Создатель хромосомной теории наследственности ученый. Хромосомная теория наследственности
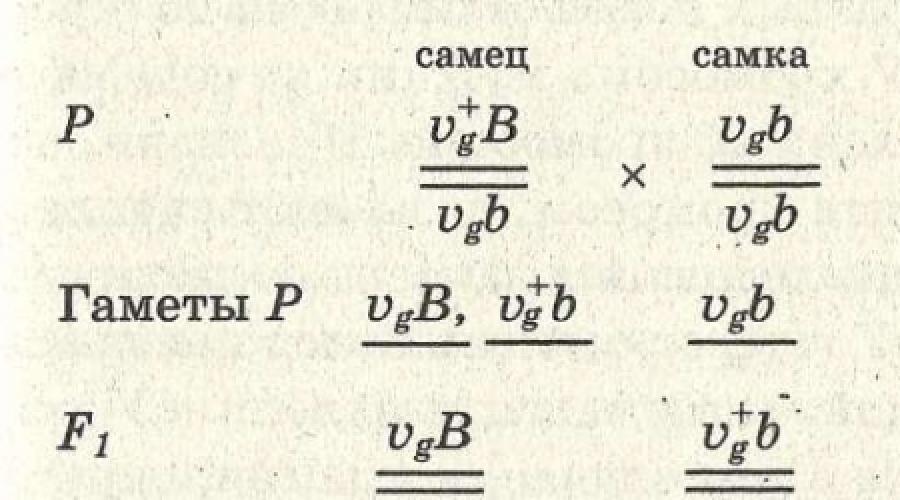
Читайте также
Заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, т.е. преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Х. т. н. возникла в начале 20 в. на основе клеточной теории и использования для изучения наследственных свойств организмов гибридологического анализа.
В 1902 У. Сеттон в США, обративший внимание на параллелизм в поведении хромосом и менделевских т. н. "наследственных факторов", и Т. Бовери в Германии выдвинули хромосомную гипотезу наследственности, согласно которой менделевские наследственные факторы (название впоследствии генами) локализованы в хромосомах. Первые подтверждения этой гипотезы были получены при изучении генетического механизма определения пола у животных, когда было выяснено, что в основе этого механизма лежит распределение половых хромосом среди потомков. Дальнейшее обоснование Х. т. н. принадлежит американскому генетику Т. Х. Моргану , который заметил, что передача некоторых генов (например, гена, обусловливающего белоглазие у самок дрозофилы при скрещивании с красноглазыми самцами) связана с передачей половой Х-хромосомы, т. е. что наследуются признаки, сцепленные с полом (у человека известно несколько десятков таких признаков, в том числе некоторые наследственные дефекты — дальтонизм, гемофилия и др.).
Доказательство Х. т. н. было получено в 1913 американским генетиком К. Бриджесом, открывшим нерасхождение хромосом в процессе мейоза у самок дрозофилы и отметившим, что нарушение в распределении половых хромосом сопровождается изменениями в наследовании признаков, сцепленных с полом.
С развитием Х. т. н. было установлено, что гены, расположенные в одной хромосоме, составляют одну группу сцепления и должны наследоваться совместно; число групп сцепления равно числу пар хромосом, постоянному для каждого вида организмов; признаки, зависящие от сцепленных генов, также наследуются совместно. Вследствие этого закон независимого комбинирования признаков должен иметь ограниченное применение; независимо должны наследоваться признаки, гены которых расположены в разных (негомологичных) хромосомах. Явление неполного сцепления генов (когда наряду с родительскими сочетаниями признаков в потомстве от скрещиваний обнаруживаются и новые, рекомбинантные, их сочетания) было подробно исследовано Морганом и его сотрудниками (А. Г. Стёртевантом и др.) и послужило обоснованием линейного расположения генов в хромосомах. Морган предположил, что сцепленные гены гомологичных хромосом, находящиеся у родителей в сочетаниях и , в мейозе у гетерозиготной формы ® могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами и последующему соединению разорванных концов в новом сочетании: Реальность этого процесса, названного перекрестом хромосом, или кроссинговером, была доказана в 1933 нем, учёным К. Штерном в опытах с дрозофилой и американскими учёными Х. Крейтономи Б. Мак-Клинток — с кукурузой. Чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Зависимость частоты кроссинговера от расстояний между сцепленными генами была использована для построения генетических карт хромосом. В 30-х гг. 20 в. Ф. Добржанский показал, что порядок размещения генов на генетических и цитологических картах хромосом совпадает.
Согласно представлениям школы Моргана, гены являются дискретными и далее неделимыми носителями наследственной информации. Однако открытие в 1925 советскими учёными Г. А. Надсоном и Г. С. Филипповым, а в 1927 американским учёным Г. Мёллером влияния рентгеновских лучей на возникновение наследственных изменений (мутаций) у дрозофилы, а также применение рентгеновских лучей для ускорения мутационного процесса у дрозофилы позволили советским учёным А. С. Серебровскому, Н. П. Дубинину и др. сформулировать в 1928—30 представления о делимости гена на более мелкие единицы, расположенные в линейной последовательности и способные к мутационным изменениям. В 1957 эти представления были доказаны работой американского учёного С. Бензера с бактериофагом Т4. Использование рентгеновских лучей для стимулирования хромосомных перестроек позволило Н. П. Дубинину и Б. Н. Сидорову обнаружить в 1934 эффект положения гена (открытый в 1925 Стёртевантом), т. е. зависимость проявления гена от места расположения его на хромосоме. Возникло представление о единстве дискретности и непрерывности в строении хромосомы.
Х. т. н. развивается в направлении углубления знаний об универсальных носителях наследственной информации — молекулах дезоксирибонуклеиновой кислоты (ДНК). Установлено, что непрерывная последовательность пуриновых и пиримидиновых оснований вдоль цепи ДНК образует гены, межгенные интервалы, знаки начала и конца считывания информации в пределах гена; определяет наследственный характер синтеза специфических белков клетки и, следовательно, наследственный характер обмена веществ. ДНК составляет материальную основу группы сцепления у бактерий и многих вирусов (у некоторых вирусов носителем наследственной информации является рибонуклеиновая кислота); молекулы ДНК, входящие в состав митохондрий, пластид и др. органоидов клетки, служат материальными носителями цитоплазматической наследственности.
Х. т. н., объясняя закономерности наследования признаков у животных и растительных организмов, играет важную роль в с.-х. науке и практике. Она вооружает селекционеров методами выведения пород животных и сортов растений с заданными свойствами. Некоторые положения Х. т. н. позволяют более рационально вести с.-х. производство. Так, явление сцепленного с полом наследования ряда признаков у с.-х. животных позволило до изобретения методов искусственного регулирования пола у тутового шелкопряда выбраковывать коконы менее продуктивного пола, до разработки способа разделения цыплят по полу исследованием клоаки — отбраковывать петушков и т.п. Важнейшее значение для повышения урожайности многих с.-х. культур имеет использование полиплоидии. На знании закономерностей хромосомных перестроек основывается изучение наследственных заболеваний человека.
Закономерности, открытые школой Моргана, а затем подтвержденные н углубленные на многочисленных объектах, известны под общим назва-нием хромосомной теории наследственности.
Основные положения ее следующие:
1. Гены находятся в хромосомах; каждая хромосома представляет со-бой группу сцепления генов; число групп сцепления у каждого вида рав-но числу пар хромосом.
2. Каждый ген в хромосоме занимает определенное место (локус); гены в хромосомах расположены линейно.
3. Между гомологичными хромосомами происходит обмен аллельными генами.
4. Расстояние между генами (локусами) в хромосоме пропорциональ-но числу кроссинговера между ними.
Основоположник теории Томас Гент Морган, американский генетик, нобелевский лауреат, выдвинул гипотезу об ограничении законов Менделя.
В экспериментах он использовал плодовую мушку-дрозофилу, обладающую важными для генетических экспериментов качествами: неприхотливостью, плодовитостью, небольшим количеством хромосом (четыре пары), множеством четко выраженных альтернативных признаков.
Морган и его ученики установили следующее:
- Гены, расположенные в одной хромосоме, наследуются совместно или сцепленно.
- Группы генов, расположенных в одной хромосоме, образуют группы сцепления. Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и п+1 у гетерогаметных особей.
- Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссинговера возникают гаметы, хромосомы которых содержат новые комбинации генов.
- Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера. За единицу расстояния между генами принимают 1 морганиду (1% кроссинговера) или процент появления кроссоверных особей. При значении этой величины в 10 морганид можно утверждать, что частота перекреста хромосом в точках расположения данных генов равна 10% и что в 10% потомства будут выявлены новые генетические комбинации.
- Для выяснения характера расположения генов в хромосомах и определения частоты кроссинговера между ними строят генетические карты. Карта отражает порядок расположения генов в хромосоме и расстояние между генами одной хромосомы. Эти выводы Моргана и его сотрудников получили название хромосомной теории наследственности . Важнейшими следствиями этой теории являются современные представления о гене как о функциональной единице наследственности, его делимости и способности к взаимодействию с другими генами.
Пример сцепленного наследования:
- Vg - нормальные крылья дрозофилы;
- vg - зачаточные крылья;
- ВВ - серая окраска тела;
- bb - темная окраска тела.
Запись в хромосомном выражении:
В данном случае правило единообразия гибридов первого поколения соблюдается. В соответствии со вторым и третьим законами Менделя следовало ожидать при последующем анализирующем скрещивании по 25% каждого из возможных фенотипов (серых, длиннокрылых мух, серых короткокрылых, черных длиннокрылых и черных короткокрылых). Однако опыты Моргана не дали таких результатов. При скрещивании рецессивной по обоим признакам самки VgVgbb с гибридным самцом из F1 образовалось 50% серых мух с короткими крыльями и 50% мух с черным телом и длинными крыльями:

Если же скрещивают дигибридную самку с гомозиготным рецессивным самцом, то в образуется потомство: 41,5% - серых с короткими крыльями, 41,5% - черных с длинными крыльями, 8,5% - серых с длинными крыльями, 8,5% - черных с короткими крыльями.
Данные результаты свидетельствуют о наличии сцепления генов и кроссинговере между ними. Так как в потомстве от второго скрещивания было получено 17% рекомбинантных особей, то расстояние между генами Vg и В равно 17%, или 17 морганидам.
Наследование, сцепленное с полом
Хромосомные наборы разных полов отличаются по строению половых хромосом. Y-хромосома мужчин не содержит многих аллелей, имеющихся в Х-хромосоме. Признаки, определяемые генами половых хромосом, называют сцепленными с полом. Характер наследования зависит от распределения хромосом в мейозе. У гетерогаметных полов признаки, сцепленные с Х-хромосомой и не имеющие аллеля в Y-хромосоме, проявляются даже в том случае, когда ген, определяющий развитие этих признаков, - рецессивен. У человека У-хромосома передается от отца к сыновьям, а Х-хромосома - к дочерям. Вторую хромосому дети получают от матери. Это всегда Х-хромосома. Если мать несет патологический рецессивный ген в одной из Х-хромосом (например, ген дальтонизма или гемофилии), но при этом сама не больна, то она является носительницей. В случае передачи этого гена сыновьям они могут родиться с данным заболеванием, ибо в У- хромосоме нет аллеля, подавляющего патологический ген. Пол организма определяется в момент оплодотворения и зависит от хромосомного набора образовавшейся зиготы. У птиц гетерогаметными являются самки, а гомогаметными - самцы. У пчел половых хромосом вообще нет. Самцы гаплоидны. Самки пчел диплоидны.

Основные положения хромосомной теории наследственности:
- каждый ген имеет в хромосоме определенный локус (место);
- гены в хромосоме расположены в определенной последовательности;
- гены одной хромосомы сцеплены, поэтому наследуются преимущественно вместе;
- частота кроссинговера между генами равна расстоянию между ними;
- набор хромосом в клетках данного типа (кариотип) является характерной особенностью вида.
Статья на конкурс «био/мол/текст»: В 2015 году исполняется 100 лет хромосомной теории наследственности . Ее основные положения были сформулированы Т. Морганом, А. Стёртевантом, Г. Мёллером и К. Бриджесом в книге «Механизм менделевской наследственности», вышедшей в Нью-Йорке в 1915 году. А позднее Томас Морган получил первую «генетическую» Нобелевскую премию - за открытие роли хромосом в наследственности. Юбилею хромосомной теории была посвящена международная конференция «Хромосома 2015», прошедшая в августе 2015 года в Новосибирском Академгородке. Нижеизложенный текст - это авторские комментарии к постеру об истории исследований хромосом , представленному на конференции, а теперь и на «Биомолекуле» - в самой «живой» конкурсной номинации «Наглядно о ненаглядном ».
Обратите внимание!
Более полную информацию можно найти в книге - Коряков Д.Е., Жимулев И.Ф. . Новосибирск: Изд-во СО РАН, 2009 г. - 258 с., ISBN 978-5-7692-1045-7
Спонсором номинации «Лучшая статья о механизмах старения и долголетия» является фонд «Наука за продление жизни ». Спонсором приза зрительских симпатий выступила фирма Helicon .
Спонсоры конкурса: Лаборатория биотехнологических исследований 3D Bioprinting Solutions и Студия научной графики, анимации и моделирования Visual Science .
Нижеизложенный текст - это краткие комментарии к постеру, а более полную информацию можно найти в книге: Коряков Д.Е., Жимулев И.Ф. Хромосомы. Структура и функции . Новосибирск: Изд-во СО РАН, 2009 г. - 258 с., ISBN 978-5-7692-1045-7.
Нажмите на изображение, чтобы увеличить (откроется в отдельном окне).
Генетическая роль хромосом
Каждый организм воспроизводит лишь себе подобных, и даже в мелких чертах внешности и поведения детей можно увидеть сходство с их родителями. Первый шаг на пути к пониманию, почему так происходит, сделал монах из австрийского города Брюнн (сейчас это чешский Брно) Г. Мендель (G. Mendel ). В 1865 году на заседании Брюннского общества испытателей природы он сделал доклад под названием «Опыты над растительными гибридами » (Versuche über Pflanzen-Hybriden ), а в 1866 году опубликовал его в сборнике трудов этого общества. Монах-естествоиспытатель описал результаты скрещиваний разных форм гороха и предположил наличие особых факторов, от которых зависят внешние признаки растения. Закономерности наследования этих факторов позднее были названы законами Менделя . Однако современники не поняли значения этого открытия и забыли про него, и лишь в 1900 году Г. де Фриз (H. de Vries , Нидерланды), К. Корренс (C. Correns , Германия) и Э. Чермак (E. Tschermak , Австрия) независимо друг от друга переоткрыли законы Менделя.
Задолго до всех этих исследований, которые сейчас бы назвали генетическим анализом, ученые, занимавшиеся ботаникой, зоологией, эмбриологией, гистологией и физиологией, заложили основу цитогенетики - науки о хромосомах. В разных статьях и книгах приоритет открытия хромосом отдан разным людям, но чаще всего годом их открытия называют 1882, а их первооткрывателем - немецкого анатома В. Флемминга (W. Flemming ). Однако справедливее было бы сказать, что он не открыл хромосомы, а лишь собрал и упорядочил в своей фундаментальной книге «Клеточное вещество, ядро и деление клетки » (Zellsubstanz, Kern und Zellteilung ) всё, что было известно о них на тот момент. Сам же термин «хромосома» ввел в науку немецкий гистолог Х. Вальдейер (H. Waldeyer ) в 1888 году, и в буквальном переводе термин означает «окрашенное тело».
Сейчас сложно сказать, кто сделал первое описание хромосом. В 1842 году швейцарский ботаник К. Нэгели (C. Nägeli ) опубликовал работу, в которой изобразил некие тельца, возникающие на месте ядра во время деления клетки при образовании пыльцы у лилии и традесканции. Возможно, это и были первые рисунки хромосом. Первое (1873 год) подробное описание митоза у плоского червя Mesostoma ehrenbergii принадлежит, как считают, немецкому зоологу А. Шнайдеру (F.A. Schneider ). Он описал не просто отдельные стадии митоза, которые видели и до него, а всю последовательность сложных изменений ядра: возникновение на его месте нитевидных телец, их расхождение в противоположные стороны и формирование новых ядер в дочерних клетках. Другой тип деления - мейоз - впервые подробно описал Э. ван Бенеден (E. van Beneden , Бельгия) в 1883 году, наблюдая за образованием гамет у аскариды. Он обнаружил, что в мейозе число хромосом уменьшается вдвое, а при оплодотворении восстанавливается, и, несмотря на различие в размерах, мужская и женская гаметы привносят в зиготу равное число хромосом.
* - Немного о месте и предназначении мобильных генетических элементов в про- и эукариотических геномах: «Мобильные генетические элементы прокариот: стратификация „общества“ бродяжек и домоседов », «Геном человека: полезная книга, или глянцевый журнал? », «„Мусорная“ ДНК управляет эволюцией млекопитающих? » - Ред.
Еще одним вариантом обмена участками является сестринский хроматидный обмен (СХО). Если при кроссинговере обмениваются хроматиды разных хромосом, то в случае СХО обмениваются хроматиды внутри одной хромосомы. Впервые СХО увидел американский генетик Д. Тейлор (J. Taylor ) в 1958 году.
С кроссинговером, хоть и неоднозначно, но связано формирование в профазе мейоза особой структуры из пары гомологичных хромосом - синаптонемного комплекса . Он был открыт в 1956 году независимо двумя американскими цитологами: М. Мозесом (M. Moses ) у речного рака и Д. Фоцеттом (D. Fawcett ) у мыши.
Многообразие хромосом
Если понимать под хромосомами любые носители наследственной информации, то они исключительно разнообразны по размеру, форме, внешнему виду, составу и числу. Хромосомы вирусов и бактерий могут быть кольцевыми и линейными. Хромосомы хлоропластов и митохондрий имеют кольцевую форму. Ядерные хромосомы эукариот имеют линейную форму, и именно они в виде телец X- и V-образной формы обычно приходят на ум при упоминании хромосом. Их называют митотическими или метафазными , поскольку такой вид они имеют во время деления - митоза (а метафаза - это одна из его стадий).
В 1912 году российский ботаник и цитолог С.Г. Навашин показал, что метафазные хромосомы обладают индивидуальным набором признаков, включающим размер, соотношение длин плеч, наличие спутников и перетяжек. Используя положение центромеры или соотношение длин плеч, С.Г. Навашин предложил классификацию митотических хромосом, которую используют и по сей день: метацентрики, субметацентрики, акроцентрики и телоцентрики.
Число хромосом у разных видов организмов может варьировать в самых широких пределах: от двух (у пары видов растений и одного из австралийских муравьев) до 1440 у папоротника Ophioglossum reticulatum и даже 1600 у морской радиолярии Aulacantha scolymantha . У человека число хромосом составляет 46, и оно было определено только в 1955 году, а опубликовано в 1956 цитогенетиком китайского происхождения Д. Чио (J. Tjio ) в соавторстве со своим руководителем А. Леваном (A. Levan ) в Швеции. Несколькими месяцами позже число подтвердили британцы Ч. Форд (C. Ford ) и Д. Хамертон (J. Hamerton ). Количество хромосом человека пытались определить еще с конца XIX века. В разных случаях получались разные значения: 18, 24, 47 или 48, - и только в 1955 году убедились, что хромосом у человека 46. В честь этого события на здании Института генетики Университета шведского города Лунда (где это событие и случилось) в 2003 году была открыта мемориальная доска с изображением той самой метафазной пластинки, по которой и были посчитаны хромосомы. Любопытно, что число хромосом шимпанзе (48) было выяснено на 15 лет раньше.
Общепринято, что число хромосом у каждого вида живых организмов постоянно, и в подавляющем большинстве случаев так и есть. Однако у некоторых животных и растений существуют так называемые сверхчисленные , или добавочные , хромосомы. Все хромосомы основного набора называют A-хромосомами . Они присутствуют всегда, и потеря или добавление хотя бы одной из них ведет к серьезным последствиям. Добавочные же хромосомы называют B-хромосомами , и их главные особенности - необязательность наличия и непостоянство числа. Впервые сверхчисленные хромосомы были найдены Э. Уилсоном (E. Wilson , США) в 1906 году у клопа Metapodius terminalis .
Своеобразный тип хромосом, названный хромосомами типа «ламповых щеток » , можно видеть в профазе первого деления мейоза при формировании ооцитов у птиц, рыб, рептилий и земноводных. Впервые их упоминает в своей фундаментальной книге (1882) В. Флеминг, который обнаружил эти хромосомы у аксолотля . Название они получили за сходство с ершиком для чистки керосиновых ламп.
Совершенно особое место среди всех типов хромосом занимают политенные хромосомы , которые имеют вид длинных толстых шнуров с поперечными полосками. Их открыл французский эмбриолог Э. Бальбиани (E. Balbiani ) в 1881 году в ядрах клеток слюнных желез личинок комара Chironomus plumosus . Политенные хромосомы сыграли выдающуюся роль в развитии генетики, цитогенетики и молекулярной биологии. С их помощью была показана линейность расположения генов и однозначно доказана генетическая роль хромосом. На политенных хромосомах дрозофил был впервые описан хромосомный полиморфизм диких популяций. Именно на политенных хромосомах были открыты гены белков теплового шока - компонентов системы, охраняющей клетки всех организмов от стрессорных воздействий. Политенные хромосомы сыграли ключевую роль в исследовании системы дозовой компенсации у дрозофилы.
Эволюция хромосом и геномов
В современных цитогенетических исследованиях важную роль играет дифференциальная окраска . Впервые способность хромосом окрашиваться дифференциально (то есть неодинаково по длине) продемонстрировали англичане С. Дарлингтон (C. Darlington ) и Л. Ла Кур (L. La Cour ) в 1938 году. Другой важный метод исследования - это гибридизация in situ , которая позволяет определить положение любого фрагмента ДНК на хромосоме. В основе метода лежит способность нуклеиновых кислот образовывать двуцепочечные молекулы, как ДНК-ДНК, так и РНК-ДНК. Придумали этот метод в 1969 году Д. Голл (J. Gall ) и М. Пардью (M. Pardue ) из США и Х. Джон (H. John ), М. Бирнстил (M. Birnstiel ) и К. Джонс (K. Jones ) из Великобритании .
Комбинация этих методов дает возможность подробно исследовать эволюцию хромосом и геномов*, а неизменным спутником эволюционного процесса являются хромосомные перестройки . По мере эволюции вида в его хромосомах неизбежно возникают перестройки, которые меняют порядок генов по сравнению с предковым видом. Чем дальше виды уходят друг от друга, тем больше хромосомных перестроек их отличает, и тем больше меняется порядок генов. Известны разные типы перестроек: делеции (потеря), дупликации (удвоение) и транслокации (перемещение) участков хромосом, которые обнаружил К. Бриджес в 1916, 1919 и 1923 годах соответственно. Еще один тип - это инверсии (поворот участка хромосомы на 180°), описанные А. Стёртевантом в 1921 году. Кроме того, существует особый тип перестроек, называемый Робертсоновской транслокацией (или центрическим слиянием). Первым ее описал американец У. Робертсон (W. Robertson ) в 1916 году, сравнивая хромосомные наборы близких видов саранчи. Суть этой перестройки сводится к слиянию двух акроцентрических хромосом в одну метацентрическую или субметацентрическую. Существует и обратный процесс - центрическое разделение. В этом случае мета- или субметацентрическая хромосома делится на две акроцентрических.
* - На биомолекуле можно найти внушительную подборку статей, так или иначе затрагивающих эволюцию геномов и изменения генетического кода: «Вирусные геномы в системе эволюции », «Под „генную гармошку“ », «Аллополиплоидия, или как разные геномы научились жить под одной крышей », «Полные геномы галапагосских вьюрков наконец-то раскрыли механизмы их эволюции », «Как составлялся геном эукариот: эндосимбиоз VS. непрерывный горизонтальный перенос »; «Таинственный код нашего генома », «Эволюция генетического кода », «У истоков генетического кода: родственные души », «Такие разные синонимы » и др. - Ред.
Положение хромосом в ядре
В конце XIX века Т. Бовери выдвинул идею о том, что хромосомы в интерфазном ядре не перемешаны случайным образом, а каждая из них занимает свое пространство. В 1909 году для обозначения этого пространства он ввел термин «хромосомная территория ». Первые доказательства существования хромосомных территорий были получены лишь в 1982 году немецким исследователем Т. Кремером (T. Cremer ) с соавторами. Позднее они визуализировали эти территории с помощью флуоресцентных красителей разного цвета. Оказалось, что хромосомы крупного размера с гораздо большей вероятностью можно найти в периферической части ядра, тогда как мелкие сосредоточены в основном в центральной. Кроме этого, на периферии ядра расположены районы хромосом, обедненные генами. Районы же, обогащенные генами, наоборот, расположены ближе к центру ядра.
Состав хромосом. ДНК
Хромосомы представляют собой структуры, состоящие из сложного комплекса ДНК, РНК и белков. Такой комплекс называется хроматином .
ДНК как химическое вещество открыл и выделил в чистом виде молодой швейцарский исследователь Ф. Мишер (F. Miescher ), работая в 1868–1869 годах в университете немецкого города Тюбингена. Он изучал химический состав лейкоцитов, источником которых служил гной с бинтов из местной хирургической клиники. Ф. Мишер разработал метод разделения ядер и цитоплазмы клеток и анализировал состав ядер. Помимо белков и липидов он обнаружил вещество, которое назвал нуклеином (от слова nucleus - ядро), а сейчас оно известно как ДНК. То, что именно ДНК является носителем наследственной информации, первыми установили в 1944 году американцы О. Эйвери (O. Avery ), К. МакЛауд (C. MacLeod ) и М. МакКарти (M. McCarty ) в экспериментах по заражению мышей пневмококками.
Структуру молекулы ДНК в виде двойной спирали расшифровали в 1953 году Ф. Крик (F. Crick ), Д. Уотсон (J. Watson ), М. Уилкинс (M. Wilkins ) и Р. Франклин (R. Franklin ), работавшие в Великобритании. За это открытие первые три исследователя получили Нобелевскую премию в 1962 году (историю открытия увлекательно описал в книге «Двойная спираль » Джеймс Уотсон, очень рекомендуем - Ред. ). Среди лауреатов нет Розалинды Франклин, поскольку она умерла от рака за четыре года до этого. Известно, что молекула ДНК состоит из последовательности четырех типов нуклеотидов : аденина, тимина, гуанина и цитозина*. За разработку метода определения их последовательности (секвенирования ) в 1980 году Нобелевской премии были удостоены П. Берг (P. Berg , США), У. Гилберт (W. Gilbert , США) и Ф. Сэнгер (F. Sanger , Великобритания).
* - Помимо четырех «классических» нуклеотидов в ДНК находят и их эпигенетически модифицированные варианты: метилцитозин и метиладенин («Шестое ДНК-основание: от открытия до признания »). А для некоторых бактериофагов Bacillus subtilis описано включение в ДНК «РНК-ового» урацила - Ред .
Если вначале секвенирование было трудоемким процессом, который позволял за раз «прочитать» лишь небольшой фрагмент, то по мере развития технологии стало возможным определить, например, полную последовательность митохондриальной ДНК человека (1981 год). В 1990 году был запущен амбициозный проект с целью полного секвенирования человеческого генома, а первый результат был представлен в 2001 году (биомолекула: «Геном человека: как это было и как это будет »). При этом секвенирование одного генома обошлось в колоссальную сумму - сотни миллионов долларов. Но технологии не стоят на месте, и появление новых методов позволило снизить затраты в тысячи раз*. Теперь секвенирование целого генома стало рядовым событием, и в 2009 году был запущен проект «Genome 10K». Его цель - это секвенирование и полная «сборка» в хромосомы 10 тысяч геномов животных.
* - «Закон» Мура прямо таки обречен на достижение конечных точек в разных науках (куда только его удалось притянуть). Биология даже обогнала электронику: постепенное падение стоимости секвенирования в 2007-м ушло в крутое пике, приближая эру рутинного чтения геномов в сельских фельдшерских пунктах по полисам ОМС. Правда, в обозримой перспективе всё же придется раскошелиться - долларов на 1000 плюс транспортные расходы: «Технология: 1,000 $ за геном ». Но и о таком могли лишь мечтать до появления новых методов секвенирования ДНК: «454-секвенирование (высокопроизводительное пиросеквенирование ДНК) ». И для понимания базовых (на уровне клетки) процессов развития организма и победы над онкозаболеваниями мечтать есть еще о чём: «Секвенирование единичных клеток (версия - Metazoa) » - Ред.
Новые технологии позволили развиться такому направлению, как исследование древней ДНК (биомолекула: «Древняя ДНК: Привет из прошлого »). Стало возможным выделять ДНК из костей возрастом десятки тысяч лет, и в 2008 году, например, был секвенирован митохондриальный геном неандертальца. Исследование древней ДНК, да и всю современную молекулярную биологию невозможно представить без использования ПЦР - полимеразной цепной реакции . За ее открытие американец К. Муллис (K. Mullis ) получил в 1993 году Нобелевскую премию .
Состав хромосом. Белки́
ДНК в хромосомах претерпевает несколько последовательных уровней упаковки, и на самом первом уровне двойная спираль ДНК оборачивается вокруг белковой глобулы, образуя нуклеосому (биомолекула: «Катится, катится к ДНК гистон »). В состав глобулы входят четыре типа белков, называемых гистонами . В 1982 году английский молекулярный биолог А. Клюг (A. Klug ) получил Нобелевскую премию за расшифровку трехмерной структуры нуклеосом. Косвенно нуклеосомы отмечены еще одной Нобелевской премией - в 1910 году ее получил немецкий биохимик А. Коссель (A. Kossel ) за изучение химического состава веществ, образующих ядро клетки, и в том числе за открытие гистонов.
C-концевые части молекул гистонов плотно свернуты, а N-концевые не имеют определенной структуры и свободно расходятся в стороны. В 1963–1964 годах было обнаружено, что некоторые аминокислотные остатки в гистонах могут быть ковалентно модифицированы, то есть ацетилированы или метилированы. Сейчас список модификаций значительно расширился, к остаткам аминокислот могут быть присоединены как относительно простые группы - метильная, ацетильная, фосфатная, - так и сложные крупные молекулы: биотин, олигопептиды или цепочки ADP-рибозы. Модификации появляются в основном на N- и, в гораздо меньшей степени, на С-концевой частях молекул гистонов.
Согласно теории гистонового кода , модификации, которые присутствуют на нуклеосомах в данном участке хроматина, не случайны, а «кодируют» какой-либо процесс. Такую точку зрения сформулировали в 2000–2001 годах Б. Штраль (B. Strahl , США), С. Эллис (C. Allis , США) и Т. Йенувайн (T. Jenuwein , Австрия). Схематично процесс работы гистонового кода можно составить из трех этапов. На первом этапе работают ферменты, которые производят модификацию определенных остатков в гистонах. На втором этапе с модифицированными аминокислотами связываются белки, имеющие для этой цели специальные домены. Каждый из доменов подходит только к «своей» модификации. На последнем же этапе эти связавшиеся белки привлекают другие белковые комплексы, запуская тем самым какой-то процесс.
* - О блестящих перспективах и отрезвляющих сомнениях в области применения ИПСК: «Французским исследователям удалось омолодить клетки столетних людей », «Снежный ком проблем с плюрипотентностью ». - Ред.
Гетерохроматин
Одним из объектов исследования многообразных эпигенетических процессов является гетерохроматин . Его как более темные участки хромосом открыл в 1907 году немецкий цитолог С. Гутхерц (S. Gutherz ), а термины «гетерохроматин» и «эухроматин» ввел в 1928 году другой немецкий цитолог Э. Хайц (E. Heitz ). Если совсем кратко, то эухроматин - это части хромосом, в которых расположено подавляющее большинство генов, тогда как гетерохроматин - это в основном районы с некодирующей ДНК, состоящей из коротких многократно повторенных последовательностей. Кроме этого, эу- и гетерохроматин различаются временем репликации в течение S-фазы клеточного цикла. Первым это отличие описал в 1959 году А. Лима-де-Фария (A. Lima-de-Faria , США), исследуя процесс репликации ДНК в семенниках у саранчи Melanoplus differentialis . Он показал, что гетерохроматин и начинает, и заканчивает репликацию своей ДНК позже эухроматина.
Важным свойством гетерохроматина является способность инактивировать помещенные в него эухроматиновые гены. Это явление называется эффектом положения мозаичного типа . Оно было обнаружено в 1930 году Г. Мёллером у дрозофилы. В результате хромосомной перестройки ген white попал в гетерохроматин. Этот ген отвечает за красный цвет глаз, а если он не работает, то глаза становятся белыми. У Г. Мёллера же получились мухи, глаза которых были ни красными, ни белыми, а пятнистыми, и у разных мух пятна были разной формы и размера. Это объясняется тем, что сам ген остается неповрежденным, а лишь случайным образом инактивируется в одних клетках глаза и работает в других.
Несмотря на многолетние исследования, процесс формирования гетерохроматина во многом до сих пор не ясен, особенно его самый первый этап. Предполагают, что ключевую роль в нём играет процесс, схожий с интерференцией РНК (биомолекула: «Обо всех РНК на свете, больших и малых »). За открытие этого явления два американца Э. Файр (A. Fire ) и К. Мелло (C. Mello ) получили Нобелевскую премию в 2006 году. Процесс интерференции сложен и многостадиен, но если не вдаваться в детали, то введение в клетку двухцепочечной РНК, гомологичной какому-либо гену, приводит к инактивации этого гена.
Теломеры
Интенсивное исследование теломер началось после того, как в 1978 году американцы Э. Блэкберн (E. Blackburn ) и Д. Голл секвенировали теломеру у инфузории Tetrahymena thermophila . Оказалось, что теломеры содержат последовательность из шести нуклеотидов, повторенную от 20 до 70 раз. В 1985 году К. Грейдер (C. Greider ) и Э. Блэкберн всё у той же инфузории открыли фермент, названный теломеразой , задачей которого является достраивание теломер. В 2009 году Э. Блэкберн , К. Грейдер и Д. Шостак (J. Szostak , США) получили Нобелевскую премию за исследование теломер и открытие фермента теломеразы (биомолекула: «„Нестареющая“ Нобелевская премия: в 2009 году отмечены работы по теломерам и теломеразе », «Старение - плата за подавление раковых опухолей? »).
Дозовая компенсация
Огромное число видов живых организмов, и человек в их числе, имеет негомологичные половые хромосомы, например, X и Y. При этом возникает необходимость в процессе, который называется дозовой компенсацией . Суть его заключается в следующем: поскольку число аутосом одинаково и у самцов, и у самок, то число аутосомных генов, а следовательно, и количество их продуктов, также будет одинаковым. А вот количество продуктов, синтезированных с генов, расположенных в половой хромосоме, у одного пола будет в 2 раза больше, чем у другого. Получается диспропорция, которую надо как-то регулировать, то есть уравнять «дозу генов». Решить эту проблему призвана система дозовой компенсации (биомолекула: «, США) выдвинули гипотезу, согласно которой у самок млекопитающих одна из двух X-хромосом инактивируется, и выбор ее случаен. Таким способом система дозовой компенсации млекопитающих уравнивает число работающих X-хромосом у разных полов: у самцов Х-хромосома всего одна, а у самок из двух только одна работает.
У дрозофилы природа изобрела другой механизм, противоположный по сути механизму млекопитающих: единственная X-хромосома самцов гиперактивируется и работает как две X-хромосомы самок. То, что суммарная активность двух копий какого-либо гена из X-хромосомы у самок и одной копии у самцов дрозофилы одинакова, было обнаружено еще на заре развития генетики. Это сделали К. Штерн в 1929 году и Г. Мёллер в 1931 году, так что дрозофила - это первый организм, у которого нашли дозовую компенсацию.
Ну и наконец...
Пара слов об открытии, которое не связано напрямую с хромосомами, но его очень активно используют, в том числе и для исследования разных сторон жизни хромосом. В 2008 году О. Шимомура (O. Shimomura ), М. Чалфи (M. Chalfie ) и Р. Циен (R. Tsien ) из США получили Нобелевскую премию за открытие, выделение и применение зеленого флуоресцирующего белка (GFP) медузы Aequorea victoria . С помощью молекулярных манипуляций можно соединить ген белка GFP с геном любого другого белка и получить химерный белок, который будет выполнять как свою исходную функцию, так и светиться зеленым цветом. Это дает возможность видеть, в каких клетках работает белок, в ядре или цитоплазме, в каких частях хромосом. Кроме зеленого (GFP) сейчас известны красный (RFP) и желтый (YFP) флуоресцирующие белки*.
* - О многообразии флуоресцентных белков и их применении в биологических исследованиях рассказывают материалы: «Флуоресцирующая Нобелевская премия по химии », «Флуоресцентные белки: разнообразнее, чем вы думали! », «„Нарисуем“ живую клетку ». А о биолюминесценции у наземных и морских организмов и работе люциферин-люциферазной системы - статьи: «Биолюминесценция: возрождение », «Микроскопическое свечение космического масштаба ». - Ред.
В клетках каждого организма находится определенное число хромосом. Генов в них очень много. У человека 23 пары (46) хромосом, генов около 100 000. Гены находятся в хромосомах. В одной хромосоме локализовано много генов. Хромосома со всеми находящимися в ней генами образует группу сцепления. Число групп сцепления равно гаплоидному набору хромосом. У человека 23 группы сцепления. Гены, находящиеся в одной хромосо- ме, сцеплены не абсолютно. Во время мейоза при конъюгации хромосом гомологичные хромосомы обмениваются частями. Это явление называют кроссинговером, который может произойти в любом участке хромосомы. Чем дальше расположены друг от друга локусы в одной хромосоме, тем чаще между ними может происходить обмен участками (рис. 76).
У мухи дрозофилы гены длины крыльев (V - длинные и v - короткие) и окраска тела (В - серая и b - черная) находятся в одной паре гомологичных хромосом, т.е. относятся к одной группе сцепления. Если скрестить муху, имеющую серый цвет тела и длинные крылья, с мухой черного цвета с короткими крыльями, то в первом поколении все мухи будут иметь серый цвет тела и длинные крылья (рис. 77).
В результате скрещивания дигетерозиготного самца с гомозиготной рецессивной самкой мухи будут похожи на родителей. Это происходит потому, что гены, находящиеся в одной хромосоме, наследуются сцепленно. У самца мухи дрозофилы сцепление полное. Если скрестить дигетерозиготную самку с гомозиготным рецессивным самцом, то часть мух будет похожа на родителей, а у
Рис. 76. Кроссинговер.
1 - две гомологичные хромосомы; 2 - их перекрест во время конъюгации; 3 - две новые комбинации хромосом.
другой части произойдет перекомбинация признаков. Такое наследование имеет место для генов одной группы сцепления, между которыми может произойти кроссинговер. Это пример неполного сцепления генов.
Основные положения хромосомной теории наследственности
. Гены находятся в хромосомах.
. Гены в хромосоме расположены линейно.

Рис. 77. Сцепленное наследование генов окраски тела и состояния крыльев у плодовой мухи.
Ген серого цвета (В) доминирует над геном черного цвета тела (b), ген длинных крыльев (V) - над геном коротких крыльев (v). В и V находятся в одной хромосоме.
а - полное сцепление генов вследствие отсутствия перекреста хромосом у самцов дрозофилы: РР - самка серая с длинными крыльями (BBVV) скрещена с черным короткокрылым самцом (bbvv); F 1 - серый самец с длинными крыльями (BbVv) скрещен с черной короткокрылой самкой (bbvv); F 2 - поскольку у самца не происходит кроссинговера, появятся два вида потомков: 50% - черных короткокрылых и 50% - серых с нормальными крыльями; б - неполное (частичное) сцепление признаков вследствие перекреста хромосом у самок дрозофилы: РР - самка с длинными крыльями (BBVV) скрещена с черным короткокрылым самцом (bbvv); F 1 - серая самка с длинными крыльями (BbVv) скрещена с черным короткокрылым самцом (bbvv). F 2 - поскольку у самки происходит кроссинговер гомологичных хромосом, образуются четыре типа гамет и появятся четыре вида потомков: некроссоверы - серые с длинными крыльями (BbVv) и черные короткокрылые (bbvv), кроссоверы - черные с длинными крыльями (bbVv), серые короткокрылые (Bbvv).
. Каждый ген занимает определенное место - локус.
. Каждая хромосома представляет собой группу сцепления. Число групп сцепления равно гаплоидному числу хромосом.
Между гомологичными хромосомами происходит обмен аллельными генами. Расстояние между генами пропорционально проценту кроссинговера между ними.
Вопросы для самоконтроля
1. Где находятся гены?
2. Что такое группа сцепления?
3. Чему равно число групп сцепления?
4. Как сцеплены гены в хромосомах?
5. Как наследуется признак длины крыльев и цвета тела у мухи дрозофилы?
6. Потомство с какими признаками проявится при скрещивании гомозиготной самки с длинными крыльями и серым цветом тела с гомозиготным черным самцом с короткими крыльями?
7. Потомство с какими признаками появится при скрещивании дигетерозиготного самца с гомозиготной рецессивной самкой?
8. Какое сцепление генов имеет место у самца дрозофилы?
9. Какое потомство будет при скрещивании дигетерозиготной самки с гомозиготным рецессивным самцом?
10. Какое сцепление генов имеет место у самки дрозофилы?
11. Каковы основные положения хромосомной теории наследственности?
Ключевые слова темы «Хромосомная теория наследственности»
гены
группа сцепления
длина
клетки
конъюгация
кроссинговер
крылья
линейно локус место муха
наследственность
обмен
окраска
организм пары
перекомбинация
поколение
положение
потомки
расстояние
результат
родители
самец
самка
скрещивание
тело
теория
участок
хромосомы
цвет
часть
человек
число
Хромосомный механизм определения пола
Фенотипические различия между особями разного пола обусловлены генотипом. Гены находятся в хромосомах. Есть правила индивидуальности, постоянства, парности хромосом. Диплоидный набор хромосом называют кариотипом. В женском и мужском кариотипе 23 пары (46) хромосом (рис. 78).
22 пары хромосом одинаковы. Их называют аутосомами. 23-я пара хромосом - половые хромосомы. В женском кариотипе одина-

Рис. 78. Кариотипы разных организмов. 1 - человека; 2 - комара; 3 растения скерды.
ковые половые хромосомы ХХ. В мужском кариотипе половые хромосомы XY. Y-хромосома очень мала и содержит мало генов. Сочетание половых хромосом в зиготе определяет пол будущего организма.
При созревании половых клеток в результате мейоза гаметы получают гаплоидный набор хромосом. В каждой яйцеклетке есть 22 аутосомы+Х-хромосома. Пол, образующий гаметы, одинаковые по половой хромосоме, называют гомогаметным полом. Половина сперматозоидов содержит - 22 аутосомы+Х-хромосома, а половина 22 аутосомы+Y. Пол, образующий гаметы, различные по половой хромосоме, называют гетерогаметным. Пол будущего ребенка определяется в момент оплодотворения. Если яйцеклетка оплодотворена сперматозоидом, имеющим Х-хромосому, развивается женский организм, если Y-хромосому - мужской (рис. 79).

Рис. 79. Хромосомный механизм образования пола.
Вероятность рождения мальчика или девочки равна 1:1 или 50%:50%. Такое определение пола характерно для человека и мле- копитающих. У некоторых насекомых (кузнечики и тараканы) нет Y-хромосомы. Самцы имеют одну Х - хромосому (Х0), а самки - две (ХХ). У пчел самки имеют 2n набор хромосом (32 хромосомы), а самцы - n (16 хромосом). У женщин в соматических клетках две половые Х-хромосомы. Одна из них образует глыбку хроматина, которая бывает заметна в интерфазных ядрах при обработке реактивом. Эта глыбка - тельце Барра. У мужчин тельце Барра отсутствует, потому что у них всего одна Х-хромосома. Если при мейозе в яйцеклетку попадает сразу две ХХхромосомы и такая яйцеклетка будет оплодотворена сперматозоидом, то зигота будет иметь большее число хромосом.

Например, организм с набором хромосом ХХХ (трисомия по Х- хромосоме) по фенотипу - девочка. У нее недоразвиты половые железы. В ядрах соматических клеток выделяются два тельца Барра.
Организм с набором хромосом ХХY (синдром Клайнфельтера) по фенотипу - мальчик. У него недоразвиты семенники, отмечается физическая и умственная отсталость. Есть тельце Барра.
Хромосомы ХО (моносомия по Х-хромосоме) - определяют синдром Шерешевского-Тернера. Организм с таким набором - девочка. У нее недоразвиты половые железы, малый рост. Нет тельца Барра. Организм, не имеющий Х-хромосомы, а содержащий только Y- хромосому - нежизнеспособен.
Наследование признаков, гены которых находятся в Х- или Y- хромосомах, называют наследованием, сцепленным с полом. Если гены находятся в половых хромосомах, они наследуются сцепленно с полом.
У человека в Х-хромосомах есть ген, определяющий признак свертывания крови. Рецессивный ген вызывает развитие гемофилии. В Х-хромосоме есть ген (рецессивный), который отвечает за проявление дальтонизма. У женщин две Х-хромосомы. Рецессивный признак (гемофилия, дальтонизм) проявляется только в том случае, если гены, отвечающие за него, будут находиться в двух Х-хромосомах: X h X h ; X d X d . Если в одной Х-хромосоме будет доминантный ген Н или D, а в другой - рецессивный h или d, то гемофилии или дальтонизма не будет. У мужчин одна Х-хромосома. Если в ней есть ген H или h, то эти гены обязательно проявят свое действие, потому что Y-хромосома не несет этих генов.
Женщина может быть гомозиготна или гетерозиготна по генам, локализованным в Х-хромосоме, но рецессивные гены проявляются только в гомозиготном состоянии.

Если гены находятся в Y-хромосоме (голандрическое наследование), то признаки, ими обусловленные, передаются от отца сыну. Например, через Y-хромосому наследуется волосатость ушей. У мужчин одна Х-хромосома. Все гены, находящиеся в ней, в том числе и рецессивные, проявляются в фенотипе. У гетерогаметного пола (мужского) большинство генов, локализованных в Х-хромосоме, находятся в гемизиготном состоянии, т. е. не имеют аллельной пары.
Y-хромосома содержит некоторые гены, гомологичные генам Х-хромосомы, например, гены геморрагического диатеза, общей цветной слепоты и др. Эти гены наследуются как через Х-, так и через Y-хромосому.
Вопросы для самоконтроля
1. Какие правила хромосом имеются?
2. Что такое кариотип?
3. Сколько аутосом у человека?
4. Какие хромосомы у человека отвечают за развитие пола?
5. Какова вероятность рождения мальчика или девочки?
6. Как определяют пол у кузнечиков и тараканов?
7. Как определяют пол у пчел?
8. Как определяют пол у бабочек и птиц?
9. Что такое тельце Барра?
10. Как можно определить наличие тельца Барра?
11.Чем можно объяснить появление большего или меньшего числа хромосом в кариотипе?
12.Что такое сцепленное с полом наследование?
13. Какие гены у человека наследуются сцепленно с полом?
14. Как и почему проявляют свое действие рецессивные гены, сцепленные с полом у женщин?
15. Как и почему проявляют свое действие рецессивные гены, сцепленные с Х-хромосомой у мужчин?
Ключевые слова темы «Хромосомное определение пола»
аутосомы
бабочки
вероятность
волосатость ушей
гаметы
генотип
гены
гетерогаметный пол
глыбка хроматина
гомогаметный пол
дальтонизм
девочка
действие
женщина
зигота
индивидуальность
кариотип
кузнечики
мальчик
мейоз
млекопитающее
момент
моносомия
мужчина
набор
насекомые
наследование
носитель
обработка реактивом оплодотворение
организм
особь
парность
пары
пол
половые клетки
потомство
правила
признак
птицы
пчелы
развитие
различия
рождение
рост
свертывание крови семенники синдром Дауна
синдром Клайнфельтера
синдром Шершевского-Тернера
слепота
созревание
состояние
сочетание
сперматозоиды
сын
тараканы
тельце Барра
трисомия
Y-хромосома
фенотип
хромосома
Х-хромосома
человек
ядро
яйцеклетка
§ 5. Т. Г. Морган и его хромосомная теория
Томас Гент Морган родился в 1866 г., в штате Кентукки (США). Окончив в двадцать лет университет, в двадцать четыре года Морган удостаивается звания доктора наук, а в двадцать пять лет становится профессором.
С 1890 г. Морган занимается экспериментальной эмбриологией. В первом десятилетии 20-го века увлекается вопросами наследственности.

Звучит парадоксально, но Морган вначале своей деятельности был ярым противником учения Менделя и собирался опровергать его законы на животных объектах - кроликах. Однако попечители Колумбийского университета сочли этот опыт слишком дорогостоящим. Так Морган начал свои исследования на более дешевом объекте - плодовой мушке дрозофиле и затем не только не пришел к отрицанию законов Менделя, но и стал достойным продолжателем его учения.

Исследователь в опытах с дрозофилой создает хромосомную теорию наследственности - крупнейшее открытие, занимающее, по выражению Н. К. Кольцова , "то же место в биологии, как молекулярная теория в химии и теория атомных структур в физике".
В 1909-1911 гг. Морган и его не менее прославленные ученики А. Стёртевант, Г. Меллер, К. Бриджес показали, что третий закон Менделя требует внесения существенных дополнений: наследственные задатки не всегда наследуются независимо; порой они передаются целыми группами - сцепленно друг с другом. Такие группы, расположенные в соответствующей хромосоме, могут перемещаться в другую гомологичную при конъюгации хромосом во время мейоза (профаза I).
Полностью хромосомная теория была сформулирована Т. Г. Морганом в период с 1911 по 1926 г. Своим появлением и дальнейшим развитием эта теория обязана не только Моргану и его школе, но и работам значительного числа ученых, как зарубежных, так и отечественных, среди которых в первую очередь следует назвать Н. К. Кольцова и А. С. Серебровского (1872-1940).
Согласно хромосомной теории, передача наследственной информации связана с хромосомами , в которых линейно , в определенном локусе (от лат. locus - место), лежат гены. Поскольку хромосомы парны, то каждому гену одной хромосомы соответствует парный ген другой хромосомы (гомолога), лежащий в том же локусе. Эти гены могут быть одинаковыми (у гомозигот) или разным (у гетерозигот). Различные формы генов, возникающие путем мутации из исходного, называются аллелями , или аллеломорфами (от греч. алло - разный, морфа - форма). Аллели по-разному влияют на проявление признака. Если ген существует более чем в двух аллельных состояниях, то такие аллели в популяции * образуют серию так называемых множественных аллелей. Каждая особь в популяции может содержать в своем генотипе любые два (но не более) аллеля, а каждая гамета - соответственно лишь один аллель. В то же время в популяции могут находиться индивидуумы с любыми аллелями этой серии. Примером множественных аллелей могут служить аллели гемоглобина (см. главу I, § 5).
* (Популяцией (от лат. popularus - население) называется группа особей одного вида, объединенных взаимным скрещиванием, в той или иной мере изолированная от других групп особей данного вида. )
Степень доминирования в сериях аллелей может возрастать от крайнего рецессивного гена до крайнего доминантного. Можно привести большое число примеров такого типа. Так, у кроликов рецессивным геном серии множественных аллелей является ген с, обусловливающий развитие альбинизма * . Доминантным по отношению к этому гену будет ген c h гималайской (горностаевой) окраски (розовые глаза, белое тело, темные кончики носа, ушей, хвоста и конечностей); над этим геном, а также над геном с доминирует ген светло-серой окраски (шиншилловой) c ch . Еще более доминантной ступенью оказывается ген агути - с а (доминирует над генами с, c h и c ch). Самый доминантный из всей серии ген черной окраски С доминирует над всеми "нижними ступенями аллелей" - генами c, c h , c ch , с а.
* (Отсутствие пигмента (см. главу VII, § 5). )
Доминантность, как и рецессивность аллелей,- не абсолютное, а относительное их свойство. Степени доминантности и рецессивности могут быть различны. Один и тот же признак может наследоваться по доминантному или рецессивному типу.
Так, например, складка над внутренним углом глаза (эпикантус) у монголоидов наследуется доминантно, а у негроидов (бушмены, готтентоты) - рецессивно.
Как правило, заново возникающие аллели рецессивны, наоборот, аллели старых сортов растений или пород животных (еще в большей степени диких видов) - доминантны.
Каждая пара хромосом характеризуется определенным набором генов, составляющих группу сцепления. Именно поэтому группы разных признаков иногда наследуются совместно друг с другом.
Так как соматические клетки дрозофилы содержат четыре пары хромосом (2n = 8), а половые - вдвое меньше (1n = 4), то у плодовой мушки насчитывается четыре группы сцепления; аналогично этому у человека число групп сцепления равно числу хромосом гаплоидного набора (23).
Для ряда организмов (дрозофила, кукуруза) и некоторых хромосом человека * составлены хромосомные, или генетические, карты, представляющие собой схематичное расположение генов в хромосомах.
* (К настоящему времени установить точную локализацию генов человека (если принять во внимание общее число генов) удалось лишь в отдельных и относительно редких случаях, например для признаков, сцепленных с половыми хромосомами. )

В качестве примера приведем хромосомную карту части Х-хромосомы дрозофилы (рис. 24). С большей или меньшей точностью в этой карте отражены последовательность генов и расстояние между ними. Определить расстояние между генами удалось при помощи генетических и цитологических анализов кроссинговера, который происходит при конъюгации гомологичных хромосом во время зигонемы профазы I мейоза (см. главу II, § 7).

Перемещение генов из одной хромосомы в другую происходит с определенной частотой, которая обратно пропорциональна расстоянию между генами: чем меньше расстояние, тем выше процент перекреста (единица расстояния между генами названа в честь Моргана морганидой и равна минимальному расстоянию в хромосоме, которое может быть измерено путем кроссинговера). Кроссинговер изображен на рис. 25.
В настоящее время известно тесное сцепление некоторых локусов генов и для них вычислен процент пере-креста. Сцепленные гены определяют, например, проявление резус-фактора и генов MN-системы крови (о наследовании свойств крови см. главу VII, § 3). В отдельных семьях удалось проследить сцепление резус-фактора с овалоцитозом (наличие примерно 80-90% эритроцитов овальной формы - аномалия протекает, как правило, без клинических проявлений), которые дают около 3% перекреста. До 9% кроссинговера наблюдается между генами, контролирующими проявления групп АВО крови и фактором Lu. Известно, что ген, влияющий на аномалию строения ногтей и колена, также сцеплен с локусами АВО-системы; процент перекреста между ними около 10. Значительно лучше изучены группы сцепления (а следовательно, и хромосомные карты) Х- и Y-хромосом человека (см. главу VII, § 6). Известно, например, что тесно связаны между собой гены, определяющие развитие дальтонизма (цветовой слепоты) и гемофилии (кровоточивости); процент перекреста между ними равен 10.
Правильность гипотезы Моргана была подтверждена в начале века Куртом Штерном (цитологические исследования) и сотрудниками Моргана Теофилусом Пайнтером (цитологом) и Кальвином Бриджесом (генетиком) на гигантских хромосомах слюнных желез личинок дрозофилы (подобных гигантским хромосомам других двукрылых). На рис. 26 показана часть гигантской хромосомы слюнной железы личинки хирономуса (мотыля).

При изучении гигантских хромосом с помощью обычного светового микроскопа хорошо заметна поперечная исчерченность, образованная чередованием светлых и более темных полос дисков - хромомеров ; они образованы сильно спирализованными, плотно лежащими рядом друг с другом участками.
Формирование таких гигантских хромосом называется политенией , т. е. редупликацией хромосом без увеличения их числа. При этом редуплицированные хроматиды остаются рядом, плотно прилегая друг к другу.
Если хромосома, состоящая из пары хроматид, будет девять раз последовательно удваиваться, то число нитей (хромонем) в такой политенной хромосоме будет 1024. Благодаря частичной деспирализации хромонем длина такой хромосомы увеличивается по сравнению с обычной в 150-200 раз.
В 1925 г. Стертевант показал наличие неравного кроссинговера: в одной из гомологичных хромосом может оказаться два одинаковых локуса, в которых располагаются, например, гены, влияющие на форму глаза дрозофилы - Ваr, а в другой - ни одного локуса. Так получились мухи с резко выраженным признаком узких полосковидных глаз (ген Ultra Bar) (см. рис. 31).
Кроме цитологических доказательств правильности хромосомной теории, были проделаны генетические эксперименты - скрещивание разных рас дрозофилы. Так, среди множества сцепленных генов в плодовой мушке имеются два рецессивных гена: ген черной окраски тела (bleack ) и ген зачаточных крыльев (vestigial ).
Назовем их условно генами а и б. Им соответствуют два доминантных аллеля: ген серого тела и нормально развитых крыльев (А и Б). При скрещивании чистолинейных мух аабб и ААББ все первое поколение гибридов будет иметь генотип АаБб. Рассуждая теоретически, во втором поколении (F 2) следует ожидать следующих результатов.

Однако в небольшом, но постоянном проценте случаев встречались необычные потомки из необычных гамет. Таких гамет в каждом скрещивании наблюдалось около 18% (9% Аб и 9% аБ).

Появление таких исключений хорошо объясняется процессом кроссинговера. Таким образом, и генетические исследования позволили установить, что нарушение сцепления - кроссинговер, приводящий к увеличению изменчивости форм, статистически постоянен.
В заключение отметим, что целый ряд положений классической генетики на сегодняшний день претерпел ряд изменений.
Мы многократно употребляли термины "доминантные" и "рецессивные" гены (аллели) и признаки. Однако исследования последних лет показали, что так называемые рецессивные гены могут фактически полностью ими не быть. Правильнее сказать, что рецессивные гены дают очень слабое видимое или невидимое проявление в фенотипе. Но и в последнем случае рецессивные аллели, внешне незаметны в фенотипе, могут быть обнаружены при помощи специальных биохимических методик. Кроме этого, один и тот же ген при одних условиях среды может вести себя как доминантный, при других - как рецессивный.
Так как развитие всех организмов происходит в зависимости и под воздействием внешней среды, то и на проявление генотипа в определенном фенотипе влияют факторы среды (температура, пища, влажность и газовый состав атмосферы, ее давление, наличие патогенных для данного организма форм, химический состав воды, почвы и пр., а для человека и явления социального порядка). В фенотипе никогда не проявляются все генотипические возможности. Поэтому в разных условиях фенотипические проявления близких генотипов могут сильно отличаться друг от друга. Таким образом, в проявлении признака участвуют (в большей или меньшей степени) как генотип, так и среда.