла.
В начале 1900-х годов хромосомы были идентифицированы в качестве носителей наследственной информации, что в дальнейшем дало объяснение биологической роли митоза, заключающейся в образовании генетически идентичных дочерних клеток .
В 1970-х годах началась расшифровка и детальное изучение регуляторов митотического деления, благодаря серии экспериментов по слиянию клеток, находящихся на разных этапах клеточного цикла. В тех опытах, когда клетку в М-фазе объединяли с клеткой, находящейся в любой из стадий интерфазы (G 1 , S или G 2), интерфазные клетки переходили в митотическое состояние (начиналась конденсация хромосом и распадалась ядерная оболочка).
В итоге был сделан вывод, что в цитоплазмемитотической клетки присутствует фактор (или факторы), стимулирующий митоз, или, иначе, М-стимулирующий фактор (МСФ, от англ. M-phase-promoting factor, MPF).
Впервые «фактор стимуляции митоза» был открыт в зрелых неоплодотворенных яйцах шпорцевой лягушки, находящихся в М-фазе клеточного цикла. Цитоплазма такого яйца, инъецированная в ооцит, приводила к преждевременному переходу в М-фазу и к началу созревания ооцита (первоначально сокращение MPF означало Maturation Promoting Factor, что переводится как «фактор, способствующий созреванию»). В ходе дальнейших экспериментов были установлены универсальное значение и вместе с тем высокая степень консервативности «фактора стимуляции митоза»: экстракты, приготовленные из митотических клеток весьма разнообразных организмов, при введении в ооциты шпорцевой лягушки переводили их в М-фазу.
В ходе последующих исследований выяснилось, что фактор, стимулирующий митоз, представляет собой гетеродимерный комплекс, состоящий из белка циклина и зависимой от циклина протеинкиназы. Циклин является регуляторным белком и обнаруживается у всех эукариот. Его концентрация периодически возрастает в течение клеточного цикла, достигая максимума в метафазе митоза. С началом анафазы наблюдается резкое сокращение концентрации циклина, вследствие его расщепления с помощью сложных белковых протеолитических комплексов - протеосом. Зависимая от циклина протеинкиназа представляет собой фермент (фосфорилазу), модифицирующий белки за счёт переноса фосфатной группы от АТФ на аминокислоты серин и треонин. Таким образом, с установления роли и структуры основного регулятора митотического деления начались исследования тонких регуляторных механизмов митоза, которые продолжаются до настоящего времени .
Выработка единой типологии и классификации митозов осложняется целым спектром признаков, которые в различных комбинациях создают разнообразие и неоднородность картин митотического деления. При этом отдельные варианты классификации, разработанные применительно к одним таксонам, являются неприемлемыми в отношении других, поскольку не учитывают специфики их митозов. Например, отдельные варианты классификации митозов, свойственных животным или растительным организмам, оказываются неприемлемыми для водорослей.
Одним из ключевых признаков, лежащих в основе различных типологий и классификаций митотического деления, является поведение ядерной оболочки. Если образование веретена и само митотическое деление протекает внутри ядра без разрушения ядерной оболочки, то такой тип митоза называют закрытым. Митоз с распадом ядерной оболочки, соответственно, называется открытым, а митоз с распадом оболочки только на полюсах веретена, с образованием «полярных окон» - полузакрытым.
Ещё одним характерным признаком является тип симметрии митотического веретена. При плевромитозе веретено деления билатерально симметрично либо асимметрично и состоит, как правило, из двух полуверетён, располагающихся в метафазе-анафазе под углом друг к другу. Для категории ортомитозов характерна биполярная симметрия веретена деления, а в метафазе зачастую наблюдается различимая экваториальная пластинка.
В рамках обозначенных признаков наиболее многочисленным является типичный открытый ортомитоз, на примере, которого ниже рассматриваются принципы и стадии митотического деления. Данный тип митоза характерен для животных, высших растений и некоторых простейших .
Профаза начинается с конденсации хромосом, которые становятся видимыми в световой микроскоп как нитевидные структуры. Каждая хромосома состоит из двух параллельно лежащих сестринских хроматид, связанных в области центромеры. Ядрышко и ядерная оболочка к концу фазы исчезают (последняя распадается на мембранные пузырьки, сходные с элементами ЭПС, а поровый комплекс и ламина диссоциируют на субъединицы). Кариоплазма смешивается с цитоплазмой.
Центриоли мигрируют к противоположным полюсам клетки и дают начало нитям митотического (ахроматинового) веретена. В области центромеры образуются особые белковые комплексы - кинетохоры, к которым прикрепляются некоторые микротрубочки веретена (кинетохорные микротрубочки); показано, что кинетохоры сами способны индуцировать сборку микротрубочек и поэтому могут служить центрами организации микротрубочек. Остальные микротрубочки веретена называются полюсными, так как они протягиваются от одного полюса клетки к другому; лежащие вне веретена микротрубочки, расходящиеся радиально от клеточных центров к плазмолемме, получили наименование астральных или микротрубочек (нитей) сияния.
Метафаза соответствует максимальному уровню конденсации хромосом, которые выстраиваются в области экватора митотического веретена, образуя картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со стороны полюсов). Хромосомы перемещаются в экваториальную плоскость и удерживаются в ней благодаря сбалансированному натяжению кинетохорных микротрубочек. Сестринские хроматиды к концу этой фазы разделяются щелью, однако удерживаются в области центромеры.
Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромеры) и движения дочерних хромосом к противоположным полюсам клетки, которое происходит вдоль микротрубочек веретена со скоростью 0,2-0,5 мкм/мин. Сигнал к началу анафазы включает резкое (на порядок) повышение концентрации катионов кальция в гиалоплазме, выделяемого мембранными пузырьками, образующими скопления у полюсов веретена. Механизм движения хромосом в анафазе окончательно не выяснен, однако установлено, что в области веретена помимо актина имеются такие белки как миозин и динеин, а также ряд регуляторных белков. По некоторым наблюдениям, оно обусловлено укорочением (разборкой) микротрубочек, прикрепленных к кинетохорам. Анафаза характеризуется удлинением митотического веретена за счет некоторого расхождения полюсов клетки. Она завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картины звезд (стадия дочерних звезд). В конце анафазы благодаря сокращению актиновых микрофиламентов, концентрирующихся по окружности клетки (сократимое кольцо), начинает образовываться клеточная перетяжка, которая углубляясь, в следующей фазе приведет к цитотомии.
Телофаза - это конечная стадия митоза, в течение которой реконструируются ядра дочерних клеток и завершается их разделение. Вокруг конденсированных хромосом дочерних клеток из мембранных пузырьков (по другим данным, из ЭПС) восстанавливается кариолемма, с которой связывается формирующаяся ламина, вновь появляются ядрышки, которые образуются из участков соответствующих хромосом. Ядра клеток постепенно увеличиваются, а хромосомы прогрессивно деспирализуются и исчезают, замещаясь картиной хроматина интерфазного ядpa. Одновременно происходит углубление клеточной перетяжки, и клетки в течение некоторого времени остаются связанными суживающимся цитоплазматическим мостиком, содержащим пучок микротрубочек (срединное тельце). Дальнейшая перешнуровка цитоплазмы завершается формированием двух дочерних клеток. В телофазе происходит распределение органелл между дочерними клетками; равномерности этого процесса способствует то, что одни органеллы достаточно многочисленны (например, митохондрии), другие (подобно ЭПС и комплексу Гольджи) во время митоза распадаются на мелкие фрагменты и пузырьки.
Атипические митозы возникают при повреждении митотического аппарата и характеризуются неравномерным распределением генетического материала между клетками - анэуплоидией (от греч. аn - не, eu - правильное, ploon - складываю); во многих случаях цитотомия отсутствует, в результате чего формируются гигантские клетки. Атипические митозы характерны для злокачественных опухолей и облученных тканей. Чем выше их частота и чем значительнее степень анэуплоидии, тем более злокачественной является опухоль. Нарушение нормального митотического деления клеток может обусловливаться аномалиями хромосом, которые называют хромосомными аберрациями (от лат. Aberratio - отклонение). Вариантами хромосомных аберраций служат слипание хромосом, их разрыв на фрагменты, выпадение участка, обмен фрагментами, удвоение отдельных участков хромосом и др. Хромосомные аберрации могут возникать спонтанно, но чаще развиваются вследствие действия на клетки мутагенов и ионизирующего облучения.
Кариотипирование - диагностическое исследование с целью оценки кариотипа (набора хромосом) производится путем изучения хромосом в метафазной пластинке. Для кариотипирования получают культуру клеток, в которую вводят колхицин - вещество, блокирующее формирование митотического веретена. Из таких клеток извлекают хромосомы, которые далее окрашивают и идентифицируют. Нормальный кариотип человека представлен 46 хромосомами - 22 парами аутосом и двумя половыми хромосомами (XY у мужчин и XX у женщин). Кариотипирование позволяет диагностировать ряд заболеваний, связанных с хромосомными аномалиями, в частности, синдромы Дауна (трисомия 21-й хромосомы), Эдвардса (трисомия 18-й хромосомы), Патау (трисомия 13-й хромосомы), а также ряд синдромов, связанных с аномалиями половых хромосом - синдром Кляйнфельтера (генотип - XXY), Турнера (генотип - ХО) и другие .
Предполагается, что сложный митотический процесс высших организмов развивался постепенно из механизмов деления прокариот. Это предположение подтверждается тем, что прокариоты появились около миллиарда лет раньше первых эукариот. Кроме того, в митозе эукариот и бинарном делении прокариот принимают участие схожие белки.
Возможные промежуточные стадии между бинарным делением и митозом можно проследить у одноклеточных эукариот, у которых в ходе деления не разрушается ядерная оболочка. У большинства же других эукариот, в том числе растений и животных, веретено деления формируется вне ядра, а ядерная оболочка разрушается в течение митоза. Хотя митоз у одноклеточных эукариот ещё недостаточно изучен, можно предположить, что он произошёл от бинарного деления и в конечном счёте достиг того уровня сложности, который имеется у многоклеточных организмов.
У многих простейших эукариот митоз также остался процессом, связанным с мембраной, однако теперь уже не плазматической, а ядерной.
Основными регуляторными механизмами митоза являются процессы фосфорилирования и протеолиза.
Обратимые реакции фосфорилирования и дефосфорилирования обеспечивают протекание обратимых событий митоза, таких как сборка/распад веретена деления или распад/восстановление ядерной оболочки. Протеолиз лежит в основе необратимых событий митоза, таких как разделение сестринских хроматид в анафазе или разрушение митотических циклинов на поздних стадиях митоза.
Деление всех эукариотических клеток сопряжено с формированием специального аппарата клеточного деления.
Активная роль в митотическом делении клеток зачастую отведена цитоскелетным структурам. Универсальным как для животных, так и для растительных клеток является двухполюсное митотическое веретено, состоящее из микротрубочек и связанных с ними белков. Веретено деления обеспечивает строго одинаковое распределение хромосом между полюсами деления, в области которых в телофазе образуются ядра дочерних клеток.
Процесс митоза обеспечивает строго равномерное распределение хромосом между двумя дочерними ядрами, так что в многоклеточном организме все клетки имеют совершенно одинаковые (по числу и по характеру) наборы хромосом.
Хромосомы содержат генетическую информацию, закодированную в ДНК, и поэтому регулярный, упорядоченный митотический процесс обеспечивает также полную передачу всей информации каждому из дочерних ядер; в результате каждая клетка обладает всей генетической информацией, необходимой для развития всех признаков организма. В связи с этим становится понятно, почему одна клетка, взятая из полностью дифференцированного взрослого растения, может при подходящих условиях развиться в целое растение. Мы описали митоз в диплоидной клетке, но этот процесс протекает сходным образом и в гаплоидных клетках, например в клетках гаметофитного поколения растений .
Щенок растет и увеличивается в размерах за счет деления соматических клеток, называемого митозом. Митоз - непрямое деление соматической клетки, когда происходят сложные изменения в ее ядре и цитоплазме. После оплодотворения (овогамии) яйцеклетки сперматозоидом (слияния, или копуляции гамет) образуется зигота (ооциста) - новый организм, состоящий всего из одной клетки. Процесс роста и развития нового организма начинается с момента первого митотического деления этой клетки (материнской), когда из нее возникают две дочерние (точнее, сестринские), полностью схожие с ней клетки, и продолжается до смерти
Рис.1 Строение клетки
В процессе митоза происходит:
1- удвоение вещества хромосом;
2- изменение физического состояния и химической организации хромосом;
3- расхождение сестринских хромосом к полюсам клетки;
4- последующее деление цитоплазмы и полное восстановление двух ядер в новых клетках.
В митозе заложен жизненный цикл ядерных генов: удвоение, распределение и функционирование. Период между делениями клетки называется интерфазой , во время которой в ней протекают активные процессы жизнедеятельности и подготовка к следующему делению. Весь цикл изменений, происходящих в клетке
от одного ее деления до другого, называется митотическим циклом. Последний, состоит из двух основных периодов - интерфазы и непосредственно митоза.
В результате митоза из одной клетки образуются две, имеющие идентичные хромосомы. Таким образом, митоз обеспечивает преемственность и постоянство числа и набора, то есть качественной специфичности хромосом в последовательных поколениях делящихся клеток (см. рис. 2).
В интерфазе, периоде между двумя последовательными делениями клетки, в ядре происходит репликация (аутодупликация, или самоудвоение) ДНК, а значит, и числа хромосом в клетке (образование сестринских хроматид, удерживаемых вместе центромерой, то есть тельцем, выполняющим функцию механического центра хромосомы), а также деспирализация последних.
В метафазе, или центральной фазе деления ядра, хромосома, состоящая из двух хроматид, превращается в две дочерние хромосомы.
Рис. 2. Митоз
 1 - три пары хромосом; 2 - репликация хромосом с образованием сестринских хроматид, соединенных в области центромеры; 3 - у полюсов ядерной оболочки видны центросомы с астеральными лучами, идущими к центромерному участку хроматид, выстроенных по экватору, чтобы разъединить центромеры сестринских хроматид и развести их к разным полюсам; 4 - деспирализация хромосом, восстановление ядерной оболочки и образование клеточной перегородки с образованием идентичных материнской двух дочерних клеток с точно таким же, как в ней, числом хромосом
1 - три пары хромосом; 2 - репликация хромосом с образованием сестринских хроматид, соединенных в области центромеры; 3 - у полюсов ядерной оболочки видны центросомы с астеральными лучами, идущими к центромерному участку хроматид, выстроенных по экватору, чтобы разъединить центромеры сестринских хроматид и развести их к разным полюсам; 4 - деспирализация хромосом, восстановление ядерной оболочки и образование клеточной перегородки с образованием идентичных материнской двух дочерних клеток с точно таким же, как в ней, числом хромосом
В анафазе происходит деление и расхождение дочерних хромосом к полюсам клетки, то есть восстановление их должного числа. В телофазе, конечной стадии деления клетки, хромосомы приобретают тот же вид, что и до начала деления, а количество ДНК в каждом дочернем ядре уменьшается вдвое по сравнению с предыдущими стадиями. Таким образом, обе дочерние клетки содержат одинаковые количества цитоплазмы и идентичные наборы хромосом и готовы пройти митоз.
Постоянно делятся не все соматические клетки организма. В процессе эмбрионального развития происходит дифференциация органов и тканей, развивающихся по своему специфическому, генетически заложенному пути. Поэтому одни клетки превращаются в клетки мозга, другие - в клетки крови и т. д. Причем одни из них делятся постоянно, а другие лишь на определенной стадии развития или при необходимости, отвечая, например, за регенеративные
(восстановительные) процессы.