Биосинтез клеточной стенки. Клетчатка в питании Биосинтез целлюлозы в растениях
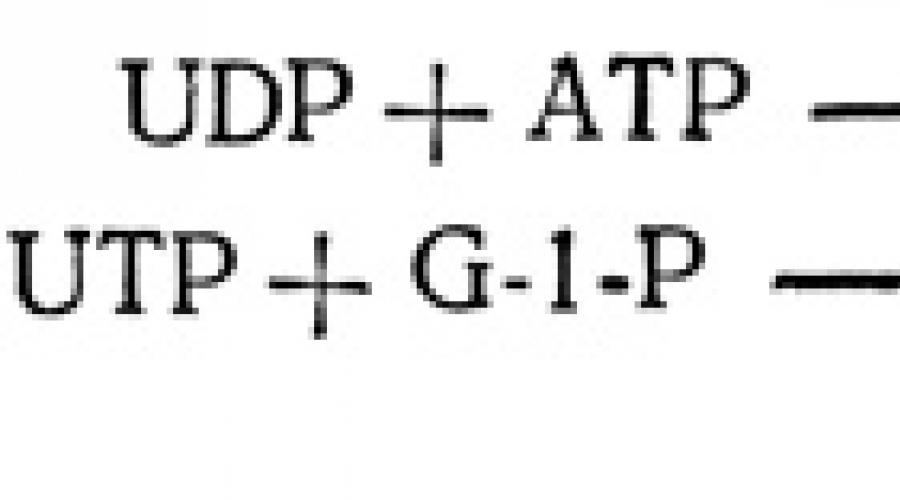
Читайте также
В цикле Кальвина - Бенсона образуется, как уже отмечалось выше, фруктозо-6-фосфат (F-6-P). Этот гексозофосфат может под действием специфических ферментов превращаться в другие фосфорилированные гексозы, а именно в глюкозо-6-фосфат (G-6-P) и глюкозо-1-фосфат (G-1-P). Легко происходит и обратное превращение.
Из этих трех гексозофосфатов строятся затем цепи углеводных молекул, используемых для транспорта, хранения и в реакциях синтеза. Чтобы такие превращения могли произойти, гексозофосфаты предварительно должны быть активированы. Это обычно достигается в результате их присоединения к нуклеотидам - сложным кольцевым структурам, сходным с адениловой кислотой АТР. Продуктом такой реакции присоединения оказываются нуклеотидные производные моносахаридов, или нуклеотидсахара. Чаще других встречаются уридиндифосфоглюкоза (UDPG), образующаяся в реакции между уридинтрифосфатом (UTP) и глюкозо-1-фосфатом (G-1-P). Сам UTP образуется непрямым путем, в результате переноса фосфатной группы от АТР к UDP (уридиндифосфату).

Нуклеотиды АТР и UTP присутствуют во всех клетках, потому что они используются наряду с другими нуклеотидами в синтезе ДНК и РНК.
Сахара транспортируются по растению в виде сахарозы - дисахарида, состоящего из остатков глюкозы и фруктозы (рис. 5.2). Сахароза образуется в реакции между UDPG и F-6-P:


Равновесие этой реакции сильно сдвинуто в сторону синтеза сахарозы, чем обеспечивается возможность накопления данного дисахарида в значительных концентрациях. Для последующего использования сахароза должна предварительно подвергнуться расщеплению: фермент инвертаза катализирует ее гидролиз с образованием свободной глюкозы и фруктозы.

Энергия гликозидной связи в такой реакции растрачивается впустую, распределяясь между двумя молекулами. Поэтому если глюкозе и фруктозе предстоит распад в процессе дыхания или участие (в качестве сырья) в синтезе полисахаридов, то они должны предварительно вновь подвергнуться фосфорилирова- нию за счет АТР. Процессы синтеза и распада сахарозы наглядно показывают, что часто анаболические и катаболические реакции (реакции синтеза и распада) идут по разным путям.
Синтез крахмала и целлюлозы
Длинные полимерные цепи крахмала и целлюлозы построены из одних и тех же элементарных звеньев - остатков глюкозы, только соединенных по-разному. Это структурное различие обусловливает то, что два рассматриваемых полимера глюкозы (глюканы) существенно различаются по своей природе; крахмал, например, легко переваривается в организме человека" а целлюлоза совсем не переваривается. Главное же их различие состоит в том, что 1-й и 4-й углеродные атомы двух соседних остатков глюкозы соединены у крахмала α-связями, а у целлюлозы (β-связями (рис. 5.3). Крахмал представлен двумя формами: линейным полимером, или амилозой, не содержащим никаких других связей, кроме α-1,4-гликозидных, и разветвленным полимером, или амилопектином, в котором наряду с α-1,4-гликозидными связями имеются и 1,6-связи. Различие в характере связей определяет и неодинаковое пространственное расположение полимерных цепей. Крахмал - главный запасной полисахарид растения. Он нерастворим в воде и отлагается слой за слоем в крахмальных зернах, содержащихся в хлоропластах (см. рис. 2.20) или в лишенных хлорофилла лейкопластах запасающих тканей стебля, корней и семян. Иногда клетки запасающей ткани оказываются буквально забиты крахмальными зернами, которые легко в них выявить, поскольку они способны окрашиваться иодом в синий цвет. Будучи нерастворим в воде, крахмал в отличие от сахарозы и от гексоз не вызывает в клетках осмотического эффекта (см. гл. 6). Поэтому образование крахмала в клетках листа в периоды интенсивного фотосинтеза предотвращает подавление последнего, происходящее в результате накопления продуктов фотосинтеза. В темноте крахмал постепенно снова гидролизуется с образованием глюкозофосфата, который затем превращается в сахарозу, транспортируемую в другие части растения.

Рис. 5.3. Структура крахмала (А) и целлюлозы (Б) (С изменениями по J. Bonner, A. W. Galston. 1952. Principles of Plant Physiology, San Francisco, W. H. Freeman and Co.) Обратите внимание, что химические формулы крахмала и целлюлозы одинаковы, различаются же эти полисахариды пространственной ориентацией их кислородных мостиков. А. Крахмал, главный запасной полисахарид растения, построен из двух хорошо различимых компонентов: амилозы с ее длинными неразветвленными цепями из глюкозных звеньев и амилопектина, состоящего из большого числа коротких разветвленных цепей. Б. Целлюлоза, главный компонент первичной клеточной стенки, существует в виде длинных полимерных цепей. Цепи объединяются в мицеллярные тяжи, а последние - в микрофибриллы. Микрофибриллы, достаточно крупные для того, чтобы их можно было рассмотреть при помощи электронного микроскопа, составляют "основу" и "уток" клеточной стенки
Исходным продуктом для синтеза крахмала служит аденозиндифосфоглюкоза (ADPG), образующаяся из АТР и G-1-P:

Молекула крахмала строится путем постепенного добавления одного глюкозного остатка за другим в реакции ADPG с предобразованной глюкозной цепью:
При низком содержании сахарозы крахмал расщепляется и. переводится в сахарозу. Однако сначала он расщепляется до остатков глюкозы и к каждому из них присоединяется остаток фосфорной кислоты, т. е. образуется G-1-P, чем обеспечивается сохранение энергии связи:
Этот G-1-P может затем использоваться для синтеза сахарозы, который мы описали выше. В семенах и в некоторых других органах, в которых одновременно идет расщепление больших количеств крахмала, он распадается до дисахарида мальтозы (G-G) под действием аα-амилазы. Мальтоза затем распадается до глюкозы, из которой (для транспорта) вновь синтезируется сахароза. На этом втором пути в отличие от первого энергия связи не сохраняется, поэтому здесь для превращения глюкозы в глюкозо-6-P требуется АТР.

Целлюлоза, самый распространенный на Земле углевод, служит главным компонентом первичной клеточной стенки. Молекулы ее строятся подобно тому, как строятся молекулы крахмала, с тем, оцнако, отличием, что роль донора глюкозы играет другое нуклеотидное производное моносахарида - гуанозин- дифосфоглюкоза (GDPG) - и что связь между мономерными.звеньями принадлежит не к α-, а к β-типу.

В некоторых случаях донором глюкозы для синтеза целлюлозы может быть и UDPG.
В организме высших растений целлюлоза расщепляется редко (если не считать распада, обусловленного деятельностью микробов). Два известных исключения из этого правила касаются клеток в отделительной зоне листа, образующейся перед сбрасыванием листьев, и сосудов ксилемы, у которых поперечные стенки растворяются. В отделительной зоне листа фермент целлюлаза разрушает клеточные стенки, расщепляя содержащуюся в них целлюлозу до отдельных мономерных единиц, т. е. до глюкозы. Клеточные стенки, ослабленные этим процессом, в конце концов разрываются, и лист сбрасывается.
Целлюлозные микрофибриллы в клеточной стенке скреплены при помощи матрикса из смешанных полисахаридных цепей, главным образом ксилоглюканов и арабиногалактанов (см. рис. 2.31). (Ксилоза и арабиноза - пятиуглеродные сахара (пентозы), а галактоза - гексоза, родственная глюкозе.) Эти полисахариды синтезируются также из предшественников, нуклео- тидсахаров, преимущественно в диктиосомах. Отшнуровывающиеся от диктиосом пузырьки в конце концов сливаются с плазмалеммой и таким путем передают свое содержимое формирующейся клеточной стенке.
Итак, все полисахариды легко переходят один в другой, но синтез их всегда идет через стадию нуклеотидсахаров, тогда как распад совершается более прямым путем.
В большинстве продуктов растительного происхождения содержатся так называемые балластные вещества, не перевариваемые в желудочно-кишечном тракте,- клетчатка и пектин. Однако они необходимы человеку. Если пища бедна ими, возникают атония кишечника и запоры. Таким образом, клетчатка является регулятором двигательной функции кишечника.
Клетчатка улучшает работу кишечника, замедляет гнилостные процессы и газообразование, уменьшает всасывание некоторых вредных веществ. Так, например, для профилактики профессиональных заболеваний у работающих с солями металлов и с радиоактивными веществами рекомендуется употреблять в пищу красную смородину , которая содержит много пектинов.
Полисахарид: Клетчатка
Клетчатка (целлюлоза) - полисахарид, составляющий главную массу клеточных стенок растений. Клетчатка нерастворима в воде, она лишь набухает в ней. Клетчатка составляет более 50% древесины. В волокнах хлопка она составляет более 90%. При кипячении с крепкой серной кислотой клетчатка нацело превращается в глюкозу. При более слабом гидролизе из клетчатки получается целлобиоза.
В молекуле клетчатки остатки целлобиозы связаны гликозидными связями в виде длинной цепочки. Молекулярная масса клетчатки точно не установлена. Полагают, что клетчатка не является индивидуальным веществом, а представляет собой смесь гомологичных веществ. Молекулярные массы клетчатки, полученной из различных источников, весьма сильно колеблются: хлопок - 330 000 (в цепочке 2020 гликозидных остатков); рами - 430 000 (2660 остатков), еловая древесина - 220 000 (1360 остатков). С помощью рентгеноструктурного анализа установлено, что молекулы клетчатки имеют нитевидную форму. Эти нитевидные молекулы соединяются в пучки - мицеллы. Каждая мицелла состоит приблизительно из 40-60 молекул клетчатки.
Соединение отдельных молекул клетчатки в мицеллы происходит благодаря водородным связям, которые осуществляются как за счет водородных атомов гидроксильных групп клетчатки, так и за счет адсорбированных клетчаткой молекул воды. В клеточных стенках растений мицеллы клетчатки связаны водородными связями с различными гетерополисахаридами. Например, у белого клена ими являются соединенные между собой гликозидными связями ксилоглюкан, состоящий из остатков глюкозы, ксилозы, галактозы и фукозы; арабиногалактан, построенный из остатков арабинозы и галактозы; рамногалактуронан, образованный остатками галактуроновой кислоты и рамнозы. Кроме того, имеются данные о том, что в построении клеточной стенки растений, особенно на ранних этапах ее образования, принимает участие также особый, богатый оксипролином гликопротеид экстензин. При одревеснении клеточных стенок в них накапливается также лигнин.
Клетчатка не переваривается в желудочно-кишечном тракте человека. Она, переваривается лишь жвачными животными, в желудке которых имеются особые бактерии, гидролизующие клетчатку с помощью выделяемого ими фермента целллюлазы.
Гемицеллюлозы (полуклетчатки). Под этим названием объединяют большую группу высокомолекулярных полисахаридов, не растворяющихся в воде, но растворимых в щелочных растворах. Гемицеллюлозы содержатся в значительном количестве в одревесневших частях растений: соломе, семенах, орехах, древесине, кукурузных початках. Большое количество гемицеллюлоз содержится в отрубях. Гемицеллюлозы гидролизуются кислотами легче, чем клетчатка. При этом они образуют маннозу, галактозу, арабинозу или ксилозу и поэтому соответственно носят названия - маннаны, галактаны и пентозаны (арабан или ксилан).
В растениях в процессе фотосинтеза образуются не только фосфорные эфиры сахаров или простые сахара, но и более сложные формы углеводов - сахароза, крахмал, клетчатка. Распад сложных форм углеводов до простых протекает тоже очень быстро. Это наблюдается, например, при прорастании семян, старении вегетативных органов и др. Образующиеся при распаде простые сахара или их фосфорные эфиры оттекают в репродуктивные органы, где из них вновь синтезируются сложные углеводы. И, наконец, в растениях очень легко идут процессы взаимных превращений углеводов.
Взаимопревращение моносахаридов проходит через фосфорные эфиры сахаров или их уридиндифосфатпроизводные (УДФ-производные). УДФ-производные сахаров представляют собой тот или иной сахар, соединенный через два остатка фосфорной кислоты с уридином, например:
Рис.1. Уридиндифосфатглюкоза
Синтез сахарозы
Сахароза является наиболее важным из образующихся в растениях лигосахаридов, в форме которого связанный углерод и энергия транспортируются по всему растению. Она состоит из α-D-глюкозного остатка в его пиранозной форме, связанного гликозидной связью с β-D-фруктозой в фуранозной форме. Поскольку аномерный углеродный атом обоих моносахаридов участвует в образовании гликозидной связи, их полуацетальные группы заблокированы и ни одно из колец не может открыться. Таким образом, сахароза – это невосстанавливающий сахар (не восстанавливает реактивы Фелинга и Бенедикта), и за исключением ее чрезвычайной чувствительности к кислотному гидролизу, она химически инертна. При нагревании с кислотами, или под действием сахаразы (инвертазы) сахароза гидролизуется, образуя инвертный сахар – смесь глюкозы и фруктозы. Сахароза хорошо растворима в воде и обладает сладким вкусом.
Сахароза используется как продукт питания, а также в производстве
поверхностно-активных веществ (эфиры сахарозы с высшими кислотами). Основным
источником получения сахарозы является сахарная свекла, содержащая до 23 %
сахарозы, и сахарный тростник, стеб-ли которого содержат 10–18 % сахарозы. В
настоящее время установлено, что сахароза синтезируется не только в
хлоропластах, но и в цитоплазме фотосинтезирую-щих клеток из УДФ-глюкозы и
фруктозо-6-фосфата, возникшего из
дигидроксиацетонфосфата. Это вещество образуется при фотосинтезе в хлоропластах
и затем поступает в цитоплазму. В нефотосинтезирующих тканях (например, в
эндосперме прорастающих бобов клещевины) образование сахарозы из УДФ-глюкозы и
фруктозо-6-фосфата также происходит в цитоплазме клеток.
Сахароза (тростниковый, свекловичный сахар) – самый распространенный в природе дисахарид. В растениях он образуется из глюкозы и фруктозы. На первом этапе идет фосфорилирование глюкозы:
Глюкоза + АТФ → глюкоза-6-фосфат + АДФ,
затем глюкозо-6-фосфат изолируется в глюкозо-1-фосфат. Глюкозо-1-фосфат соединяется с УТФ, в результате чего отщепляется пирофосфатная кислота и образуется соединение глюкозы с уридиндифосфорной кислотой (УДФ) - уридиндифосфатглюкоза.
Одновременно идет фосфорилирование фруктозы под действием фермента фруктокиназы с участием АТФ:
фруктоза + АТФ → фруктозо-6-фосфат + АДФ.
После этого происходит взаимодействие УДФ-глюкозы с фруктозо-6-фосфатом с участием фермента сахарозофосфат-УДФ-глюкозилтрансферазы. Наконец, образовавшийся сахарозо-6-фосфат под действием фермента фосфатазы гидролизуется с образованием свободной сахарозы. Таким образом, для биосинтеза одной молекулы сахарозы необходимы 3 макроэргические фосфатные связи, эта реакция необратима. В нефотосинтезирующих тканях некоторых растений, например, в корнеплодах сахарной свеклы, клубнях картофеля и других сахароза может образоваться из свободной фруктозы:
УДФ-глюкоза + фруктоза ↔ сахароза + УДФ.
Реакция катализируется ферментом сахарозо-УДФ-глюкозилтрансферазой и в зависимости от условий может быть направлена как в сторону синтеза, так и в сторону расщепления сахарозы.
Синтез крахмала
Крахмал – запасной полисахарид растений, причем он может храниться в растении долго, или расходоваться достаточно быстро. На длительное время он запасается во многих семенах, клубнях и корневищах и используется только тогда, когда эти органы прорастают. На короткое время крахмал образуется в хлоропластах в период быстрого фотосинтеза, а в последующий темновой период он расходуется и оттекает из листьев в форме сахарозы. Крахмал всегда образуется и запасается в виде крахмальных зерен, находящихся в пластидах – хлоропластах либо амилопластах. Крахмальные зерна – это высокоорганизованные структуры, форма и размер которых очень разнообразны, но часто характерны для данного вида растения. Форма зерен может быть сферической, яйцевидной, чечевицеобразной или неправильной; размер может колебаться 1 до 100 мкм. Наиболее крупные крахмальные зерна у картофеля, а самые мелкие у риса и гречихи. Крахмальные зерна содержат до 20 % воды (из которых 10 % химически связано с крахмалом) и ряда концентрических слоев крахмала. Крахмальные зерна образуются путем наслаивания вновь образованных слоев на ранее существующие.
Содержание минеральных веществ в крахмале очень невелико – 0,2–0,7 %, они представлены в основном фосфорной кислотой. В крахмале найдены некоторые высокомолекулярные жирные кислоты (пальмитиновая, стеариновая и др.), содержание которых достигает 0,6 %. Крахмал представляет собой смесь двух полисахаридов – амилозы и амилопектина. Молекулы амилозы – это длинные неразветвленные цепи, содержащие от 100 до нескольких тысяч глюкопиранозных остатков, соединенных гликозидными связями. Изучение дифракции рентгеновских лучей показало, что молекулы амилозы имеют спиралевидную структуру диаметром 1,3 нм с шестью последовательными глюкозными остатками на один оборот спирали.
Молекулы амилозы растворимы в горячей воде, но образующийся раствор нестоек и затем происходит спонтанное осаждение амилозы, известное как ретроградация. Это происходит вследствие тенденции длинных и тонких амилозных молекул выстраиваться в линию рядом друг с другом и образовывать нерастворимые агрегаты с помощью водородных связей. Донором глюкозных остатков при биосинтезе амилозы может служить уридиндифосфатглюкоза-(УДФК). Для ее образования в реакционной среде необходимо наличие затравки, в качестве которой могут служить полисахариды, построенные всего лишь из 3-4 остатков глюкозы, соединенных α(1-4)-связями.
Остатки глюкозы переносятся на акцептор (затравку), где и происходит удлинение цепи. Реакция идет по схеме:
УДФГ + акцептор (Г)к -- УДФ + акцептор (Г)к + 1,
где Г - остатки глюкозы.
Фермент, катализирующий эту реакцию, называется УДФГ-крахмалглюкозилтрансферазой.
У большинства растений активным донором глюкозы является не УДФ-глюкоза, а аденозиндифосфатглкжозα(АДФГ). Реакция присоединения глюкозных остатков от АДФГ к низкомолекулярному акцептору идет аналогичным путем и катализируется ферментом АДФГ-крахмал-глюкозилтрансферазой. Синтез разветвленной молекулы амилопектина, имеющей α (1-6)-связи, происходит при помощи фермента α-глюкантрансферазы (Q-фермент). В синтезе крахмала участвуют и Д-фермент или глюкозилтрансфераза, образующий α(1-4)-связи и участвующий в образовании затравки.
Распад крахмала происходит при участии двух процессов - гидролиза и фосфоролиза.Гидролитический распад крахмала осуществляется под действием четырех ферментов класса гидролиз α-амилаза, катализирует расщепление α(1-4)-связи, причем связи разрываются беспорядочно. Конечный продукт такого распада - мальтоза, глюкоза, декстрины. Под действием β-амилазы происходит расщепление α (1-4)-связей с образованием остатков мальтозы. Фермент глюкоамилазы катализирует последовательное отщепление остатков глюкозы от молекулы крахмала. Амилопектин-1,6-глюкозидаза или R-фермент катализирует расщепление α(1-6)-связей в молекуле амилопектина, т. е. действует на точки ветвления.
Фосфоролиз - это присоединение фосфорной кислоты по месту разрыва глюкозидной связи между остатками моносахаридов в цепи полисахарида, при этом происходит образование глюкозо-1-фосфата. Эта реакция катализируется ферментомаглюконфосфорилазой, относящимся к классу трансфераз. Крахмал в растении может подвергаться очень быстрому распаду, так как ферменты распада находятся во всех органах растения.
Синтез целлюлозы
Целлюлоза не растворима в воде, она лишь набухает в ней. При кислотном гидролизе (кипячение в серной кислоте) превращается в глюкозу, при более слабом – в целлобиозу. С помощью рентгеноструктурного анализа установлено, что молекула целлюлозы имеет нитевидную форму. Эти нитевидные молекулы, благодаря водородным связям, соединяются пучками по 40–60 штук в мицеллу. В клеточных стенках растений мицеллы целлюлозы связаны во-дородными связями с различными гетерополисахаридами. Например, у белого клена ими являются соединенные между собой гликозидными связями ксилоглюкан, арабиногалактан, рамногалактурон. Кроме того, имеются данные о том, что в построении клеточной стенки растений принимает участие особый, богатый оксипролином гликопротеид экстензин.
Целлюлоза построена из остатков β-глюкозы. В биосинтезе целлюлозы принимает участие не свободная глюкоза, а ее ГДФ-производное - гуанозиндифосфатглюкоза при участии фермента целлюлозосинтетазы по схеме:
ГДФ - глюкоза + (глюкоза) к→ ГДФ + (глюкоза) к + 1
Распад целлюлозы идет преимущественно гидролитическим путем под действием фермента целлюлазы до дисахарида целлобиозы.
Транспорт углеводов осуществляется в виде сахарозы. В процессе фотосинтеза образуется много углеводов, и в этой связи большое значение имеет отток ассимилятов в другие части клетки из хлоропластов. Проникновение через мембрану хлоропластов фосфорилированных гексоз и сахарозы затруднено, наиболее легко через мембраны хлоропластов проникают триозофосфаты (ФГА и ФДА). Предполагается, что образующиеся сложные углеводы распадаются на триозофосфаты и в таком виде передвигаются в цитоплазму, где могут служить материалом для ресинтеза гексоз, сахарозы, крахмала.
Межклеточный паренхимный транспорт осуществляется двумя путями - по плазмодесмам (симпласту) или по свободному пространству (аппопласту). Сахароза, образовавшаяся в клетках мезофилла листа, десорбируется в аппопласт. Выходя из паренхимных клеток в аппопласт, сахароза расщепляется инвертазой на гексозы. Гексозы передвигаются по аппопласту к передаточным клеткам проводящих пучков по градиенту концентраций. При соприкосновении с передаточными клетками флоэмы они снова превращаются в сахарозу. Далее происходит загрузка ситовидных трубок, сахароза поступает против градиента концентраций, и требуется расход энергии (АТФ).
Предполагается, что сахароза преодолевает мембрану с помощью переносчика в комплексе с протоном. При этом благодаря работе Н + -АТФ-азы ионы Н + выкачиваются из клеток флоэмы, а затем поступают обратно по градиенту рН, увлекая за собой сахарозу против градиента ее концентрации. Основной транспортной формой углеводов по флоэме служит сахароза (С 12 Н 22 О 11). У некоторых видов наряду с сахарозой транспортной формой углеводов служат олигосахара (рафиноза, стахиоза), а также некоторые спирты.